
Abstract
Low-back pain caused by intervertebral disc degeneration could be recovered by the regeneration of the nucleus pulposus (NP). This study aimed to establish a chondrogenic recovery model with promising a human NP (hNP) cell line, an immortalized hNP (ihNP), which could be a screening platform to identify regenerative drugs. The ihNP cells were created from primary human NP cells transfected with a retroviral vector-driven HPV16 E6/E7. Growth properties and characteristics of ihNP were evaluated by comparing with parental NP cells. Successful immortalization of ihNP cells stably expressed HPV 16 E6/E7 mRNA. The doubling time of ihNP was shortened to 53.16±2.63 h compared with parental hNP-P1. Cell cycle regulators, including p53, p21, and pRB were downregulated compared to parental hNP-P1. The in vivo neoplastic forming assay also demonstrated that the ihNP was nontumorigenic. After 25 generations of cell cultures, the ihNP cells, yet stably expressed chondrogenic genes, including (SOX9), type II collagen (Col II), aggrecan, decorin, biglycan, and versican. Higher expressions of chondrogenic proteins, including Col II, phosphorylated SOX9 (p-SOX9), and CD44 were also determined. Under the stressful inflammatory conditions induced by lipopolysaccharides (LPS), the regenerative and anti-inflammatory potentials of ihNP in two-dimensional culture with the presence of platelet-rich plasma (PRP) were evaluated by reverse transcriptase polymerase chain reaction. PRP showed significant effects on restoring diminished chondrogenic markers and deleterious inflammatory responses induced by LPS in ihNP. The therapeutic potentials of ihNP in three-dimensional neocartilage model could also be exerted by PRP using histological evaluation and immunological staining. Hence, the established ihNP cells can provide a chondrogenic recovery model as a regenerative drug screening tool for further regenerative drug discovery and development.
Introduction
L
Models for regeneration basically consist of three components, including scaffold, survival signals, and cells. First, a collagen gel mimicking a cartilaginous microenvironment provides an engineered scaffold for regeneration of IVD and articular cartilage.11–15 The chondrocytes/collagen-generated neocartilage preserve chondrogenic capabilities for remodeling and growth, which are bearing the potentials to be used for repairing or replacing degenerative tissue. Second, platelet-rich plasma (PRP), as a survival signal, stimulates anti-inflammatory and chondrogenic potentials of chondrocytes and has been demonstrated in our previous studies both in vitro and in vivo.14–16 Moreover, cell is the most important factor in a regeneration model. Originally, for disc regeneration, primary chondrocytes such as NP cells were used in the model for regeneration.14,17 However, primary chondrocytes isolated from cartilaginous tissue lost its phenotype, including the decreased cell proliferation and matrix synthesis during expanded culture. The process of primary culture is also inconvenient to gain sufficient cell materials for model insight into regenerative medicine. 18 Thus, the alternative chondrocyte cell line with genetic modification through viral vector-mediated gene transfer is emergent.18–22
For genetic modification, telomerase reverse transcriptase (TERT) and oncogene SV-40LT have been introduced into primary cells. However, the immortalization resulted in disadvantages, including cell hypertrophy, senescence, and karyotypic instability.20,23,24 Immortalizing cells with human papillomavirus-16 E6/E7 (HPV-16 E6/E7) efficaciously extend the stage of growth before senescence well beyond normal. 25 Previously, we have successfully established hPi cells, an immortalized human articular chondrocytes by HPV-16 E6/E7, preserving the relevant features without neoplastic transformation. 13 Based on this hPi cell line, the in vitro stage-specific chondrogenesis of mesenchymal stem cells (MSCs) was first elucidated. 12 The hPi cells were also served as a screening platform for the anabolic response and metabolic stimulus. 15
For disc therapy, there is still no efficient screening platform for disc regenerative drug development. The aim of this study was to establish a NP cell line, immortalized human NP (ihNP), through the immortalization of HPV-16 E6/E7. The ihNP cells not only provide a promising cartilage regeneration model, but also produce an extracellular matrix (ECM) and maintain the innate phenotype almost identical to their parental cells. Their characteristics were determined by the expressions of chondrogenic genes and proteins. Moreover, the therapeutic potentials were also evaluated under the degenerative environment. The established functional cell model ihNP can be viewed as an effective screening platform of regenerative drug development for IVD regeneration.
Materials and Methods
Isolation and culture of human NP
hNP cells were collected from a 39-year-old male IVD donor. The volunteer gave an informed consent for the use of the NP cells in accordance with the principles of the Ethics Committee of Taipei Medical University Hospital. The NP tissue was harvested aseptically and minced into pieces in the Hank's balanced salt solution (HBSS; Gibco BRL) containing antibiotics. NP cells were then isolated from these slices in an enzymatic solution (0.4% collagenase I and 0.04% protease, purchased from Sigma) for 4 h at 37°C. The cell suspension was filtered through a 40-μm nylon mesh, centrifuged, and then resuspended in the Dulbecco's modified Eagle's medium (DMEM/F-12; Gibco BRL) with 10% fetal bovine serum (FBS). Isolated NP cells were then seeded into 10-cm tissue culture dishes and incubated at 37°C in 5% CO2 before subsequent experiments.
Transfection with HPV-16 E6/E7 gene
The retroviral vector transduction has been previously described. 26 In brief, the HPV-16 E6/E7 retroviral vector (LXSN16E6E7)26,27 containing a virus produced by the PA317 cell line (purchased from the American Type Culture Collection, ATCC) was expanded in the DMEM with 10% FBS (Gibco). The isolated primary hNP cells were recovered by trypsinization, seeded on six-well plates at a density of 2.5×105 cells per well, and infected with 1 mL of virus stock in a medium containing 8 μg/mL polybrene for 48 h. The virus was then removed and the medium was replaced with the DMEM/F12 supplemented with 10% FBS. The cells were passaged at 1:3 on the next day. Three weeks after transduction, cells were harvested when obvious clones of actively dividing cells developed. The infected cells, ihNP cells, were continuously grown in the same medium as used for primary culture twice a week and passaged at a 1:3 ratio when the cells appeared subconfluently.
