
Abstract
Concerns over fetal bovine serum (FBS) limit the clinical application of cultured tissue-engineered constructs. Therefore, we investigated if platelet-rich plasma (PRP) can fully replace FBS for meniscus tissue engineering purposes. Human PRP and platelet-poor plasma (PPP) were isolated from three healthy adult donors. Human meniscal fibrochondrocytes (MFCs) were isolated from resected tissue after a partial meniscectomy on a young patient. Passage-4 MFCs were cultured in monolayer for 24 h, and 3 and 7 days. Six different culture media were used containing different amounts of either PRP or PPP and compared to a medium containing 10% FBS. dsDNA was quantified, and gene expression levels of collagen types I and II and aggrecan were measured at different time points with quantitative polymerase chain reaction in the cultured MFCs. After 7 days, the dsDNA quantity was significantly higher in MFCs cultured in 10% and 20% PRP compared to the other PRP and PPP conditions, but equal to 10% FBS. Collagen type I expression was lower in MFCs cultured with medium containing 5% PRP, 10% and 20% PPP compared to FBS. When medium with 10% PRP or 20% PRP was used, expressions were not significantly different from medium containing 10% FBS. Collagen type II expression was absent in all medium conditions. Aggrecan expression did not show differences between the different media used. However, after 7 days a higher aggrecan expression was measured in most culture conditions, except for 5% PRP, which was similar compared to FBS. Statistical significance was found between donors at various time points in DNA quantification and gene expression, but the same donors were not statistically different in all conditions. At 7 days cell cultured with 10% PRP and 20% PRP showed a higher density, with large areas of clusters, compared to other conditions. In an MFC culture medium, FBS can be replaced by 10% PRP or 20% PRP without altering proliferation and gene expression of human MFCs.
Introduction
Meniscus tissue engineering will lead to improved biological and functional mechanical properties of the preseeded scaffold. 11 Meniscal fibrochondrocytes (MFCs) have shown to be suitable cells for meniscus tissue engineering. 12 MFCs can be easily isolated from resected meniscus tissue of the patient itself, thereby avoiding immunological responses and providing the proper phenotype. 4 Further, MFCs have been shown to enable in vitro meniscus tissue engineering.12,13
In research for tissue engineering applications, fetal bovine serum (FBS) is widely used as a supplement to the standard cell culture medium to induce proliferation and matrix synthesis of a variety of cells. However, the potential risk of bovine pathogens in FBS serum limits its use in clinical applications.14,15 An alternative for FBS is therefore urgently needed. Platelet-rich plasma (PRP) could potentially be a good alternative for FBS. Platelets contain a variety of growth factors, of which transforming growth factor beta, insulin-like growth factor, vascular endothelial growth factor, and platelet-derived growth factor are the most abundant.16–20 PRP has been shown to stimulate the repair of a wide variety of soft and hard musculoskeletal tissues such as bone, muscle, skin, tendon, and cartilage.21,22 In a number of tissue engineering studies, the effect of various PRP volume percentages supplemented to the medium on MFCs, chondrocytes, and subchondral mesenchymal progenitor cells was investigated.23–26 Ishida et al. studied the effect of PRP (3%, 10%, and 30%) on rabbit MFCs in culture and showed increased proliferation and matrix synthesis compared to their FBS control. 23 Currently, there are three concentrations of FBS used for meniscus cell cultures—1%, 10%, and 20%. 19 However, 10% FBS has been most widely used in the meniscus fibrochondrocyte cultures.19,20,24,27–29
The objective of this study was to determine if and which volume concentrations (5%, 10%, and 20%) of PRP can be used as a functional substitute for the standard 10% FBS in the meniscus fibrochondrocyte cell culture medium.
Materials and Methods
Cell isolation
Meniscal tissue was obtained during a partial meniscectomy on a young physically very active male donor (age 18). MFCs were isolated from the meniscus tissue according to previously described protocols. 13 Isolated MFCs were washed twice with PBS and expanded in a monolayer culture. Once cells reached 70% confluence, the cells were passed once, and the subsequent confluent cell cultures were trypsinized and cryopreserved until use. All cells were cultured in a 5% CO2, 37OC controlled incubator with a standard culture medium. A Standard culture medium was defined as Dulbecco's Modified Eagle Medium (DMEM)/F12+10% FBS+1% penicillin/streptomycin/fungizone (PSF; Gibco).
