
Abstract
Cardiac progenitor cells (CPCs) are committed to the cardiac lineage but retain their proliferative capacity before becoming quiescent mature cardiomyocytes (CMs). In medical therapy and research, the use of human pluripotent stem cell-derived CPCs would have several advantages compared with mature CMs, as the progenitors show better engraftment into existing heart tissues, and provide unique potential for cardiovascular developmental as well as for pharmacological studies. Here, we demonstrate that the CAG promoter-driven enhanced green fluorescence protein (EGFP) reporter system enables the identification and isolation of embryonic stem cell-derived CPCs. Tracing of CPCs during differentiation confirmed up-regulation of surface markers, previously described to identify cardiac precursors and early CMs. Isolated CPCs express cardiac lineage-specific transcripts, still have proliferating capacity, and can be re-aggregated into embryoid body-like structures (CAG-EGFPhigh rEBs). Expression of troponin T and NKX2.5 mRNA is up-regulated in long-term cultured CAG-EGFPhigh rEBs, in which more than 90% of the cells become Troponin I positive mature CMs. Moreover, about one third of the CAG-EGFPhigh rEBs show spontaneous contractions. The method described here provides a powerful tool to generate expandable cultures of pure human CPCs that can be used for exploring early markers of the cardiac lineage, as well as for drug screening or tissue engineering applications.
Introduction
H
According to animal studies, mature CMs confer only a modest functional benefit when inserted into a damaged heart tissue.6,23,24 In contrast, cardiovascular progenitor cells (CVPCs) and cardiac progenitor cells (CPCs) may be better candidates for a functional engraftment and regeneration of a damaged myocardium.2,3,23,25 For drug-screening applications, the isolation of progenitors and a subsequent scale-up culturing may also be more suitable than the direct purification of the quiescent CMs. 26 Therefore, methods enabling the selection of progenitors with cardiac-restricted differentiation potential, but still having a high proliferative capacity, are of utmost importance.
Based on specific cell surface markers, CVPCs have already been isolated, but these progenitors gave rise to a mixed population, containing cardiac, endothelial, and smooth muscle cells. 27 Recently, a method has been developed for the generation and long-term maintenance of multipotent CVPCs, which gave rise to CMs with ∼80% efficiency. 26 CPCs, the descendants of CVPCs, are still able to proliferate but show only cardiac restricted differentiation potential. Methods producing CPCs from hPSCs have not yet been identified.
In our earlier work, we have established a system for genetically modifying hPSCs to enable a selective enrichment of human embryonic stem cell-derived cardiomyocytes (hESC-CMs). This was based on the use of an artificial CMV enhancer—chicken β-actin—rabbit β1-globin fusion (CAG) promoter providing exceptionally high levels of enhanced green fluorescence protein (EGFP) expression in differentiated CMs. 28 The CAG is a constitutively active promoter, providing EGFP expression in all types of tissues, however with different expression intensities. This non-typical behavior also offers the advantage to select for transgene expressing undifferentiated hESCs after transfection. The CAG promoter, most probably through multiple interactions with cardiac-specific transcription factors, strengthens the expression of EGFP in differentiated CMs 28 and testing of potential regulatory binding sites is one of the main objectives of our related patent application (PCT/IB2008/054238). However, these terminally differentiated CMs show very poor reaggregation capacity and low survival rate after sorting; thus, for the generation of pure CM populations for tissue engineering applications, the use of CPCs may be more suitable.
In the present study, we demonstrate that the applied CAG promoter drives the expression of EGFP at an exceptionally high level in the hESC-derived CPCs. These cardiac progenitors represent an earlier stage of differentiation than CMs, but are more restricted in their differentiation potential than CVPCs. CPCs isolated based on the exceptionally high EGFP express cardiac-specific genes, are able to proliferate, and generate spontaneously contracting cardiac bodies, containing more than 90% of mature CMs. Our results suggest that this method of purification and subsequent differentiation of CPCs is a promising strategy to obtain pure populations of CMs even for tissue engineering applications. Moreover, this system provides a unique opportunity to profile the expression of markers; for example, cell surface proteins during early cardiac differentiation.
Materials and Methods
hESC culture and differentiation
The previously established CAG promoter-driven EGFP expressing HUES9-CAG-EGFP and BG01V-CAG-EGFP hESC lines were used in all experiments.
28
CM differentiation was initiated via embryoid body (EB) formation as previously reported,
27
with some modifications. For further information, see Supplementary Fig. S1A and Supplementary Materials and Methods (Supplementary Data are available online at
Flow cytometry and cell sorting
Single-cell suspensions from undifferentiated HUES9-CAG-EGFP colonies and HUES9-CAG-EGFP EBs were labeled with monoclonal antibodies against human SSEA4, CD166, CD106, SIRPA, anti-Troponin I, and anti-mouse Sca-1 antibody for specific labeling of mouse feeder cells. Control staining was performed with isotype-matched control mAbs and fluorochrome-conjugated secondary antibodies. Dead cells were gated out by 7AAD staining. Mock-transfected HUES9 cells were measured to set the level for EGFP-positivity, and the HUES9-CAG-EGFP EBs were sorted into low- and high-signal intensity fractions (FACSAria High Speed Cell Sorter)—for detailed information, see Supplementary Materials and Methods.
Fluorescent plate reader measurements
For fluorescent plate reader measurements, a VICTORX3 2030 Multilabel Reader (Perkin Elmer) was used. For detection of EGFP fluorescence excitation, a wavelength of 490 nm and an F535 emission filter were used. For propidium iodide (PI) staining, the re-aggregated EBs (rEBs) were fixed with methanol on ice for 15 min and, after a washing step, they were stained with PI (excitation at 540 nm, and emission at 660 nm). Duration of image acquisiton was ∼1 min, and each repetition was started at 30 s after completion of the previous scan. Three data sets were used to calculate the average fluorescence intensity levels normalized to the background fluorescence level in the case of each rEB (n=6, from two independent sorting experiments).