Cell viability
Cell viability was measured using MTT assay (Sigma). Cells were seeded into 96-well plates at a density of 4×103 cells/well and cultured for 7 days. At day 1, 3, 5, and 7, 20 μL MTT was added to each well and incubated for 4 h. The medium was then removed from each well by suction and 150 μL DMSO (Sigma) was used to lyse cells. The absorbance of the cell lysates was measured at 570 nm by using a Multiskan PC (Thermo Labsystem). Cell population doubling time was calculated using the following function as previously described
26
:
T–T0 indicates the length of time between two measurements and N0 and N denote the OD value at two points of measurement. The experiments performed in triplicate were evaluated.
Tumorigenicity assay
The animal experiment was conducted in compliance with the protocol approved by the Institutional Animal Care and Use Committee of Taipei Medical University. The immortalized cells were trypsinized from confluent monolayer cultures and resuspended at 4×106 cells/mL in phosphate-buffered saline (PBS) (collected at passage 25). The suspended mixture was injected subcutaneously into the dorsa of each SCID/NOD mice (from National Taiwan University Laboratory Animal Center). Mice were kept in sterilized pathogen-free cages and observed daily for tumor formation over 3 months. HeLa cells were injected as positive controls and nontransfected hNP cells were used as negative controls.
Reverse transcriptase polymerase chain reaction
Total RNA was harvested and extracted using the TRIzol Reagent (Invitrogen Life Technologies). Extracted RNA was dissolved in sterilized ddH2O and stored at −80°C until use. Reverse transcription was performed with SuperScript™ III (Invitrogen Life Technologies) and an Oligo d (T)12–18 primer. Four micrograms of RNA was added into a final solution volume of 21 μL containing 10 mM dNTP mix, 10×RT buffer, 25 mM MgCl2, 0.1 M DTT, RNase Inhibitor, and RNase H. Six micrograms of reverse transcriptase (RT) product was used in polymerase chain reaction (PCR) amplification in a final volume of 50 μL containing 2.5 mM dNTP, 25 mM MgCl2, upstream/downstream primers (see Table 1), and Taq DNA polymerase (Invitrogen Life Technologies). Following an initial denaturation at 95°C for 5 min, the DNA was amplified in the Touchgene Gradient (Techne) using 35 cycles of 1 min at 94°C for denaturation and extension at 72°C for 1 min. This was followed by a final extension at 72°C for 5 min. The annealing temperature was different for specific genes (See Table 1). PCR products were analyzed on 2% agarose gels (Agarose I) and visualized with ethidium bromide (EtBr) staining. Images were analyzed using FloGel-I (Fluorescent Gel Image System). Quantification was normalized using Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an internal control.
Gel electrophoresis and Western blotting
Cells were trypsinized and dissolved in the RIPA buffer (50 mM Tris pH 7.4, 150 mM NaCl, 0.5% DOC, 1% NP-40, 0.1% SDS), followed by centrifugation for 15 min at 12,000 rpm at 4°C. The upper fluid, containing total protein, was extracted. The 40 μg of extracted protein among each groups was mixed with 6×sample buffer, denatured for 5 min at 95°C, and then separated in a 10% SDS-PAGE gel. The protein bands were transferred to a nitrocellulose membrane, blocked overnight in a blocking buffer, and then incubated with primary antibodies: (i) for the chondrogenic-specific protein, including rabbit anti-human pSox9 (Ser181) (1:250; Abcam), rabbit anti-type II collagen (1:500; Abcam), and anti-mouse/human CD44 (clone IM7) monoclonal antibody (1:400; BioLegend); (ii) for cell cycle regulatory proteins, rabbit anti-human phospho p53 (1:1000), rabbit anti-human p21 (1:2000), rabbit anti-human phospho Rb (1:1000), rabbit anti-human cyclin E2 (1:500), and mouse anti-human cyclin A2 (1:500), all purchased from Millipore corporation. The β-actin (1:5000; Millipore) was used as an internal control. After a brief wash with PBS-T, a peroxidase-labeled secondary antibody was added (diluted 1:5000 in PBS-T) and incubated for 1 h. Protein bands were detected by the ECL plus-Kit (Amersham Pharmacia) and visualized with a Biospectrum AC Imaging System (UVP BioImaging Systems).
Alcian blue staining
Cells were washed twice with PBS and fixed in 10% formalin for 10 min at room temperature. After washing with distilled water, cells were stained with 1% Alcian blue 8GX (Sigma) in 3% glacial acetic acid (pH 2.5) for 3 h at room temperature, and then washed twice with distilled water. The cells were observed under a microscope, followed by gradual elution of the bound dye by a dissociated solution. The eluted solutions were examined using a spectrophotometer (Thermo Labsystem) for quantification. The dissociated solution was used as the blank group.
Senescence-associated β-galactosidase staining
Cell senescence was determined by staining for senescence-associated β-galactosidase (SA-β-gal) using a histochemical staining kit (Sigma). Briefly, hNP or ihNP cells were fixed for 6–7 min in a fixation buffer at room temperature. After washing with PBS, the cells were incubated with the β-gal chromogenic substrate solution for 12 h at 37°C without CO2. The reaction was stopped by the addition of PBS and the stained cells were counted. The experiment was repeated at least three times and the mean value of percentage of SA-β-gal expressing cells was calculated.