PRP and platelet-poor plasma
Blood samples were collected from healthy human donors (n=3) after informed consent was obtained. Peripheral venous blood was collected in 4.5 mL hirudin-containing blood collection vials (S-Monovette; Sarstedt AG & Co.) to avoid any influence of platelet-dependent protease activating receptor (PAR) signaling by thrombin.30,31 One half of whole blood was centrifuged at 200g for 15 min to obtain PRP, the other half at 2750 g for 15 min to obtain platelet-poor plasma (PPP). After centrifugation, the upper phase containing PRP and PPP, respectively, was transferred and pooled for each donor. Whole blood, PRP, and PPP were measured with fluorescent flow cytometry to quantify the platelet concentration (XE-5000; Sysmex). PRP and PPP were stored at −20°C until use. The PRP samples were not normalized for platelet count.
Cell culture experiment
MFCs were defrosted and expanded in monolayer till passage 3 in a standard culture medium. Afterward, the cells were trypsinized, and the medium was added to neutralize trypsin, counted (with trypan blue exclusion and DMEM/F12), seeded (50,000 MFCs per well) in 24-well plates, and cultured overnight in a standard culture medium containing FBS. After overnight culture, cells were washed twice with DMEM/F12, and subsequently the experimental culture medium was added to each well. Seven different experimental culture media were tested. The base medium was DMEM/F12+1% PSF with one of the following conditions: 10% FBS, 5%, 10%, or 20% PRP or 5%, 10%, or 20% PPP (v/v). PRP and PPP vials of each donor were defrosted only once before use. MFCs were cultured for 24 h and 3 and 7 days in triplicate for each donor. On days 3 and 6, the experimental medium was refreshed with the corresponding experimental culture medium.
Picogreen analysis
After 24 h and 3 and 7 days of culture, cells from each experimental culture medium (n=3) were frozen in MilliQ (−20°C). After repeated freeze/thaw cycles, the PicoGreen dsDNA assay kit was used according to the manufacturer's protocol to quantify the amount of dsDNA (Invitrogen).
RNA isolation and gene expression
After 24 h and 3 and 7 days, Trizol (Sigma-Aldrich) was added to the cells of each condition (n=3) and stored at −80°C. RNA was isolated via standard chloroform-phase separation. 32 The supernatant was isolated, and RNA was precipitated with isopropanol, washed twice with 75% ETOH, air-dried, solubilised in RNAse-free water, and subjected to reverse transcriptase–polymerase chain reaction (PCR) to obtain cDNA. Finally, a quantitative PCR (qPCR) was performed on individual samples by an ABI/PRISM 7000 sequence detection system (Applied Biosystems). qPCR was prepared as follows: 1 μL forward primer (5 μM), 1 μL reverse primer (5 μM), and 5 μL of Sybr Green PCR Master mix (Applied Biosystems) were added to 3 μL cDNA. qPCR was performed for 40 cycles. Three specific gene expression markers for meniscus tissue engineering, that is, collagen type I and II and aggrecan, were measured with GAPDH as household gene (see Table 1 for primer sequences). The Ct value shows the point in which the amplification plot passes the threshold, due to the release of the fluorescence from mRNA transcription. GAPDH was the internal control for all genes and used to normalize delta Ct levels for any variability in cell numbers. Delta-delta Ct levels were calculated by subtracting the delta Ct level of the gene of interest from the delta Ct level of the control (FBS). The fold change in the gene expression relative to the control was calculated using the equation 2−ΔΔCt. FBS is designated with a dashed line at 1 on the y-axis, as seen in Figures 2 and 3. All primers were validated in the Rheumatology Department at the Nijmegen Centre for Molecular Life Sciences. 33
Primers were obtained from Biolegio bv, Nijmegen, the Netherlands.
Histology
Monolayer cell cultures were fixated after 24 h, 3, and 7 days in their wells, stained with hematoxylin and eosin and mounted using Permacol UV (Permacol) (n=3). All samples were analyzed with light microscopy and photographed with an Olympus digital camera at the same magnification.
Statistics
All data were expressed as mean±SEM. Picogreen data were log-transformed and subjected to a two-way ANOVA with a post hoc Fisher-LSD test. For qPCR, delta Ct levels were subjected to two-way ANOVA with a post hoc Fisher-LSD test. p-Values<0.05 were considered significant. Statistical analysis was performed using SPSS v16.0 software (SPSS, Inc.). Statistical analysis of the Picogreen and qPCR data was completed in two steps. In the first two-way ANOVA, the effect of condition and the time points on dsDNA or gene expression was analyzed with the donors pooled. The second two-way ANOVA analyzed the effect of the donor and condition on time points.