Immuncytochemistry
For immunostaining, rEBs were plated onto gelatin-coated eight-well chambers, and staining was performed as previously described. 28 For differentiation, markers anti-Troponin I, βIII-Tubulin, smooth muscle actin (SMA), and alpha-fetoprotein (AFP) primary antibodies were applied—for more details, see Supplementary Materials and Methods. DAPI was used for nuclear staining, and the samples were examined by an Olympus FV500-IX confocal laser scanning microscope. At least two independent samples were used in each measurement.
Real-time polymerase chain reaction analysis
Total RNA was isolated from cells using TRIzol™ Reagent following the manufacturer's instructions. cDNA samples were prepared from 0.2 μg total RNA using the Promega Reverse Transcription System Kit as specified by the manufacturer. For real-time quantitative polymerase chain reaction (PCR), the following Pre-Developed TaqMan® assays were purchased from Applied Biosystems: NANOG as undifferentiated stem cell marker; ISL1, TBX5, ALCAM, NKX2.5, GATA4, and TNNT2 as markers of cardiac differentiation, MYL2 as ventricular-, MYL7 as atrial-, and HCN4 as nodal subtype specific marker; and P0 ribosomal protein as endogenous control for quantification. Real-time quantitative PCR (qPCR) analyses were carried out using the StepOne™ Real-Time PCR System, according to the manufacturer's instructions. The fold change of mRNA in experimental and control cells was determined using the 2−ΔΔCt method. Relative mRNA levels were presented as mean values±SEM of three independent experiments. Levels of significance were calculated by the two-tailed Student t-test.
Results
Directed differentiation of human pluripotent cells
CM differentiation from the human pluripotent cells was initiated via EB formation in suspension, as previously reported, 29 with some modifications (Supplementary Fig. S1A). Differentiation into cardiac cell types, occurring after day 6, was identified by measuring the mRNA expression of ISL1, marking the progenitors of the secondary heart field in the early embryo, 27 TBX5 and NKX2.5, transcription factors required for cardiac development, TNNT2, a gene encoding cardiac Troponin T, and the activated leukocyte cell adhesion molecule (ALCAM/CD166). ALCAM expression identifies hESC-derived CMs, 19 and it has been recently shown that CMs isolated from human heart tissue also express this cell surface antigen. 30
As shown in Figure 1A, we found that by day 2 of differentiation, a rapid down-regulation of the NANOG mRNA was followed by a transient up-regulation of brachyury mRNA expression. ISL1 expression was up-regulated on day 6, preceding the up-regulation of TBX5 and NKX2.5 expression (Fig. 1B). TNNT2 mRNA became first detectable in the day 6 sample (Fig. 1B), while the up-regulation of TNNT2 mRNA occurred around day 14, simultaneously with the emergence of spontaneous contractile activity in the differentiating culture. Up-regulation of ALCAM mRNA levels overlapped with the up-regulation of ISL1.
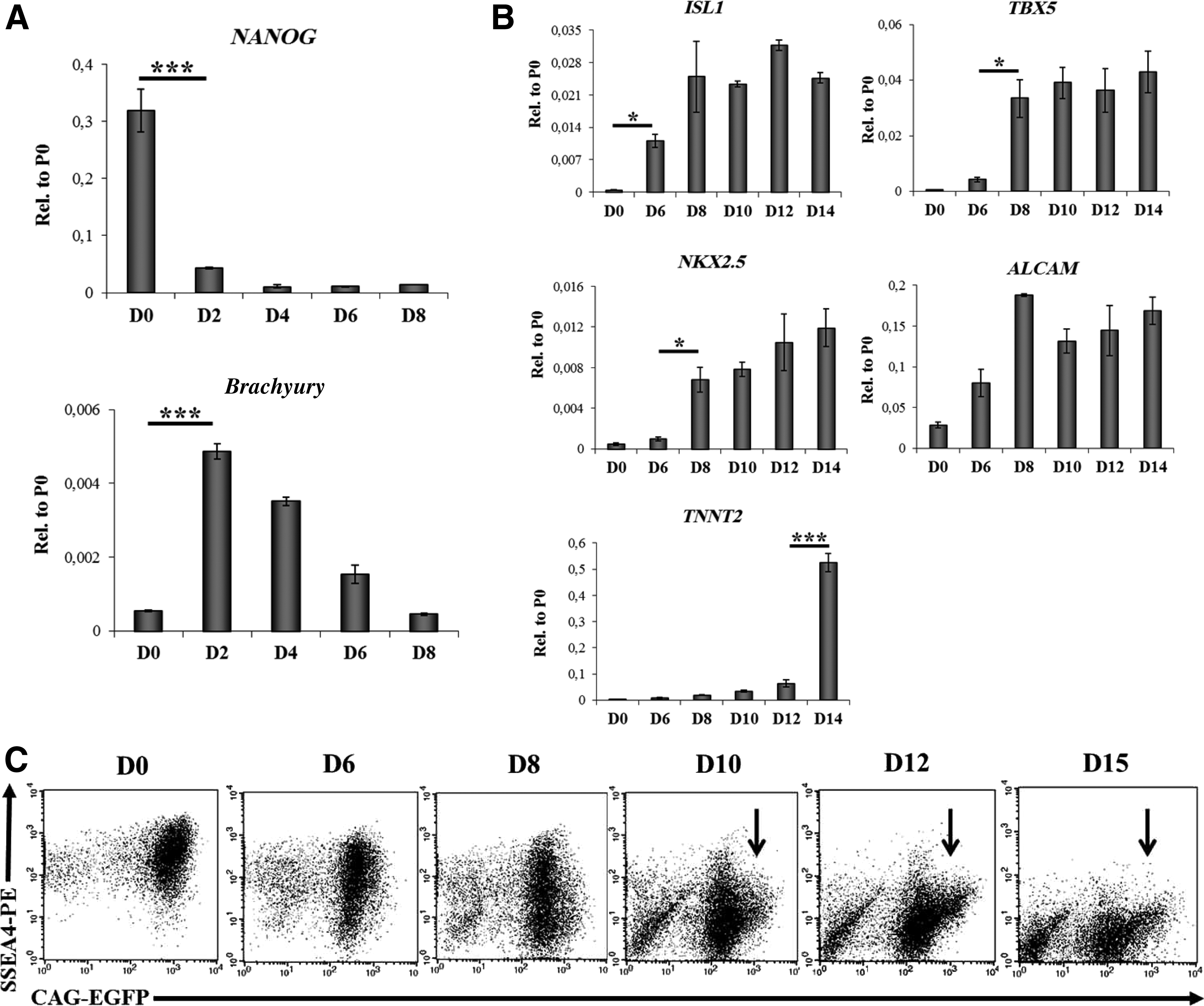
Induction of HUES9-CAG-EGFP cell differentiation.
