
Abstract
Objective:
To develop a methodology to separate intestinal stem cell (ISC)-enriched crypts from differentiated epithelial cell (DEC)-containing villi to improve the morphology of tissue-engineered intestine (TEI).
Methods:
Small intestinal tissues from 5- to 7-day-old transgenic Lgr5-EGFP mice (with fluorescently labeled ISCs) were used to measure the height of villi and the depth of crypts. Based on the significant size difference between crypts and villi, a novel cell filtration system was developed. Filtration of mixed organoid units from full-thickness intestine of transgenic Lgr5-EGFP mice allowed determination of the percentage of ISCs in the different size-based filtration fractions obtained. In vivo, 5–7-day-old Lewis rat pups were used as cell donors to obtain purified crypts and villi, and the dams of the pups served as recipients. Flat and tubular polyglycolic acid (PGA) scaffolds were seeded with either ISC-enriched crypts or DEC-containing villi and implanted intra-abdominally on the anterior abdominal wall. After 1, 3, 7, 14, 21, and 28 days of in vivo incubation, explants were processed for histologic evaluation.
Results:
Small intestine from transgenic Lgr5-EGFP mice contained villi with an average height of 134.89±41.91 μm and crypts with an average depth of 49.59±8.95 μm. After filtration, we found that the 100–200 μm fractions contained relatively pure villi in which DECs were located, whereas the 25–70 μm range fractions contained concentrated crypts in which ISCs were located. In vivo, flat PGA scaffolds implanted with purified crypts formed well-developed mucosa by day 14 postimplantation, whereas flat scaffolds seeded with villi were replaced with fibrous tissue. Tubular scaffolds seeded with the crypt fraction developed a well-formed mucosal layer on the interior surface, with 80.9% circumferential mucosal engraftment and an average villous height of 478±65 μm, which was very close to native intestine (512±98 μm), whereas tubular scaffolds seeded with the villous fraction only had 21.7% circumferential mucosal engraftment and an average villous height of 243±78 μm.
Conclusion:
The novel filtration system described can effectively and efficiently isolate ISC-containing crypts. TEI produced from ISC-containing crypts has an improved morphology that is similar to native intestine.
Introduction
T
Identification of ISCs and their markers is an active area of research. In 1998, a G-protein-coupled receptor (GPCR) with 16 leucine-rich repeats was discovered. 6 Additional leucine-rich G-protein receptors were identified and, ultimately, Hans Clevers of the Hubrecht Institute demonstrated that cells expressing Lgr5 (leucine-rich repeat-containing G-protein-coupled receptor 5, also known as Gpr49), located in the base of the crypts adjacent to Paneth cells, are multipotent ISCs. 7
In the current study, a novel filtration system with multisized sieves was used to separate cells from full-thickness intestine of transgenic Lgr5-EGFP mice that are knocked in with enhanced green fluorescent protein-labeled Lgr5. The percent of ISC-containing crypts was quantified in the different filtration fractions obtained, and the optimal size ranges have been determined to maximize the yield of ISC-containing crypts and minimize the contamination of DEC-containing villi. An in vivo study was conducted by seeding ISC-containing crypts or DEC-containing villi onto flat sheets or tubular-shaped polyglycolic acid (PGA) scaffolds and implanted on the interior surface of the anterior abdominal wall. We hypothesized that scaffolds seeded with ISC-containing crypts would produce TEI with increased villous height and circumferential mucosal engraftment and similar morphology to native intestine compared to scaffolds seeded with DEC-containing villi.