Results
Platelet count
Platelet concentrations were measured in whole blood, PRP, and PPP from each donor. The platelet concentration in whole blood was on average 131±26×109/L (donor 1: 148×109/L, donor 2: 105×109/L, and donor 3: 141×109/L). For PRP, the average platelet count was 140±20×109/L (donor 1: 165×109/L, donor 2: 98×109/L, donor 3: 162×109/L). In PPP, the platelet concentrations were not detectable in all three donors.
DNA quantification
The average amount of dsDNA in cells cultured for 24 h and 3 days in a standard medium containing 10% FBS was 51.5±1.7 and 90.1±7.9 ng/mL, respectively. During these time points, no differences were measured between conditions or in time as a condition (p>0.050, data not shown). However, at 7 days of culture, a significant increase of dsDNA was measured compared to 24-h and 3-day cultures. In addition, significant differences between the culture conditions were measured. Cells grown in a medium containing FBS and 10% and 20% PRP had a significant higher amount of dsDNA compared to all other conditions at 7 days (Fig. 1). Statistical significance between the donors was found only at 7 days; dsDNA in cells cultured with plasma of D2<D3=D1 (respectively, 55.2, 76.4, and 64.9 ng/mL). The average minimum and maximum values of donor-pooled dsDNA are presented in Table 2.
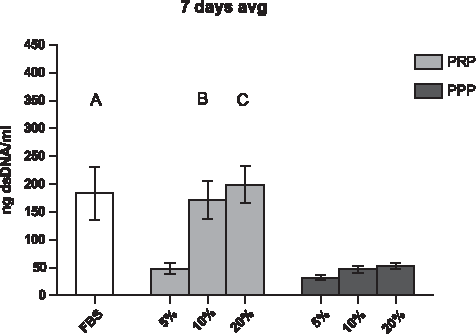
Average dsDNA levels of all donors at 7 days of monolayer cell culture (n=3). A significant increase in dsDNA was measured by using a medium containing 10% and 20% platelet-rich plasma (PRP) (p<0.05). When 5% of PRP was used, dsDNA was significantly later expressed compared to the control (fetal bovine serum [FBS]). A, B, and C represent significance between labeled conditions and all conditions of platelet-poor plasma (PPP) (5%, 10%, and 20%). FBS is designated with a dashed line at 1 on the y-axis, and all the conditions were adjusted to FBS.
PRP, platelet-rich plasma; PPP, platelet-poor plasma.
Gene expression
mRNA of collagen type I in cells cultured for 24 h was upregulated compared to 3 and 7 days (time: p<0.050, Fig. 2). Further, there was a significant difference between the different medium and the control medium containing 10% FBS at 3 and 7 days (Fig. 2B, C). At 3 days, collagen type I mRNA in cells cultured with a medium containing 5% PRP was significantly upregulated if compared to the medium containing 10% and 20% PRP (Fig. 2B). At 7 days of culture, no differences in collagen type I expression were found between the different culture conditions. All PRP and PPP conditions were downregulated at 7 days in collagen type I expression (Fig. 2C). No donor-specific differences were measured at 24 h and 7 days in collagen type I expression. However, at 3 days, D2<D3 (p<0.050) and D1=D2 and D3 (p=0.062 and 0.097, respectively).