As shown in Figure 1C, in the HUES9-CAG-EGFP cells, a well-measurable EGFP expression could be detected by flow cytometry in the pluripotent state (day 0), along with a high-level SSEA4 cell surface expression, as reported earlier. 28 A further analysis of CAG-EGFP and SSEA4 expression during cell differentiation revealed a concomitant early decrease in these expression levels (day 2–6). However, between days 10 and 15, the formation of a subpopulation expressing EGFP at a very high level (CAG-EGFPhigh, indicated with black arrows on Fig. 1C), and negative for SSEA4 expression was observed. This CAG-EGFPhigh cell population appeared already 4 days before the onset of the spontaneous beating activity (day 14) in the differentiated cell cultures. To evaluate the suitability of the CAG-EGFP method for an hESC line other than HUES9, differentiation kinetics of the BG01V hESC cells, stably expressing the CAG-EGFP construct, was also examined (see Supplementary Fig. S1B).
The CAG-EGFPhigh population is enriched in CPCs
At day 6, the EBs were plated onto gelatin-coated plates (see Materials and Methods). In order to assess whether the CAG-EGFP expression levels can be used to select the CPCs, cell sorting was performed from the trypsinized cultures of differentiating HUES9-CAG-EGFP cells on day 10 and 12 (Fig. 2A). Day 10 is the earliest time point at which the CAG-EGFPhigh cells are already SSEA4 negative (Fig. 1C), while sorting on day 12 still precedes the onset of spontaneous beating activity and significant up-regulation of TNNT2 expression occurring only later, at day 14. Therefore, this latter separation can also result in isolation of CPCs, however, with a more mature phenotype. As shown in Figure 2A, four different subpopulations of the cells, based on the GFP intensities, were identified by flow cytometry, and two separate cell populations (CAG-EGFPhigh and CAG-EGFPlow) were analyzed for marker expression. The sorted CAG-EGFPhigh and CAG-EGFPlow subpopulations were re-analyzed by repeated FACS measurements to establish sorting purity (see Supplementary Fig. S2). Cells expressing very low levels of EGFP, and probably losing the transgene construct, were not included in the further studies (see Fig. 2A).
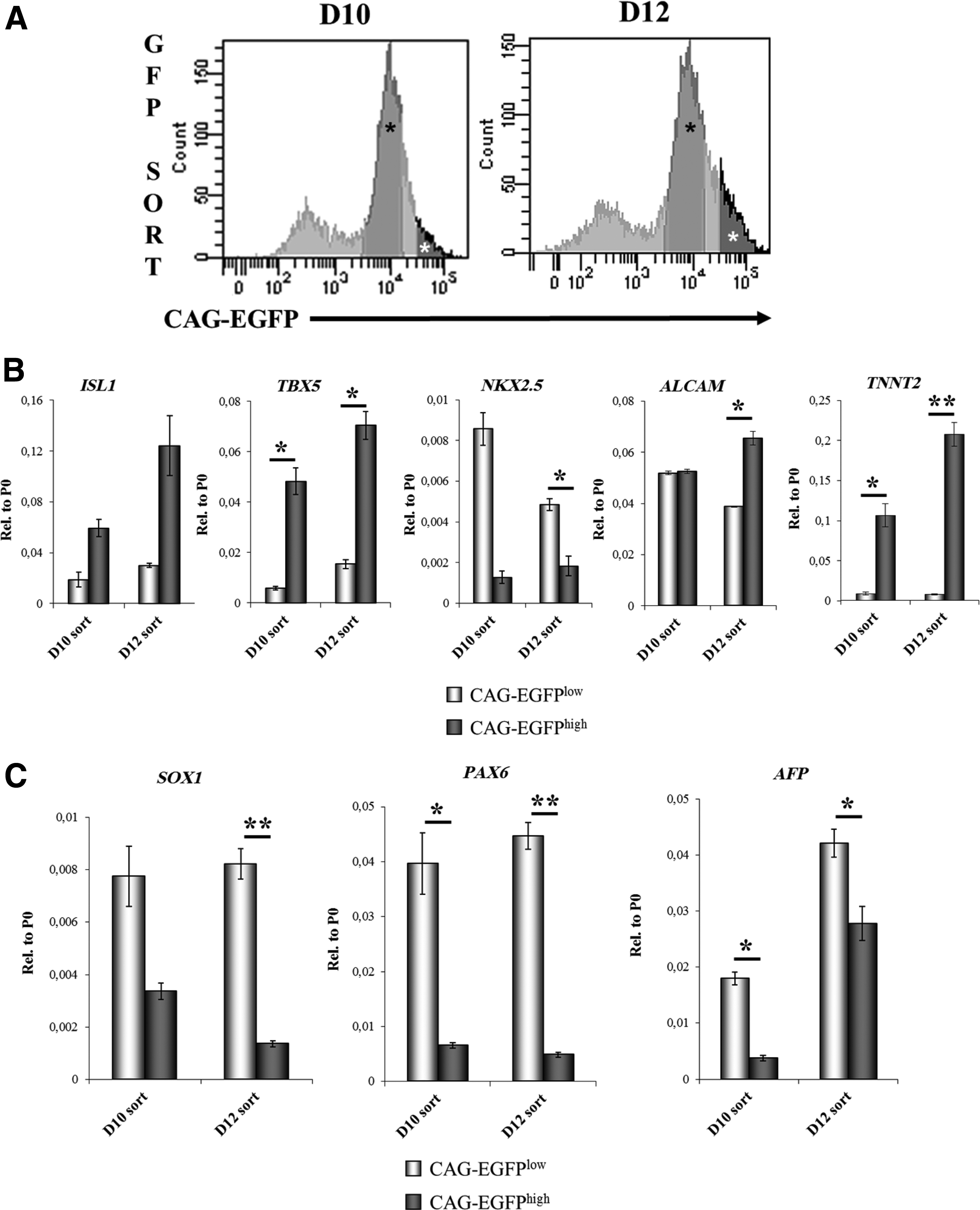
The subpopulation with highest EGFP fluorescence intensity is enriched in cardiac progenitor cells.