Proinflammatory cytokines and PRP treatment
To create an in vitro model featuring important inflammatory cellular states, lipopolysaccharides (LPS) were used to induce proinflammatory cytokines in two-dimensional (2D) cells and three-dimensional (3D) cultures. For the regeneration group, PRP was utilized as the growth factor complex that has been demonstrated to regenerate NP tissue. 14 The concentration of 1 ng/mL TGF-β1 (the highest growth factor in PRP as an indicator) of PRP has been demonstrated to be the optimal concentration for disc regeneration and was also used in this study. Briefly, ihNP cells were cultured in the DMEM/F12, and then exposed to LPS (200 ng/mL), LPS (200 ng/mL)+PRP (TGF-β1=1 ng/mL), or none of them (as control). Media were changed every 2 days and cells were cultured for 1 week. After 7 days of treatment, chondrocytic degeneration or regeneration was evaluated by gene expression analysis using RT-PCR. Differences in the expressions of SOX9, type II collagen (Col II) and aggrecan, IL-1β, TNF-α and MMP-3 between hNP and ihNP cells were compared. In this test, PRP with the optimal concentration (TGF-β1=1 ng/mL) determined in our previous study 14 was applied.
Three-dimensional constructs in collagen/PRP scaffolds
Three-dimensional (3D) chondrocytes in collagen scaffolds have been described in our previous studies.13–15 The ihNP cells were expanded, and then embedded at a density of 106 cells/mL in collagen matrices, including 2 mg/mL of purified type I+type II collagen (type I: type II=4:1) mixture. The cell/collagen constructs were then subjected into the rotatory cell culture system (RCCS, Synthecon) for neocartilage formation. The constructs were then cultured in the DMEM/F12 containing 10% FBS as the control group, while those cultured in a medium containing LPS (200 ng/mL) were used as the degeneration group. Whereas constructs cultured in a medium containing the combination of LPS and PRP were defined as the regeneration group to detect the effects of PRP. All the constructs were cultured in a 37°C, 5% CO2 incubator for 4 weeks, and then histologically analyzed with hematoxylin and eosin staining, immunohistochemistry staining (IHC) using specific antibodies for Col II (MAB1330; Chemicon International) and CD44 (103013; BioLegend), and Alcian blue staining for proteoglycan (PG). The mouse anti-human Col II monoclonal antibody (Chemicon International) used in IHC only reacts with human and bovine Col II.
Statistical analysis
The results of all experiments are shown as the mean±standard deviation (SD). Differences of mRNA expression in the response to LPS and PRP were compared and evaluated using the paired t-test. A p value<0.05 was considered as a significant difference.
Results
Cell morphology and HPV-16 E6/E7 detection
For immortalization, the amphotropic retroviral vector LXSN16E6E7 was transduced into the primary hNP chondrocytes, while after one passage from primary culture. The transduced hNP cell was designated as ihNP cells and showed a spindle-shaped fibroblast-like morphology at passage 25 (ihNP25), similar to its parental hNP cells at passage 1 (hNP-P1) (Fig. 1A). The RT-PCR products of the HPV-16 E6/E7 gene in ihNP25 cells showed a distinct band at 628 bp. In contrast, the band was not detected in parental hNP cell (Fig. 1B).
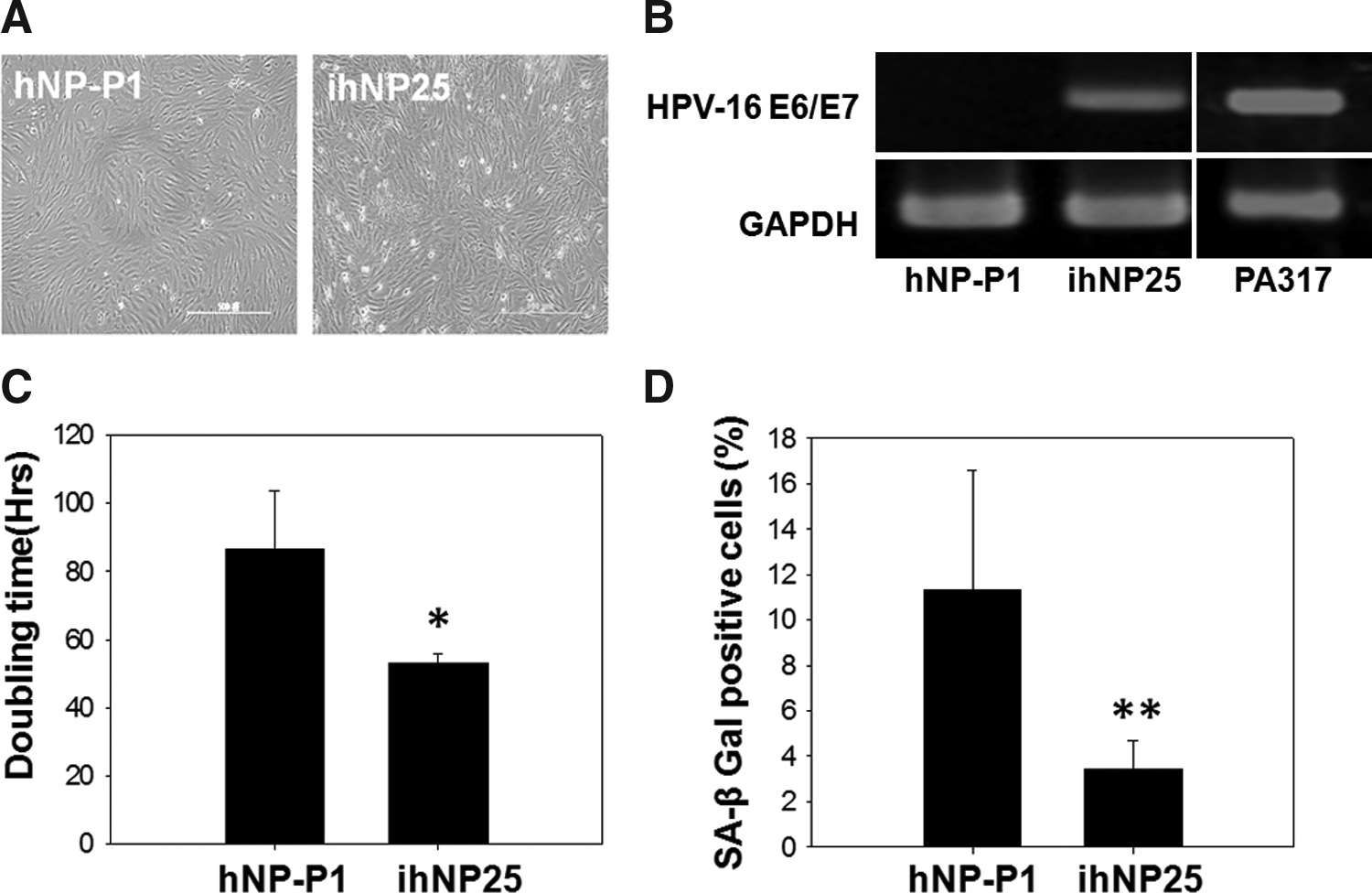
Characterization of immortalized human nucleus pulposus (ihNP) cells.