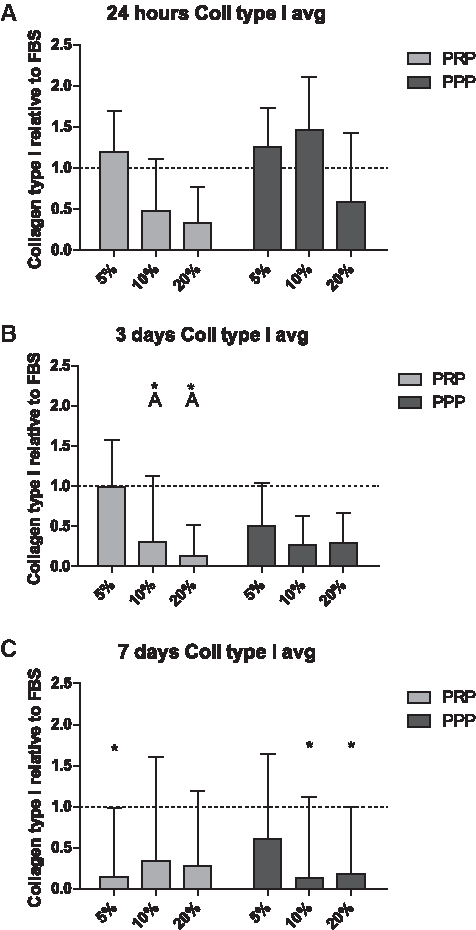
Meniscal fibrochondrocyte (MFC)-related gene expression of collagen type I at 24 h and 3 and 7 days expressed in fold change
Expression of collagen type II mRNA was late or not detected at all time points for all treatments (Ct>40, data not shown). Aggrecan showed to have stable expression for all cells independent of the plasma supplement and culture time (Fig. 3). Although at 7 days, all treatments, except 5% PRP, were upregulated by a minimum of 6-fold increase (Fig. 3C, p=0.096). Donor variability for Aggrecan expression was significantly different after 24 h (D1<D2=D3, p<0.050) and 3 days (D3>D1=D2, p<0.050), but absent at 7 days of culture (p=0.166).

MCF-related gene expression of aggrecan at 24 h and 3 and 7 days expressed in fold change
Histological analysis
At 24 h, MFCs were distributed homogeneously throughout the well plate (Fig. 4). Locally, small clusters of cells were present in each condition. At 24 h of culture, the cells reached an approximate 20% confluence, and their phenotype was mainly fibroblast-like or spindle–shaped, if located individually, and more rounded/chondrocyte-like, if cells were clustered. The phenotype changed at 3 days of culture. More cells were clustered. Cells in the center of the clusters were rounded, toward the edges of the clusters, and isolated cells showed to be mainly spindle-shaped/fibroblast-like. Cells reached an approximate confluence of 40%. At 7 days, the phenotype remained spindle-shaped/fibroblast-like; however, differences in the confluence were observed. Cells cultured with 10% PRP and 20% PRP showed a higher density (∼70% confluence) with large areas of clusters that were fused together, compared to other conditions (40%–50% confluence). No differences between donors were observed (results not shown).
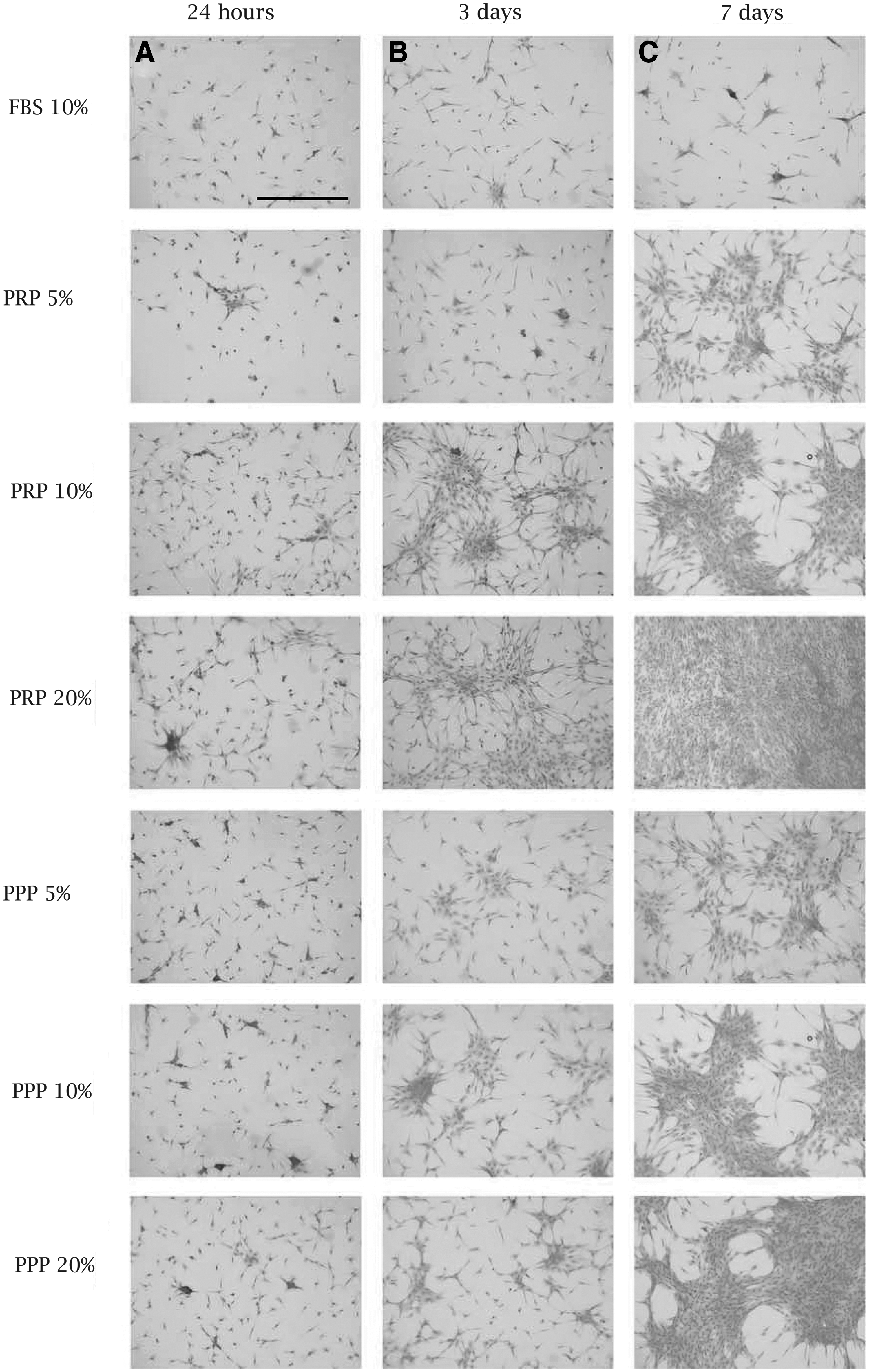
Hematoxylin-&-eosin histological analysis of MFCs in 24 h, 3 days, and 7 days in all conditions.
Discussion
Tissue engineering appears to be a promising treatment option for (partial) meniscus replacement. 34 In most meniscal tissue engineering studies, FBS is used as a potent supplement in the culture medium.23,34 However, clinical application of FBS in a cultured construct is not allowed because of the risk of disease transfer or xenoimmunization.14,15 In the present study, we showed that concentrations of 10% and 20% of PRP as an alternative for FBS supplementation induce similar effects for proliferation and gene expression on cultured MFCs as compared to the standard medium containing 10% FBS.
PRP can easily be isolated from patients' own blood or donors. Surprisingly in this study, the platelet concentrations did not increase from whole blood to PRP. This could be related to the method used for PRP isolation. Differences in centrifugation speed, the vials used, and centrifugation time have shown to influence the final platelet concentrations in PRP.22,35–38 Further, during centrifugation, platelets might be trapped in the buffy coat located between the RBCs and plasma. Since we observed that in hirudin blood vials, as were used in this study, the buffy coat was thinner compared with the citrate blood vials, we speculate that this might be one of the reasons for the relative low platelet counts. Hirudin also inhibits PAR signaling by thrombin, which also might be a factor to cause a thinner buffy coat.30,31