qPCR analysis revealed that CAG-EGFPhigh cells expressed higher levels of ISL1, TBX5, and TNNT2 than the CAG-EGFPlow cells, in samples obtained at both day 10 (D10) and day 12 (D12) (Fig. 2B). Surprisingly, NKX2.5 mRNA levels were higher in CAG-EGFPlow than in CAG-EGFPhigh cells, at both sortings, most probably due to the overrepresentation of an NKX2.5low subtype of CPCs or very early-stage progenitors in the CAG-EGFPhigh fraction. Moreover, the presence of cardiac cells in the CAG-EGFPlow cells cannot be excluded either (Supplementary Fig. S3). Expression levels of the NKX2.5 transcripts slightly increased from day 10 to 12 in the case of CAG-EGFPhigh cells; however, in the case of CAG-EGFPlow cells, a robust decrease was observed, presumably due to the higher proliferation rates of non-cardiac cells present in this fraction. Distribution of cardiac-specific mRNA levels between BG01V fractions sorted on day 12 were similar to that observed in the case of HUES9 cells (Supplementary Fig. S4).
ALCAM mRNA levels were similar in the CAG-EGFPhigh and CAG-EGFPlow cell populations at day 10, but 2 days later, ALCAM expression levels showed an increase in the CAG-EGFPhigh fraction (Fig. 2B). In contrast to the cardiac markers, the early neuronal markers SOX1 and PAX6, as well as the early endoderm marker AFP were expressed at higher levels in the CAG-EGFPlow cell fraction (Fig. 2C).
The findings on ALCAM mRNA expression were reinforced by the kinetic analysis of ALCAM and CAG-EGFP expression at the protein level. ALCAM protein positive cells were first detected at day 8, but the ALCAM+ fraction did not include CAG-EGFPhigh cells until day 12; whereas a clear co-expression of the two markers was first observed on day 14 (Fig. 3A).
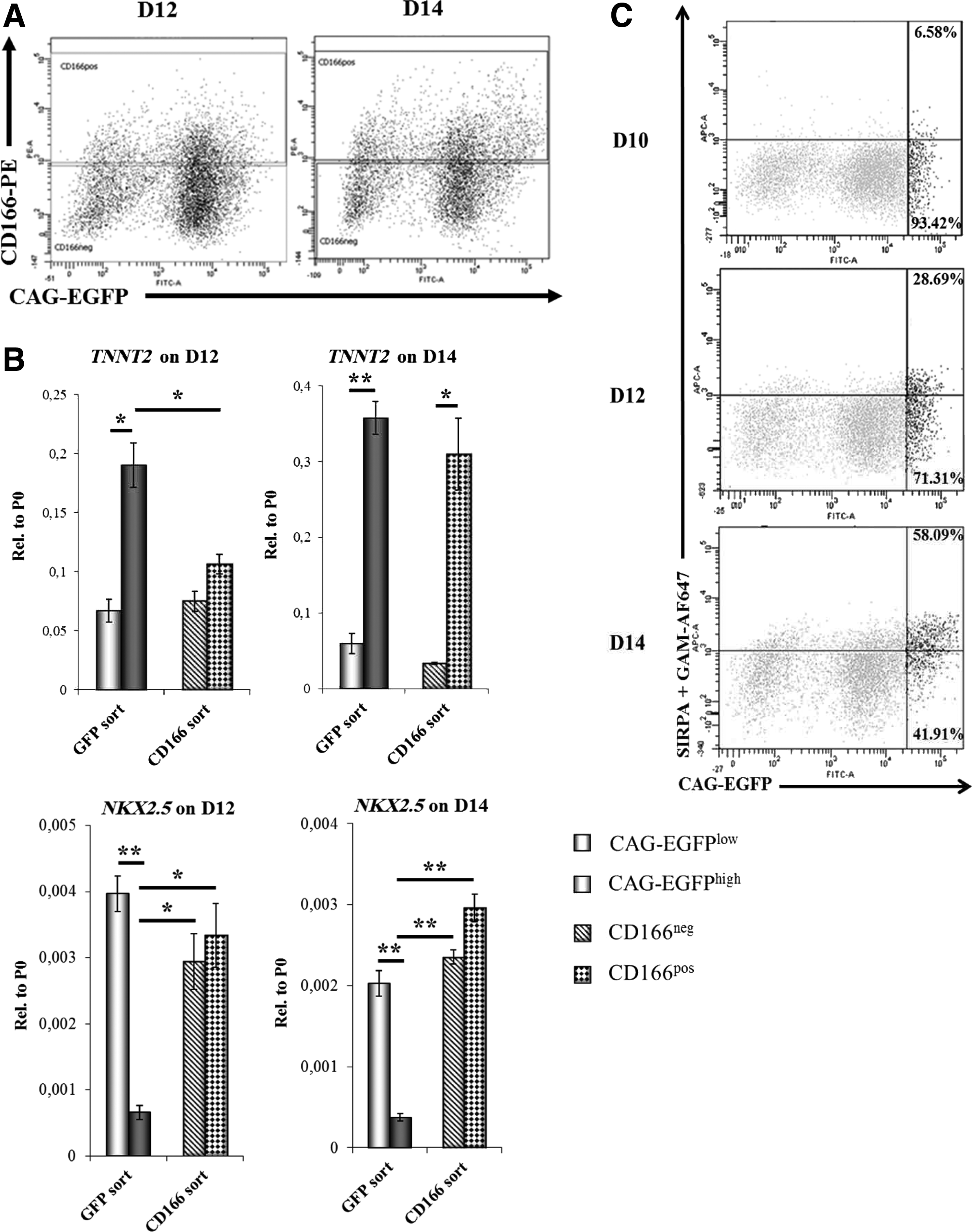
Analysis of the cardiac progenitors obtained by the CAG-EGFP separation system and those by the presence of suggested early cardiomyocyte markers.