Decreased cell senescence of ihNP cells
After immortalization, the effects of the HPV-16 E6/E7 gene on cell viability and cell senescence of ihNP cells were examined. The population doubling time of ihNP25 cells was 53.16±2.63 h, which is shorter than those of hNP-P1 cells (86.5±17.34 h) (Fig. 1C). In addition, senescence was assayed by the staining of SA β-gal, which showed a significant decrease in ihNP25 cells (3.43%±1.24%) compared to hNP-P1 cells (11.40%±5.31%) (Fig. 1D).
The regulation of cell cycle in ihNP cells
To elucidate the mechanism of HPV-16 E6/E7 during immortalization, the essential regulators participating in cell cycle were examined in both parental hNP-P1 and ihNP25 cells. After immortalization (ihNP25), the phosphorylation of p53 (Phospho-p53) and pRb (Phospho-Rb) were both entirely downregulated compared to parental hNP-P1 (Fig. 2A). p21 was also subsequently suppressed after phospho-p53 inhibition. In addition, cyclin E2 and A2, regulating cell cycle progression at G1/S transition, were both highly upregulated in ihNP cells.
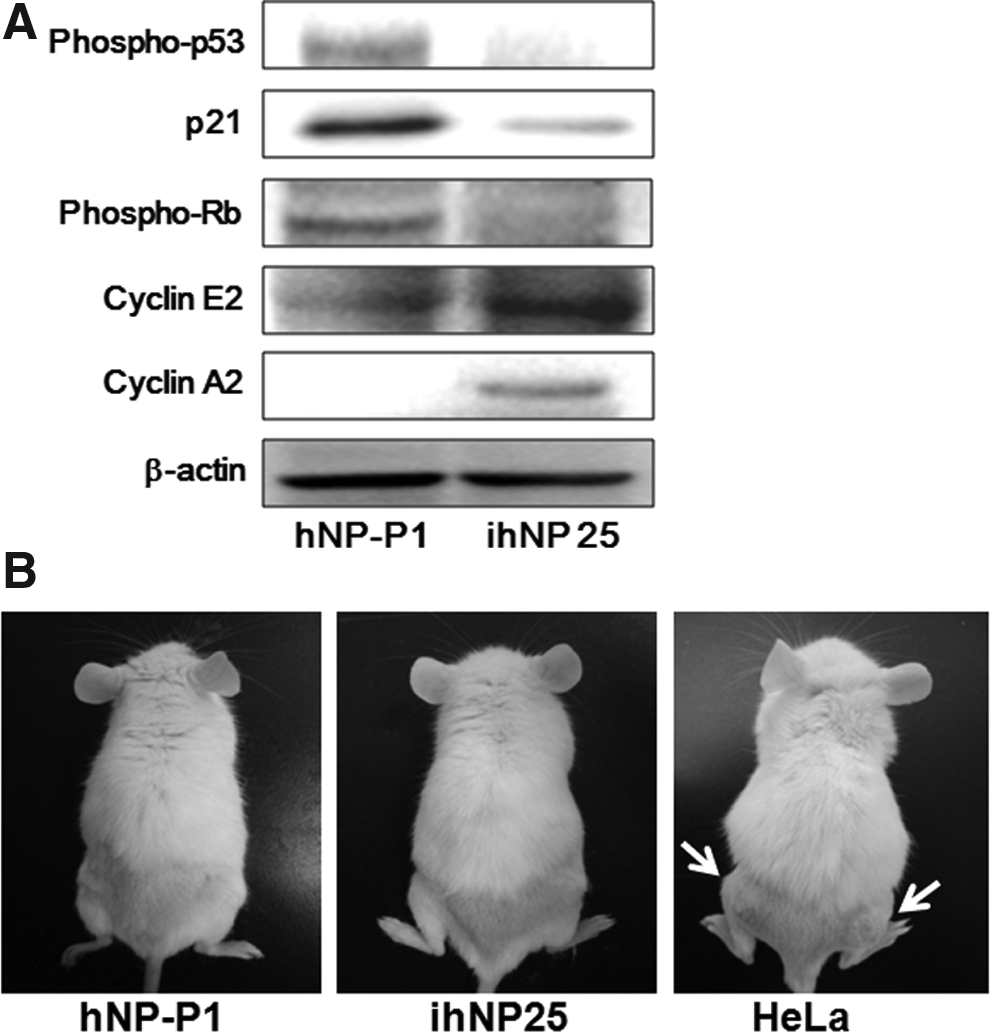
Cell cycle regulation and tumorigenicity of ihNP cells. Comparisons of
Nontumorigenicity of ihNP cells
Due to the inhibition of p53 and Rb protein by HPV-16 E6/E7 transfection, more concern should be put on tumorigenicity of ihNP25. To determine the tumorigenicity of HPV-16 E6/E7-transduced ihNP cells, 4×106 cells/mL of ihNP25 cells were injected subcutaneously into the dorsa of SCID/NOD mice, while the same number of hNP-P1 and HeLa cells (a malignant cancer cell line) were used as negative and positive control, respectively (Fig. 2B). No tumor mass was formed in the mice injected with parental hNP-P1 or immortalized ihNP25 cells for more than 3 months. However, tumor masses were found on the dorsa of mice injected with HeLa cells after 3 days. The results suggested that ihNP cells are immortalized, but nontumorigenic.