Materials and Methods
Villous height and crypt depth measurements
Transgenic Lgr5-EGFP mouse breeders were purchased from the Jackson Laboratory and used to breed neonatal mice as a source of small intestine. Both breeding and tissue harvesting protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the Research Institute at Nationwide Children's Hospital (protocol number AR12-00001). Five- to 7-day-old neonatal transgenic Lgr5-EGFP mice were euthanized and ∼5–7 cm of small intestine was harvested starting from the ligament of Treitz. After overnight fixation with 1% paraformaldehyde and 0.18% picric acid in phosphate-buffered saline (PBS), the small intestine was cut into 5 mm segments, hydrated in an optimal cutting temperature (OCT) medium for 2 h, and embedded in OCT in a dry ice 100% ethanol bath. Five micrometer sections were cut, mounted on slides, and coverslipped with Vectashield mounting media with DAPI (Vector Laboratories). Images were acquired with a confocal microscope and the height of villi and depth of crypts measured using a Zeiss LSM image browser (version 4.2.0.121), as described by Sukhotnik et al. 8 Ten high-power (20×) fields of each of 10 sections per animal from a total of 12 animals were measured, and the average villous height and crypt depth calculated.
Novel filtration system design
Figure 1 illustrates the components (Fig. 1A) and the assembly (Fig. 1B–D) of the novel filtration system designed. The bottom filter was fashioned from a filter from Tisch Scientific with the filter membrane removed. The top cups were from similar bottom filters, which were cut at the neck and the bottoms discarded. The top cups were used to secure the sieves in place and to prevent leaking at the contact surfaces. The sieves (pore sizes: 8, 15, 25, 50, 75, 100, and 200 μm) were purchased from BioDesign, Inc. and were cut to 7 cm in diameter. The cell scraper with silicone blade was purchased from USA Scientific and used to harvest cells from the sieve membranes after filtration. The bottom cup, top cups, and cell scraper were sterilized with a SterRad sterilizer using hydrogen peroxide gas plasma at a temperature of 56°C. The precut sieves were autoclaved before use. To assemble the filtration system, the smallest pore size membrane (8 μm) was placed on top of the cup of the bottom filter, and a top cup was placed on top of the sieve (Fig. 1B). By pushing down the top cup, the sieve is secured and sealed on the edge (Fig. 1C). Similarly, the other sizes of sieves are installed in the order of 15, 25, 50, 70, 100, and 200 μm. After all sieve membranes are installed, a mixed cell suspension, including villi, crypts, and individual cells, is poured into the very top cup and filtration is triggered when the vacuum is turned on. After filtration, the top cups are removed one by one, and each sieve transferred to a 25×100-mm Petri dish with sterile forceps. The cell scraper is used to remove cells from the individual sieves. Filtration with multiple sized sieves was conducted for selecting an optimal size range to maximize the yield of the cells of interest and to eliminate contamination with unwanted cells. Once the optimal size range was determined, subsequent filtration was done with the needed sieves only.

Components and assembly of the filtration system.
Concentration of ISC-containing crypts by filtration
Five- to 7-day old transgenic Lgr5-EGFP neonatal mice were euthanized and under sterile conditions the intestines were harvested starting from the ligament of Treitz to the ileocecal junction. The mesentery and smooth muscle layers were peeled off with forceps, the lumen opened lengthwise, and the intestines pooled and washed vigorously with Hank's buffered salt solution (HBSS) containing 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B to completely remove the intestinal contents. The digestion of intestines was conducted as described by Sala et al.