In order to verify the CAG-EGFP expression-based selection of CPCs (D12 cells) and early CMs (D14 cells), we have compared our system with surface antigen-based selection systems previously reported in the literature.18–20,31 Half of the cell suspension obtained by trypsinization from adherent EBs at day 12 or 14, respectively, was sorted into ALCAM-positive (ALCAMpos) and ALCAM-negative (ALCAMneg) fractions (Fig. 3A), while the other half was sorted into CAG-EGFPhigh and CAG-EGFPlow fractions (as shown in Fig. 2A). On day 12, only the CAG-EGFP-based selection resulted in samples with distinct TNNT2 mRNA levels; while the ALCAMpos and ALCAMneg samples did not differ significantly from each other either in TNNT2 or in NKX2.5 mRNA expression levels (Fig. 3B). The difference in TNNT2 mRNA expression levels between CAG-EGFPlow and CAG-EGFPhigh samples further increased from day 12 to 14. By day 14, the ALCAMpos and ALCAMneg populations also became more distinct, based on their TNNT2 and NKX2.5 mRNA expression levels (Fig. 3B). CAG-EGFPhigh cells expressed significantly lower levels of NKX2.5 mRNA than all the other samples, irrespective of the day of sorting, raising the possibility of an NKX2.5low subtype of cardiac precursor cells, marked by the CAG-EGFP system.
Expression of SIRPA, a marker of cardiac precursor cells, 18 and vascular cell adhesion molecule-1 (VCAM-1), a marker of early CMs, 20 was also examined in the CAG-EGFP-based differentiation system. Emergence of the CAG-EGFPhigh cells (D8–10, see Fig. 1C) clearly preceded the formation of the SIRPA positive population (first observed at D12, see Fig. 3C), but during the course of differentiation, CAG-EGFPhigh cells started expressing SIRPA (28.69% of the CAG-EGFPhigh cells were SIRPA+ on D12, and 58.09% were SIRPA+ on D14, respectively), suggesting ongoing CM differentiation within the CAG-EGFPhigh population (Fig. 3C). As previously reported, 31 expression of VCAM-1 only partially overlapped with SIRPA (Supplementary Fig. S5). However, SIRPA-positive CAG-EGFPhigh cells (black dots) started expressing VCAM-1 by day 14 (see Supplementary Fig. S5).
CAG-EGFPhigh cardiac progenitors give rise to mature CMs
In order to further examine the potential of the CAG-EGFPhigh subpopulation to yield mature cardiac cells, the isolated CAG-EGFPhigh and CAG-EGFPlow cells were plated in suspension cultures on either day 10 or 12, and maintained for several weeks in the presence of 10% bovine serum (Fig. 4A). Under these conditions, isolated cells re-aggregated and formed EBs within 1 day after plating. Cells sorted on day 10 exhibited better re-aggregation properties than those sorted on day 12. In order to examine the growth of the rEBs, fluorescence imaging (Fig. 4B) and fluorescence plate reader measurements were performed at 3 and 20 days after cell sorting. CAG-EGFPhigh cells from the day 10 sort exhibited greater proliferative capacity than CAG-EGFPhigh cells isolated 2 days later. This observation was confirmed not only by plate reader measurements, based on EGFP fluorescence (Fig. 4C), but also by PI staining of fixed rEBs (Fig. 4D). Fluorescence intensities of the PI staining of 3 day-old rEBs sorted on day 10 (D10 sort) and day 12 (D12 sort) were similar, corresponding to the identical cell numbers (30,000 cells/well).