Chondrogenic-specific gene expression profile of ihNP cells after immortalization
To determine the maintenance of chondrogenic-specific gene expression profiles between parental and immortalized cells, hNP-P1 and ihNP cells were analyzed by using RT-PCR. Both ihNP25 and hNP-P1 highly expressed NP-specific genes, including SOX9, Col II, aggrecan, decorin, biglycan, and versican (Fig. 3). No significant difference between parental hNP-P1 and ihNP25 cells was found after quantification (data not shown). In contrast, these specific genes were rarely detectable in parental hNP cells after culture for 5 passages in vitro (data not shown).
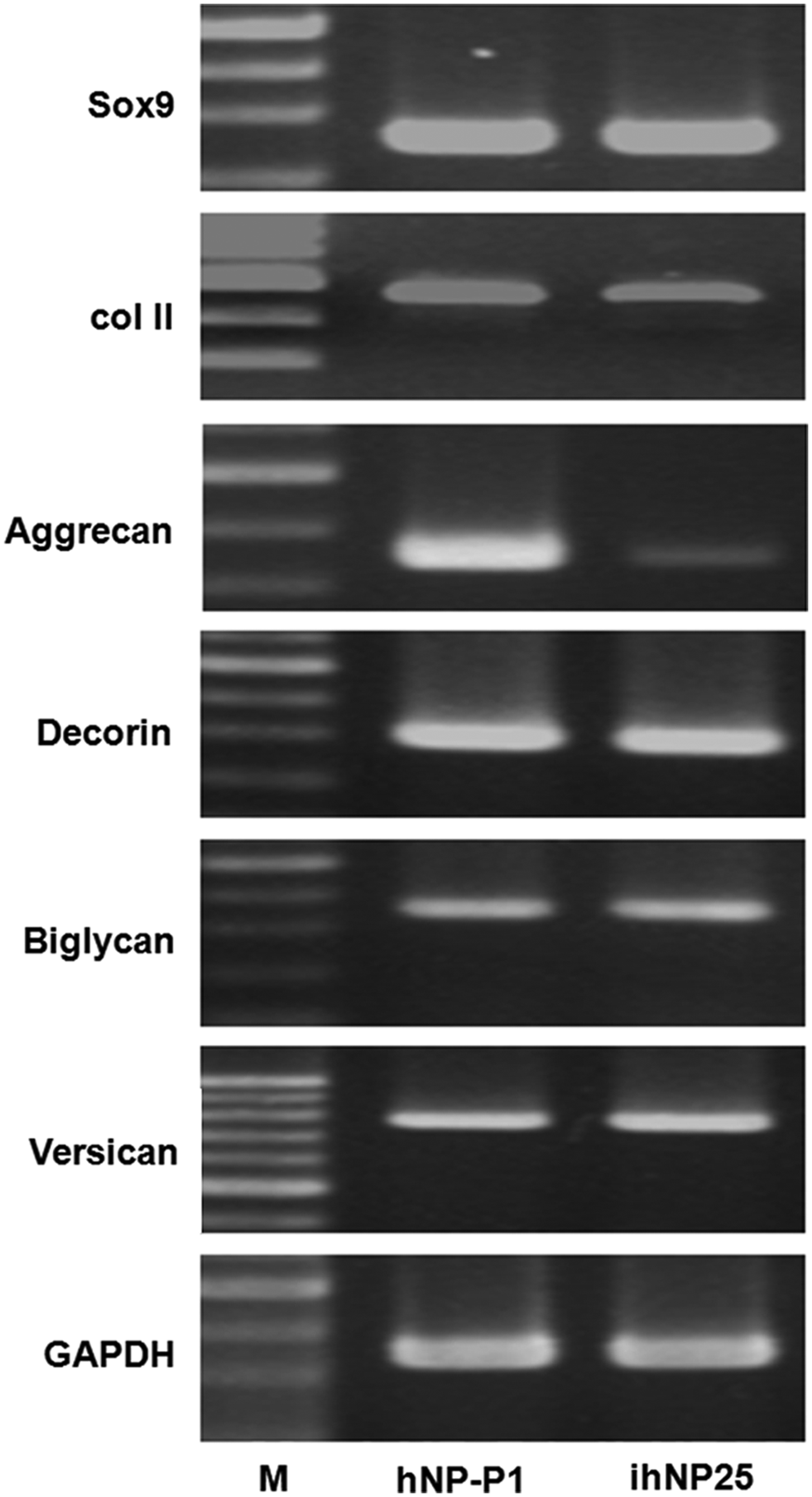
Comparison of chondrogenic-specific gene expressions between primary hNP and ihNP cells. Total RNA was isolated from hNP-P1 and ihNP25, and then subjected to RT-PCR analysis using primer pairs specific for Sox9, type II collagen (Col II), aggrecan, decorin, biglycan, and versican. M: 100 bp DNA ladder; glyceraldehyde 3-phosphate dehydrogenase (GAPDH): internal control.