4
with modifications. Briefly, the small intestines were minced into ∼5–7 mm pieces and digested in 300 U/mL of collagenase type I and 0.1 g/mL of Dispase (both from Worthington Biochemical Corporation) in serum-free DMEM for 30 min at 37°C under gentle shaking. The partial enzymatic digestion was stopped by the addition of 10% fetal bovine serum in DMEM, and the suspension, which contains a mixed population of intact crypts and villi, some fragments of partially digested crypts and villi, and individual cells, was washed with DMEM until the supernatant was clear. The cells were resuspended in DMEM and poured into the very top cup of the multisized sieve filtration system. After filtration, the cells in the different size ranges were harvested, centrifuged, plated onto glass slides, and observed by a fluorescent microscope with a FITC filter. The percent of EGFP-positive ISC-containing cell clusters (crypts) in the total cell population was calculated using ImageJ software (version 1.46r). Briefly, EGFP-positive cell clusters were counted from images taken with the FITC filter. The total number of cell clusters was counted from images taken from the same area under the same magnification without the FITC filter in place. The percent of EGFP-positive cell clusters (crypts) was calculated using the following equation:
Comparison of ISC-containing crypts and DEC-containing villi in the production of TEI
Adult Lewis rat breeders were purchased from Harlan Laboratories and breeding conducted internally. Five- to 7-day-old Lewis rat pups were used as cell donors with the dam (250–300 g and 3–4 months old) of the pups used as recipients. The yield of cells from each litter of rat pups can seed multiple scaffolds and the dam of the pups can accommodate multiple scaffolds for side-by-side comparisons. All animal breeding and procedures were approved by the IACUC of the Research Institute at Nationwide Children's Hospital (protocol number AR12-00001). Small intestines harvested from the same litter (10–15 pups) were pooled and digested together as described above. The mixed cells were first filtered with a 200-μm sieve and undigested tissues on the sieve were discarded. The filtrate was further filtered with a 100-μm sieve and the cells on the sieve were harvested, labeled as villi, and kept on ice. The filtrate was then filtered with a 70-μm sieve and the cells on the sieve discarded. The filtrate that passed through the 70-μm sieve was filtered with a 25-μm sieve and cells on the sieve were harvested, labeled as crypts, and kept on ice.
PGA Biofelt (2 mm thickness and 60 mg/cm3 density) was purchased from Biomedical Structures and used to make scaffolds for the in vivo studies. Square flat scaffolds (1×1 cm) were cut directly from the original sheet. Tubular scaffolds were prepared by wrapping 1.0×2.5 cm of PGA flat sheets around 4.7-mm-diameter stainless steel mandrels to produce 1.0 cm length and 4.7 mm diameter tubular scaffolds. Both flat and tubular scaffolds were coated with 5% polylactic acid (PLLA; Sigma) in chloroform (Fisher Scientific). Once the solvent was completely evaporated, the scaffolds were soaked in 100% ethanol for 30 min and then washed thrice with PBS. Scaffolds then underwent an additional 30-min coating with 0.4 mg/mL collagen type I (Advanced BioMatrix) and were sterilized with ethylene oxide.
Purified villi or crypts harvested, as described above, were resuspended in DMEM containing 10% premium select fetal bovine serum (Atlanta Biologicals) and seeded onto one side of the flat sheet scaffolds. Two cell-seeded surfaces of flat scaffolds were sandwiched together to keep the seeded cells trapped between the two scaffold sheets. In addition, purified villi and crypts were seeded onto the luminal surface of tubular scaffolds.
Under general anesthesia with 2% isoflurane in oxygen, a laparotomy with midline incision was performed and one tubular scaffold and one pair of flat scaffolds seeded with villi were attached to the anterior surface of left abdominal wall of the recipient with 7/0 polypropylene suture. Similarly, one tubular scaffold and one pair of flat scaffolds seeded with crypts were attached to the anterior surface of right abdominal wall of the recipient and sutured in place. Incisions were closed in layers with 6/0 polydioxanone (PDO) monofilament suture. The experiment was repeated on 72 rats, which were randomly divided into 6 groups of 12 each, for the six predetermined time points. At 1, 3, 7, 14, 21, and 28 days postimplantation, 12 rats from each time point were sacrificed and 12 explants from each of the two cell types (villi and crypts) and from each of the two types of scaffolds (flat and tubular) were harvested, fixed in 10% neutral buffered formalin, embedded in paraffin, cut in 5 μm sections, and stained with periodic acid Schiff (PAS).