We showed that proliferation of MFCs remained similar until 3 days, but at 7 days, differences in the cell numbers were found between conditions. Supplements of 10% and 20% PRP strongly showed an increase in the number of cells compared to PPP samples. Therefore, the effects can be related to the platelet content and not solely to the plasma. Furthermore, this effect seems platelet concentration-dependent, since the effect was not observed at 5% PRP. Ishida et al. studied also PRP concentration-dependent effects on meniscus fibroblasts. They investigated short-term effects (48 h) of PRP and PPP on the vitality of cultured meniscus cells. 23 After a preculture of 24 h in 10% FBS, the cultures were continued for 24 h in 1% FBS supplemented with 3%, 10%, 30% PRP, or similar concentrations of PPP. 23 PRP concentrations upregulated the viability of meniscus cells in a dose-dependent manner after 48 h. A clear comparison with our data is difficult, since the study of Ishida et al. was shorter (48 h), and the control was 1% FBS, and the PRP sand PPP was added to this 1% FBS. 23 In our study, we compared the effect of PRP and PPP to 10% FBS and studied the increase in cell numbers based on DNA and not vitality of the cells. In an in vivo model, they described a positive effect of PRP on defect healing in the meniscus compared or PPP, but no details were included about the concentration of PRP or platelets that accounted for this positive effect. 23
Collagen type I was initially upregulated in 5% PRP as well as 5% and 10% PPP at 24 h. However, in time, collagen type I was downregulated in both PRP and PPP compared to FBS at 3 and 7 days. Collagen type II was not expressed in our experiment, which might be due to that MFCs were used at passage 4 according to Gunja et al. 39 Collagen type II expression decreases as the passage number increases. 39 Aggrecan at 24 h and 2 days had similar or lower expression than that of FBS in all conditions of PRP and PPP. However, at 7 days, aggrecan was upregulated in 10% and 20% PRP and all PPP concentrations compared with FBS. The 5% PRP concentration was comparable to the 10% FBS control. This shows that potentially in the PPP and RRP serum, interesting factors are present that can influence the expression of fibrochondrocyte-specific genes independent from the effect of the platelets. Since it is well known that the meniscus fibroblasts change their phenotype in cultures at increasing passages,28,39 further research in specific factors in PPP and PRP might be beneficial for future meniscus tissue engineering. Our result also differs from the reported decreased aggrecan expression in PRP conditions reported by Ishida et al. 23 A possible explanation for this discrepancy in the amount of aggrecan expression might be that the metabolic half-life of aggrecan is much shorter compared to collagen. 40 We speculate that the age of the MFCs might play a crucial role, since aggrecan expression increases after puberty. 41 McAlinden et al. stated that aggrecan in the meniscus is important to withstand the compressive forces. 41 The increased amount of aggrecan expression in our results may be a combined effect of the age of the subject and the fact that the donor was physically very active. Karube and Shoji already showed that the GAG content of menisci decreases with aging. 42 The age of the rabbits of the study of Ishida et al. was not mentioned in their article, and further detailed studies are needed to clarify this.