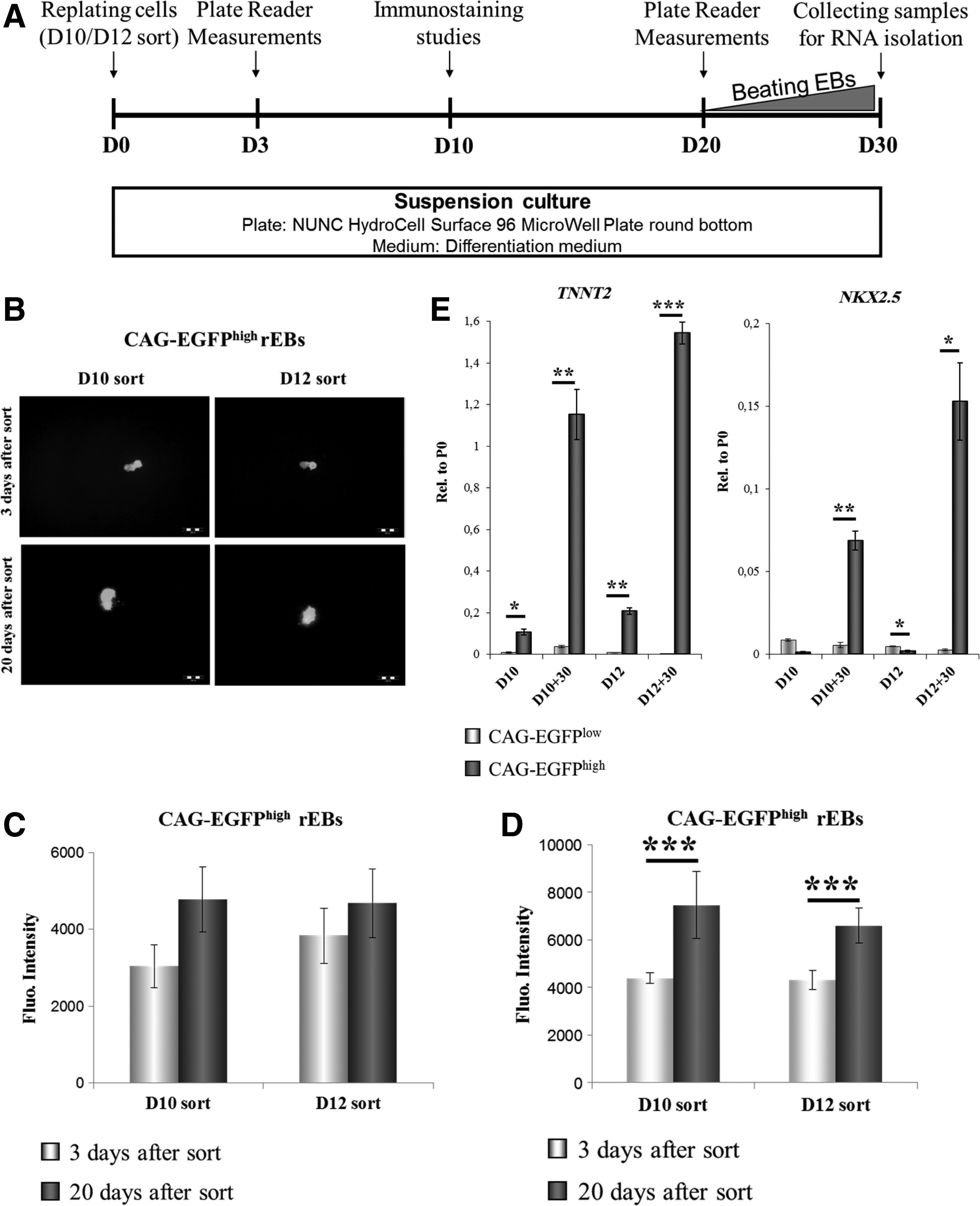
HUES9-CAG-EGFPhigh cardiac progenitor cells are able to proliferate and give rise to mature cardiomyocytes.
To enhance survival and reaggregation ability of the sorted D12 CAG-GFPhigh cells, the culture conditions were optimized (Szebenyi, unpublished experiments). The proliferation features of the CAG-EGFPhigh and CAG-EGFPlow cells were different, as CAG-EGFPlow cells generated perfect spheroids; whereas CAG-EGFPhigh rEBs seemed to grow along a certain axis (Fig. 4B and Supplementary Fig. S6).
Spontaneous beating activity was first detected at 25 days after reculturing the isolated cells. Contracting rEBs arose only from the CAG-EGFPhigh but not from the CAG-EGFPlow cells (Supplementary Movies S1 and S2).
qPCR analyses revealed that after 30 days in culture, the CAG-EGFPhigh rEBs expressed significantly higher levels of TNNT2 and NKX2.5 than the CAG-EGFPlow rEBs (Fig. 4E). Moreover, cardiac-specific mRNA levels of the CAG-EGFPhigh rEBs (D10+30 CAG-EGFPhigh and D12+30 CAG-EGFPhigh) were several fold higher than those of the initially isolated CAG-EGFPhigh cells (D10 CAG-EGFPhigh and D12 CAG-EGFPhigh), implying maturation of the CPCs into CMs. The low levels of NKX2.5 in the samples harvested on day 10 and 12, and the subsequent up-regulation of NKX2.5 expression in CAG-EGFPhigh rEBs also indicate that the cells expressing EGFP at a very high level in the early phase of differentiation can give rise to mature CMs.
In order to examine the presence or absence of endoderm and neuronal cell types in the rEBs, we have performed an immuno-histochemical analysis of the neuronal marker βIII-Tubulin, as well as the early endoderm marker AFP in rEBs, generated from CAG-EGFPhigh or CAG-EGFPlow cells after cell sorting on day 12. The floating rEBs were seeded out on eight-well confocal microscopy chambers at 3 days before immunostaining. CAG-EGFPhigh and CAG-EGFPlow rEBs exhibited different adhesion properties: CAG-EGFPlow rEBs adhered and spread out well on a gelatin-coated surface, whereas CAG-EGFPhigh rEBs showed poor adhesion ability, and a short trypsinization procedure was needed to achieve better adhesion. In CAG-EGFPlow rEBs, we found an appreciable presence of neuronal committed cells with high βIII-Tubulin expression, while this expression was absent in CAG-EGFPhigh rEBs (Fig. 5A/1). A few AFP positive cells (presumably responsible for the low AFP mRNA expression, as seen in Fig. 2C) were detected in all rEBs examined (Fig. 5A/2). Troponin I expression was only present in CAG-EGFPhigh rEBs (Fig. 5A/3a) and in cells around them (Fig. 5A/3b), presumably generated by detachment from the CAG-EGFPhigh rEBs during trypsinization. Platelet endothelial cell adhesion molecule (PECAM)-positive endothelial cells could not be found either in CAG-EGFPhigh or in CAG-EGFPlow rEBs; while in the unsorted differentiation culture, some areas stained positive for PECAM (see Supplementary Fig. S7). A low number of cells around the CAG-EGFPhigh rEBs stained positive for SMA, and cells in the areas where CAG-EGFPlow rEBs spread out and formed monolayers also stained positive for SMA (Fig. 5A/4).