Immunohistochemical staining was conducted on crypt-seeded tubular scaffolds harvested 28 days postimplantation to characterize TEI and native small intestine, using primary antibodies to the following.4,9 Chromogranin A (CHRG) to identify enteroendocrine cells (Bioss, Inc.), Desmin (Bioss, Inc.) and alpha smooth muscle actin (α-SMA; Abcam) to identify intestinal subepithelial myofibroblasts (ISEMFs) and smooth muscle cells, lysozyme (LYSO; Abcam) to identify Paneth cells, Ki67 (Millipore) to identify proliferating cells, and villin (Santa Cruz) to identify absorptive enterocytes. All secondary antibodies (goat anti-rabbit, goat anti-mouse, and donkey anti-goat) were purchased from Invitrogen. Cell viability was assessed by identification of nuclei in implanted villi and crypts. 4 Villous height was measured on crypt- or villi-seeded tubular scaffolds after 28 days of in vivo incubation as described previously. The percent of circumferential mucosal engraftment of neomucosa was quantified as neomucosal surface in the percent of total available surface area in the tubular scaffold. 10 This was calculated using an average of measurements from 24 histologic sections from 12 samples and 2 histologic sections per sample at intervals of 500 μm. Statistical analyses were performed with one-way analysis of variance using GraphPad Prism 6 software (version 6.03).
Results
Size difference between ISC-containing crypts and DEC-containing villi
Figure 2 shows the morphology of small intestine from transgenic Lgr5-EGFP neonatal mice, in which DAPI staining clearly outlines the structure of villi and crypts, with Lgr5-positive ISCs located in the crypts. The average crypt depth was 49.59±8.95 μm, and the average villous height was 134.89±41.91 μm (p<0.0001). The significant difference between the size of villi and crypts was utilized to separate them using size filtration with various sized sieves.
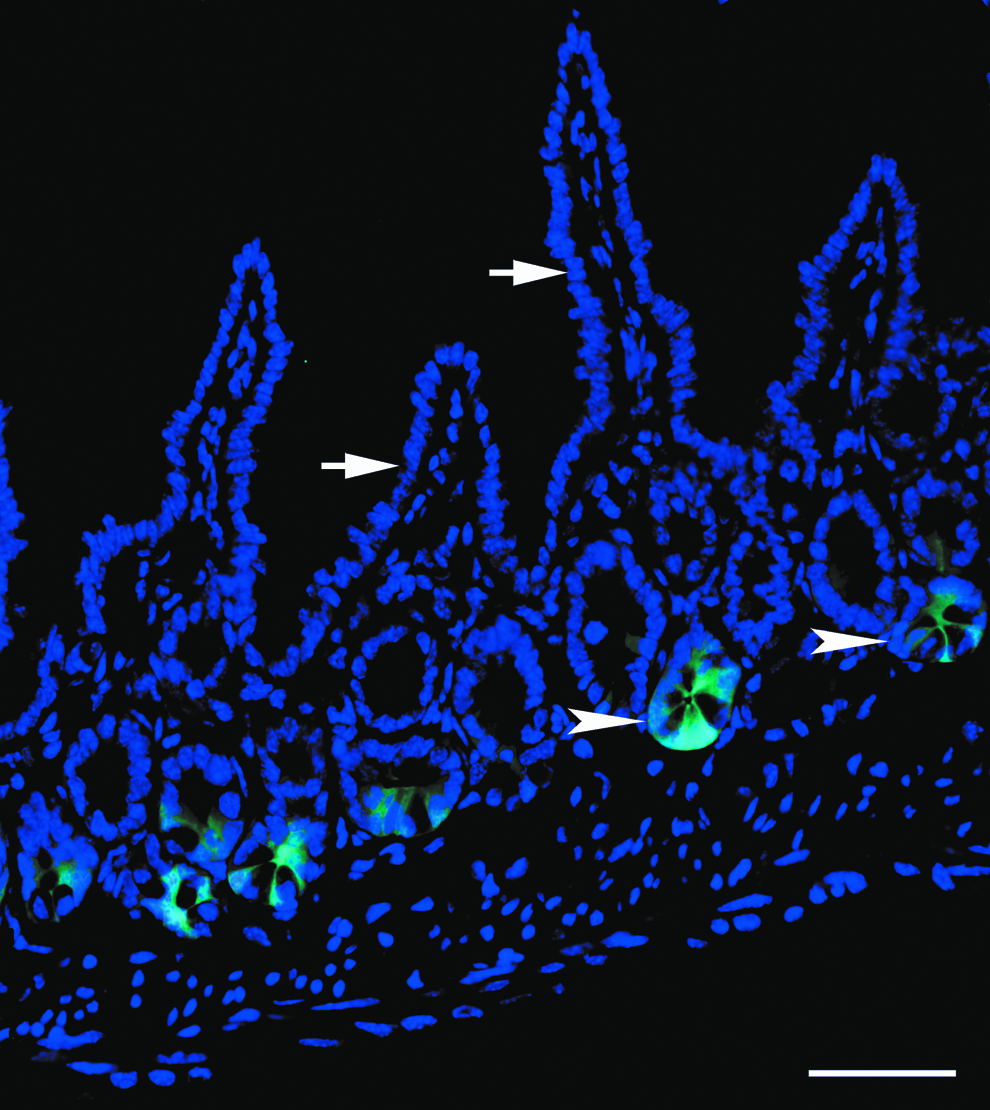
Intestinal tissue section from transgenic Lgr5-EGFP mouse. DAPI staining (blue) clearly outlines the structure of villi (white arrows) and crypts (white arrowheads) in a section of mouse jejunum. Intestinal stem cells (ISCs) are demonstrated as Lgr5-positive cells (green) and are located in the crypts. Scale bar=50 μm. ISCs, intestinal stem cells.