There are not many studies that describe a clear effect of PRP and PPP on cell proliferation. Mazzocca et al. studied the effect of PRPs obtained by different separation methods on osteoblasts, monocytes, and on tenocytes. 22 They compared the effects of three different PRP isolation procedures on proliferation (assessed by thymidine incorporation). They found a similar positive effect of all PRPs compared to 2%, 5%, 10%, or 15% bovine serum albumin, which was used as a control (dependent on the cell type). The different PRP had all a stimulatory effect on the proliferation of the investigated cell types compared to the FBS controls, but since they mixed PRPs with 2% FBS in the final tests, their results did not allow the conclusion that PRPs can replace FBS completely.
Besides an effect on proliferation, PRP might stimulate the migration and differentiation of cells. Kruger et al. showed that PRP can stimulate the migration of subchondral progenitor cells and stimulate chondrogenic differentiation of the stem cells. 26 This might also explain the positive effect on gap healing as described by Ishida et al. by PRP added to an acellular gelatin hydrogel. 23
Collagen type II expression remained undetermined at all time points. This lack of expression can be due to improper mRNA isolation. However, based on the published results in which collagen type II expression was clearly detectable in chondrocytes with the same set of primers, it is more likely due to a low or a lack of expression of collagen type II in these cells. 33 It has been reported that differences in the stimulation or culture conditions can influence the expression of the collagen-type matrix proteins. 28 In an engineered meniscus construct, the final extracellular matrix should predominantly consist of collagen type I similar to that of the native meniscus tissue, although other collagens such as collagen type II, III, and VI have been shown to be present in the meniscus as well. 43
Some differences in responsiveness between the donors were measured at the earlier time points for collagen type I and aggrecan expression, while the results between donors leveled out at 7 days of culture. Despite the fact that the platelet number may be responsible for these observed variations, a large study reported a poor correlation between platelet counts and specific growth factor concentrations.18,44 However, other studies demonstrated a clear effect between the growth factor content of platelets and the biological activity that was induced. 45 Further, it is well known that besides the growth factors, platelets contain a variety of other proteins or chemokines, which also might influence the reaction of cells or tissues. 46
In conclusion, the medium supplemented with 10% and 20% PRP can replace a standard medium supplemented with 10% FBS in MFC culture while maintaining proliferation and gene expression levels. This opens new perspectives with respect to the clinical utility of MFC in tissue engineering and the use of PRP as an autologous supplement.
Footnotes
Acknowledgments
The Dutch Arthritis Foundation and the Netherlands Institute for Regenerative Medicine sponsored this study. The authors would like to acknowledge Elly Vitters for her contribution in qPCR analysis (Department of Experimental Rheumatology, NCMLS Nijmegen, The Netherlands).
Disclosure Statement
No competing financial interests exist.