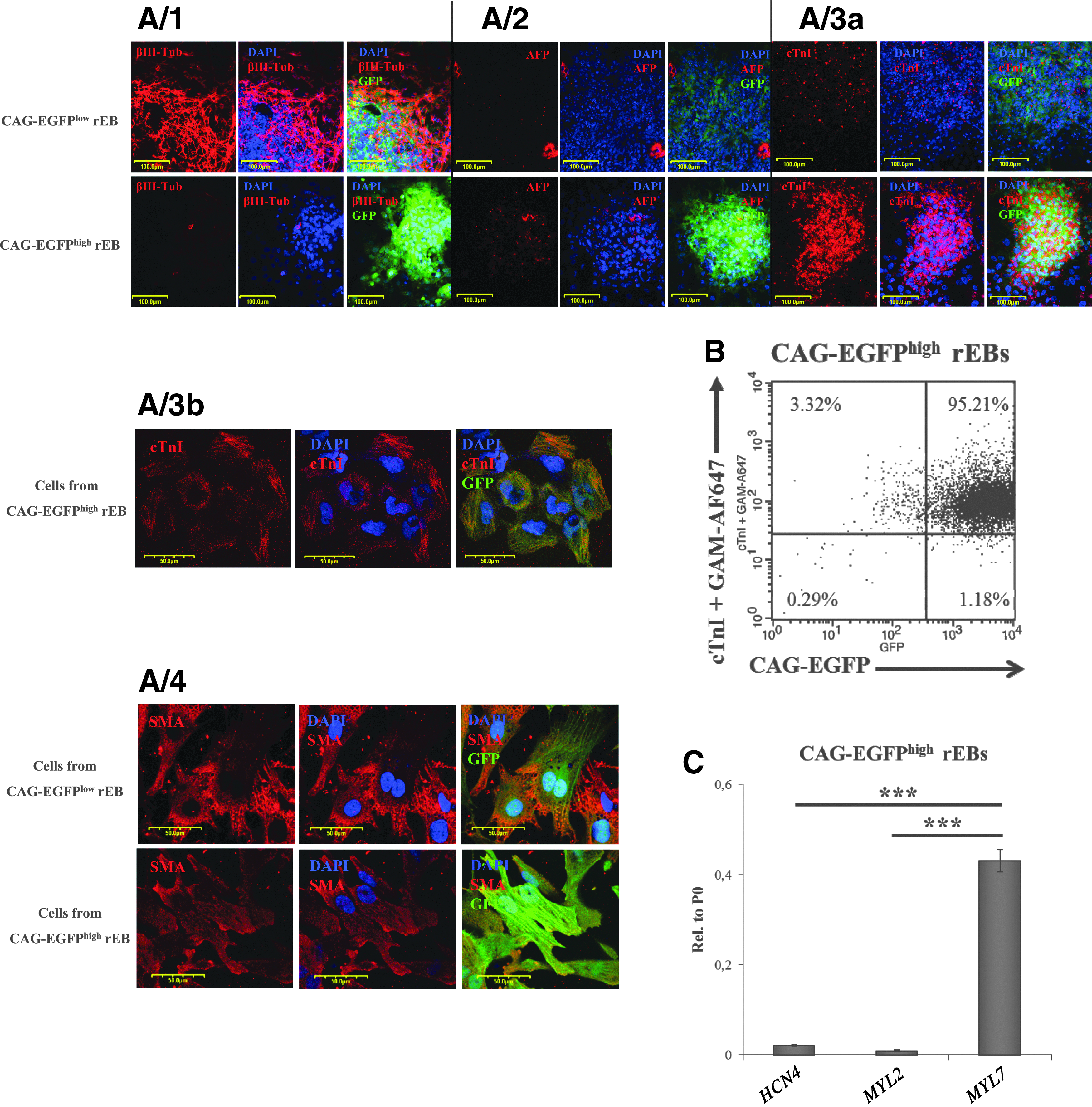
CAG-EGFPhigh rEBs are cardiac bodies predominantly consisting of cardiac cells.
In order to assess the efficiency of CAG-EGFP-based enrichment for CMs, Troponin I expression levels of the CAG-EGFPhigh rEBs were examined by flow cytometry (Fig. 5B). We found that more than 98% of the cells expressed Toponin I.
In order to determine the cardiac cell subtypes in CAG-EGFPhigh rEBs, MYL2, MYL7, and HCN4 mRNA expression was analyzed by qPCR (Fig. 5C). HCN4, the earliest expressed nodal cell marker, and MYL2, a marker of ventricular CMs, were only present at very low levels; whereas MYL7, the marker of atrial CMs, showed high-level expression in these samples. These data indicate that CMs derived from the isolated CAG-EGFPhigh cells are mostly of atrial types. This observation is in good agreement with the finding that in the unsorted differentiation cultures, some of the Troponin I-positive areas did not express EGFP at high levels (see Supplementary Fig. S3), which may indicate a differential regulation of the CAG-EGFP expression in various CM types.