Chondrogenic-specific protein and PG synthesis in ihNP cells
Western blotting analysis and Alcian blue staining were performed to evaluate chondrogenic-specific proteins expressions in ihNP cells. The chondrogenic protein phosphorylated SOX9 (p-SOX9) and Col II were highly expressed in ihNP25 cells compared to hNP-P1 cells (Fig. 4A). In addition, CD44, a specific hyaluronic acid receptor for NP development, was also highly expressed in ihNP25 cells.

Chondrogenic-specific protein expressions in parental hNP and ihNP (P25) cells.
Alcian blue staining for PG accumulation was then determined at 90% confluent phase. A higher intensive staining signal was detected in ihNP25 cells than in hNP-P1 cells (Fig. 4B).
The regenerative potentials of ihNP performed in 2D cell model
After establishing the immortalized ihNP cell line, which maintains the parental chondrogenic potential even more, subsequently, we determined the regenerative potential of ihNP25 as a platform for drug screening. Previously, we have created a 2D degeneration model and reported that the proinflammatory cytokines could decrease chondrogenic genes and enhance inflammatory molecules in chondrocytes. 15 We also reported that PRP, a growth factor complex extracted from platelets, could be ultilized as a regenerative reagent to restore chondrogenic degeneration.14–16 After treating ihNP25 with LPS and subsequently cotreated with PRP, semiquantitative RT-PCR analysis was then conducted (Fig. 5). The expressions of chondrogenic-specific genes, including SOX9, Col II (p<0.05), and aggrecan (p<0.001) were significantly diminished in ihNP25 cells after LPS treatment for 1 week, where the diminished chondrogenesis by LPS was recovered by PRP (Fig. 5A). In addition, the increased expressions of inflammatory markers by LPS, including IL-1β (p<0.05), TNF-α (p<0.05), and MMP-3 (p<0.001), were also significantly abolished by PRP (Fig. 5B).
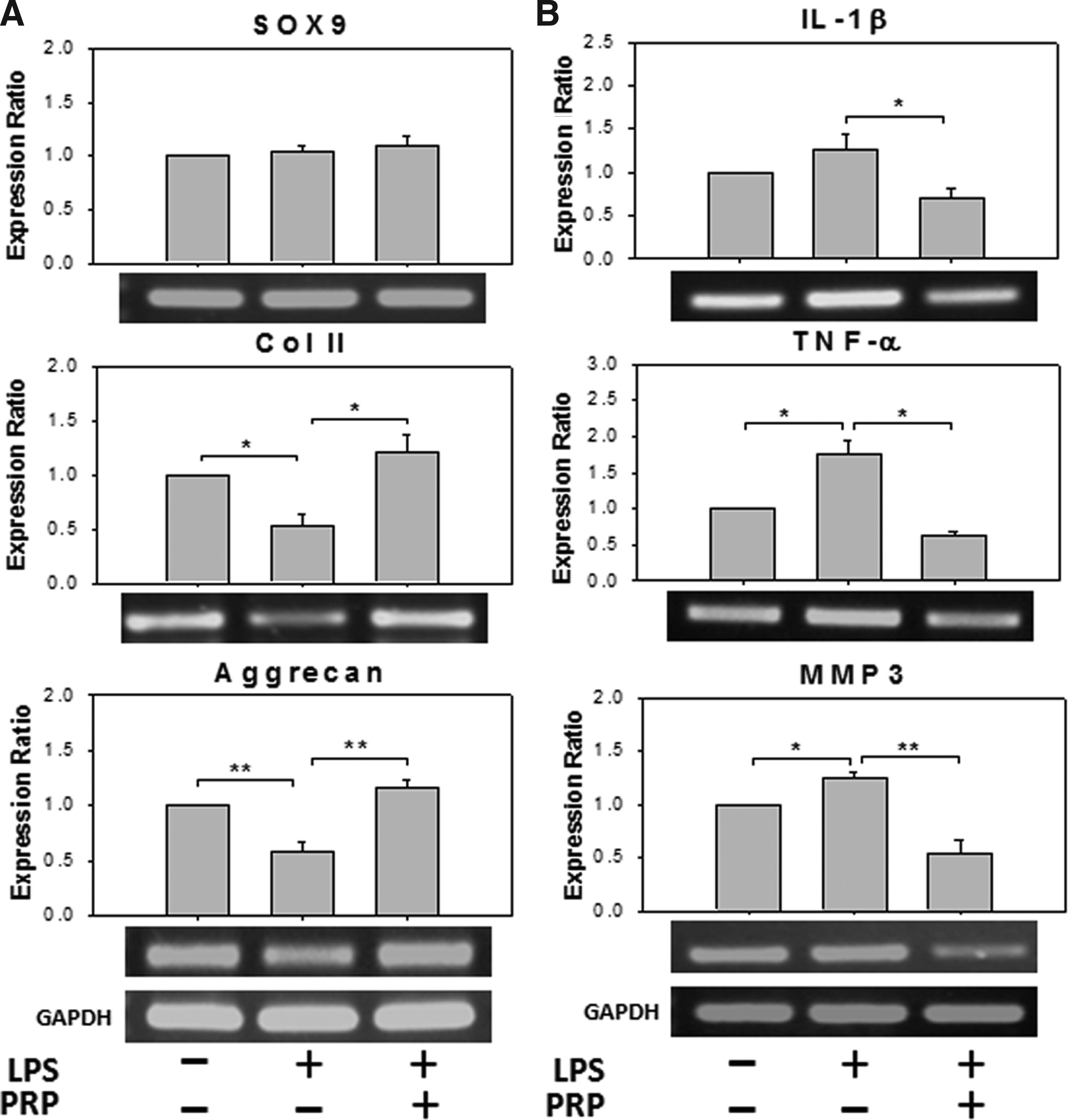
The regenerative potentials of ihNP cells in a 2D model. Semiquantitative RT-PCR analysis for chondrogenic-specific gene expressions
Therapeutic potentials of ihNP cells performed in a 3D degeneration model
After examining the regenerative potential of ihNP in the 2D model, we then used the 3D system, which has previously demonstrated the therapeutic potentials of chondrocytes in a collagen scaffold.12–16 The 3D culture embedded with ihNP25 were cultured in the presence of LPS [LPS(+)/PRP(−)], and then treated with PRP [LPS(+)/PRP(+)] (Fig. 6). The gross morphology changes were observed after 4 weeks of cultivation (Fig. 6a–c). After treated with LPS (Fig. 6b), the 3D construct showed a significantly deformed macromorphology, compared with control [LPS(−)/PRP(−)] (Fig. 6a) and PRP group (Fig. 6c). After treated with PRP, the 3D construct revealed a more elastic macromorphology (Fig. 6c). The LPS-induced cell death (Fig. 6e) was also then recovered by PRP (Fig. 6f). To examine the chondrogenic matrix accumulations, Col II and PG depositions were performed by IHC stain and Alcian blue stain, respectively (Fig. 6g–l). In the PRP-treated group, the Col II-positive signal was highly enhanced (Fig. 6i), especially in the region of cell proliferative layer, also indicating that PRP rescued the LPS-diminished PG in 3D constructs (Fig. 6l). However, in the LPS group without PRP, there showed completed depletion of Col II and PG (Fig. 6h, k) compared with the control group (Fig. 6g, j). In addition, the LPS-reduced expression of CD44 (Fig. 6n) was restored by PRP (Fig. 6o), indicating increased recovered cells acquiring chondrocytic phenotypes compared to the control group (Fig. 6m).

The therapeutic potentials of ihNP cells in a 3D model. Histological appearances of ihNP cells grown on 3D collagen constructs with the culture medium only
Discussion
IVD degeneration with aging results from both NP tissue and matrix degradation followed by cell apoptosis 28 Chondrocyte is the only cell type in cartilaginous tissue and critical to regulate the balance of tissue metabolism. We have previously indicated chondrogenic recovery of NP tissue by primary NP cells with growth factors, MSCs, and calcitonin.14,16,29 However, primary chondrocytes easily lose its chondrogenic phenotypes during in vitro cultures; therefore, an optimal chondrogenic recovery model with a promising cell line to study disc regeneration has become emergent. One of the most challenging approaches is to establish cell lines preserving chondrocyte phenotypes as a platform for cartilage regeneration. Immortalized cell lines could be extensively used for studies of pathophysiology30–32 and drug screening tools, 33 which allow mechanical investigations in disease and assessment of new therapeutic strategies. In addition, the immortalized chondrocyte model provides not only an unlimited resource, but also the stable chondrogenic phenotypes of chondrocytes for tissue engineering. Oncogenes have been introduced into primary chondrocytes for immortalization, however, disadvantages subsequently occurred. With the transduction of TERT, tumorigenic characteristics such as loss of contact inhibition and in vivo tumor formation were shown.23,34 Increased osteogenic markers, indicating the hypertrophy and senescence of chondrogenesis, were also associated with ectopic TERT expression in MSCs. 24 After immortalization with SV-40LT, the culture condition chondrocytes were limited, including the permissive temperature at 32°C and high cell density for maintaining chondrogenic phenotypes. 35 In addition, karyotypic instability and inadequate DNA repair were also noted.20,22
HPV encode oncoproteins, such as E6 and E7, can cooperatively facilitate cell immortalization, and extend the life span of animal primary cells by regulating cell cycle machinery. 36 We have previously established an HPV-16 E6/E7-immortalized articular chondrocyte hPi with innate physiological and neocartilage forming properties. 13 E6 and E7 proteins inactivate p53 and Rb proteins, respectively, and thus inhibit the downstream tumor suppressor genes. 37 The hPi cells have been utilized to study the stage-specific chondrogenesis of MSCs and the regenerative potential of PRP/collagen scaffold.12,15 The signaling pathways, stages, and molecular markers involved in chondrogenesis have been extensively investigated in our previous studies.11–16 The present study had attempted to create an ihNP cell line by transducing HPV-16 E6/E7 to permit studies of degenerative and regenerative modulations in NP chondrogenesis. The ihNP cells maintained morphological similarity with parental NP and stable expression of HPV-16 E6/E7 mRNA (Fig. 1). Rapid proliferation was found in ihNP cells by shortened population doubling time (53.16±2.63 h, Fig. 1C) and maintained for more than 50 passages (data not shown). To further determine cell cycle machinery in ihNP cells, the overexpression of HPV-16 E6 and E7, respectively, suppressed the phosphorylation of p53 and pRB, as well as p21 (Fig. 2A); however, ihNP can also be amplified without tumorigenicity (Fig. 2B). The cyclin E2 and A2, which trigger the transition of G1/S phase in cell cycle, 36 were also activated and leading to higher proliferative response in ihNP cells.