Discussion
The first human ES-cell-derived therapy was based on hESC-derived oligodendrocyte precursors, 32 as these oligodendrocyte precursors differentiated into mature oligodendrocytes after transplantation. Using precursor cells seemed the best choice, as undifferentiated hESCs, besides the differentiation into the desired cell population, formed teratomas, 33 while transplantation of differentiated motor neurons showed only limited functional recovery. 34 Cardiac progenitors have been applied in transplantational studies as well, either in the form of cell suspension 35 or as engineered cardiac patches 2 to improve the modest functional benefit of existing transplantation strategies.
Progenitor cells used in tissue engineering applications can be derived from cardiac surgical waste, from fetal heart tissue, 3 or from in vitro differentiation of hPSCs.2,35 However, human PSC-derived progenitors are easy to access, can be isolated at different stages of commitment, and retain self-renewal capability and differentiation potential during long-term maintenance. 26
Steps during in vitro cardiac differentiation include the induction of mesoderm-type cells, followed by the formation of CVPCs, which are able to give rise to CMs, smooth muscle cells, and endothelial cells. In the next stages, the so-called CPCs become more restricted in their differentiation potential and give rise solely to CMs, but still possess some remaining proliferation capacity. This proliferative capacity rapidly declines during maturation, while the CMs emerging at the end of the in vitro differentiation are still immature in their electrical and contractile properties, and resemble more a fetal-like CM phenotype.36–38 However, transplantation of hESC-derived CMs into the infarct area has only yielded transient benefits, presumably due to the limited proliferative capacity of differentiated CMs and a poor electrical coupling.
In this study, we investigated the potential use of the CAG-EGFP system to identify CPCs, applicable to generate mature CMs. The CAG-EGFP system was earlier described as a “double feature” reporter, enabling the identification of both undifferentiated hESCs and differentiated CMs, based on exceptionally high expression levels of the CAG promoter-driven EGFP in these cell types. 28
In our previous work, we used the method of EB formation for differentiation but this method resulted in very low CM generation efficiency. Therefore, we adopted a protocol to specifically induce mesoderm formation and to support the increased generation of progenitors of the cardiac lineage with the addition of cytokines such as Activin A and BMP4. In our system, cells expressing the CAG-EGFP at very high levels could be separated in an early phase of the CM differentiation, occurring at 2–4 days before the development of a spontaneous beating activity and a concomitant up-regulation of TNNT2 expression.
QPCR analysis confirmed that the CAG-EGFPhigh fraction was enriched in progenitors of the cardiac lineage. The isolated CAG-EGFPhigh cells were able to form rEBs, in which the cells still had a proliferation potential and gave rise to mature cardiomycytes. Based on immunocytochemical, flow cytometry and qPCR analysis, we concluded that the CAG-EGFPhigh cells formed cardiac bodies, as more than 90% of the cells became cTnI positive. In addition, more than 30% of the cardiac bodies showed spontaneous beating activity after 20 days in culture. This relatively late onset of spontaneous beating activity may be caused by the time required to establish firm cell connections between the dissociated progenitors and the establishment of the electrochemical syncytium after re-aggregation. All these results strongly suggest that by this method, early CPCs yielding mature CMs can be isolated.
We also performed a detailed, comparative investigation of the CAG-EGFP separation system and several other recently published methods, suggesting that the cell surface markers ALCAM, VCAM-1, and/or SIRPA could be applied for isolation of early CMs and cardiac precursors.
We found that from among the four examined markers, the up-regulation of the CAG-driven EGFP expression could be first observed, days before the appearance of other cardiac lineage-specific cell surface proteins. In the course of differentiation when the CAG-EGFPhigh subpopulation appeared, NKX2.5 mRNA was not yet up-regulated, and neither SIRPA nor VCAM1 or ALCAM positivity could be detected—all these markers became measureable only later in the course of cell differentiation. However, low expression of NKX2.5 in CAG-EGFPhigh cells also lasted when the ALCAM-based selection already provided ALCAMpos cells with increased NKX2.5 mRNA level, suggesting the existence of an NKX2.5low CAG-EGFPhigh CPC population. It has been shown in NKX-deficient zebrafish embryos that NKX gene products may play direct roles in maintaining ventricular gene expression and repressing atrial gene expression.39,40
Based on these findings, we suggest that cells characterized by high CAG-EGFP expression are cardiac progenitors in a very early stage, preceding those stages defined by the reported cell surface markers. Moreover, the finding that CAG-EGFPhigh cells give rise to CMs with more than 90% efficiency without any special lineage-patterning culture conditions supports the hypothesis that CAG-EGFPhigh cells represent a population of fully committed CPCs. In addition, low NKX2.5 mRNA levels of the CAG-EGFPhigh CPCs and high MYL7 mRNA levels of the CAG-EGFPhigh rEBs suggest an atrial subtype specificity of the CAG-EGFP system. A detailed characterization of the CAG-EGFPhigh CPCs is needed to further examine these potential atrial subtype-specific CPCs.
We conclude that this method enables the isolation of human CPCs, expressing only low levels of NKX2.5 mRNA; while further differentiation of these CPCs results in a pure population of CMs, mostly containing atrial-type myocytes. The use of CAG-EGFPhigh CPCs in tissue engineering applications, aiming at supporting the regeneration of atrial tissue, may lead to enhanced engraftment and repopulation into the host tissue. In addition, the CAG-EGFP system provides a unique opportunity to examine the selective markers for hESC-derived CPCs to enable the isolation of genetically unmodified CPCs for tissue engineering and potential further therapeutic applications.
Footnotes
Acknowledgments
The authors appreciate the gift of the HUES9 cell line from Dr. Douglas Melton, HHMI. The authors are grateful to Beáta Haraszti for the excellent technical assistance rendered and to András Füredi for useful suggestions. This work was supported by the Hungarian Scientific Research Fund (grant number NK83533) and by the National Development Agency (grant numbers KTIA_AIK_12-1-2012-0025, KMR_12-1-2012-0112).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.