To study the physiology of NP, the preservation of original NP phenotypes of ihNP should be first determined. ECMs in disc tissue, including two major macromolecular components, collagens and PGs, are produced by NP tissue and critical for maintaining disc physiology. Col II is abundant in normal human NP tissue, accounting for up 80%. As the progression of NP degeneration decreased Col II, more fibrotic and immature collagens, including Col I and Col X were detected. 38 Furthermore, the transcription factor Sox9 is specifically implicated in chondrogenic differentiation and maintenance of the chondrocytic phenotype. 39 SOX9 also plays an important role in regulating Col II and aggrecan by binding their promoters that lead to chondrogenesis;40,41 conversely, a decrease in Sox9 expression has also been found to be associated with disc degeneration. 42 Aggrecan and versican, responsible for water-retaining properties of the disc tissues43,44 are large aggregating PGs in the IVD. Decorin and biglycan are small leucine-rich and interstitial PGs and react with specific regions of Col I and Col II. 44 These small PGs are thought to have an important organizational role in the assembly and also in the repair of ECM for the IVD during growth, maturation, aging, and after injury. 45 In addition, CD44 expressed on disc cells is critical for IVD development by anchoring hyaluronic acid and leading to the formation and expansion of the ECM. 46 The changes in ECM molecules would influence the matrix quality that contributes to IVD degeneration or regeneration. Limitation of monolayer cultures of primary chondrocytes was mainly due to the rapid loss of chondrocyte phenotypes, including a decrease of Col II and PG synthesis. These master genes and specific proteins of ihNP remained to be stably expressed even higher than those of parental NP (Figs. 3 and 4), suggesting that the ihNP well preserved chondrogenic phenotype of NP after immortalization.
The ihNP cells could maintain chondrogenic phenotypes and matrix accumulations during monolayer cultures. Subsequently, ihNP cells were treated with proinflammatory cytokines for catabolic effects or/and PRP for anabolic effects to address its regenerative and therapeutic potentials. To create the degeneration model in cartilaginous tissue, proinflammatory cytokines and chemonucleolytic agents have been utilized in our previous studies.15,16 The inflammation, degenerative responses, and ECM inhibition of disc cells induced by lipopolysaccaride (LPS) have also been reported.47,48 Our RT-PCR results demonstrated the decreased chondrogenic genes and increased inflammatory molecules by LPS treatment, resembling the in vivo disc degeneration process (Fig. 5). The degradation of IVD matrix was also induced by matrix metalloproteinases (MMPs), including MMP3. 7 In this study, the production of IL-1β, TNF-α, and MMP3 were increased after LPS induction (Fig. 5B). IL-1β was implicated to increase the production of MMPs and PG degradation in the IVD.49,50 In addition, PRP, a growth factor complex purified from total blood, has also been reported as a regenerative agent for disc degeneration and osteoarthritic cartilage.14–16 The LPS-induced ihNP degeneration and inflammation were also recovered by PRP treatment, demonstrated that the presence of PRP would be beneficial for IVD regeneration induced by inflammatory stressful conditions.
Furthermore, the therapeutic potentials of ihNP cells were verified in the 3D neocartilage model (Fig. 6). Chondrogenic phenotypes can be maintained by culturing cells in a 3D environment supporting spherical cell morphology, such as agarose gels 51 or biodegradable polyglycolic acid (PGA) scaffolds. 52 Our previous findings have also demonstrated that the collagen scaffold would preserve chondrogenic ECM synthesis of articular and NP chondrocytes in neocartilage.12–16 Moreover, the addition of proinflammatory cytokines would deform the neocartilage and mimic the cartilaginous degeneration. 15 LPS strongly inhibit not only cell survival (HE stain), but also chondrogenic matrix accumulations (IHC of Col II and Alcian blue). The expression of CD44, a transmembrane glycoprotein proposed as a receptor in the NP for the anchorage of hyaluronan and the development of IVD, 46 was also decreased. For disc repair, ihNP25 was subjected into neocartilage in a LPS-induced inflammatory environment to address its therapeutic potentials by PRP. The LPS induced cell death and inhibited specific matrix synthesis, were both rescued by PRP. In both 2D monolayer and 3D environments of ihNP25, the gene and protein expressions of a specific matrix (Col II, aggrecan) by inhibiting expression of inflammatory mediators (IL-1β and TNF-α) and matrix degrading enzymes (MMP3) have been demonstrated.
In conclusion, we have successfully established the chondrogenic recovery model with an immortalized hNP cell line, ihNP, by retroviral vector transduction of HPV-16 E6/E7. The ihNP cells preserved similar characteristics with chondrogenic phenotypes as parental NP cells and showed no tumorgenicity. For its therapeutic potentials, PRP stimulated a significant expression of chondrogenic markers and inhibited the expression of inflammatory mediators and matrix degrading enzymes in ihNP cells in both environments of 2D monolayer and 3D scaffolds, under LPS-induced inflammatory conditions. Hence, ihNP cells can be viewed as a cell resource and a powerful platform for investigating the NP regenerative mechanisms and physiology for drug discovery and development.
Footnotes
Acknowledgments
This work was supported by the National Science Council (NSC 99-2628-B-038-010-MY3, 101-2811-B-038-005, and 101-2314-B-038-023), the Department of Health (DOH, DOH101-TD-PB-111-NSC010), the DOH to Taipei Medical University- Center of Excellence for Cancer Research (TMU-CECR, DOH101-TD-C-111-008), Teaching Resource Center of Excellence, Taipei Medical University (A0051-7301), HealthBanks Biotech Co., Ltd., and Kooper Biotech Co., Ltd.
Disclosure Statement
No competing financial interests exist.