Optimization of sieve size to concentrate ISC-containing crypts
To optimize the sieve sizes required to obtain concentrated ISC-containing crypts and to eliminate DEC-containing villi, mixed cells from transgenic Lgr5-EGFP mouse intestine were filtered with multiple sized sieves, and the percent of ISCs in the different size range fractions was quantified. Cell clusters >200 μm containing undigested tissue debris were discarded. Villi were located in the 100–200 μm cell population, which contained no EGFP-labeled ISCs (Fig. 3A). A small fraction of ISCs (8.3%±4.5%) were identified in the 70–100 μm cell population (Fig. 3B). Relatively pure ISC-containing crypts (90.8%±5.6%) were located in the 25–70 μm cell population (Fig. 3C). Some individual ISCs or smaller cell clusters were located in the 8–25 μm cell population (Fig. 3D). The percent of the total cell population that consisted of ISC-containing crypts is listed in Table 1. The fraction of cells in the range of 25–70 μm were marked as “crypts” and the fraction of cells in the range of 100–200 μm were marked as “villi,” and both cell populations were used for in vivo studies comparing the quality of TEI produced from each.
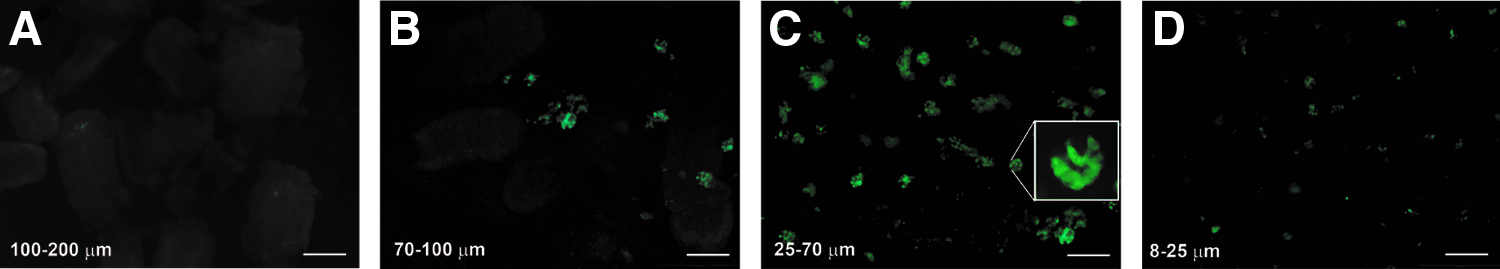
Size differentiated fractions from transgenic Lgr5-EGFP mice with EGFP-labeled ISCs.
ISC, intestinal stem cell.
In vivo cell viability on flat scaffolds
To track cell viability after implantation, crypt- or villi-enriched OUs were seeded onto one surface of flat sheets of PGA scaffolds, which were then sandwiched together with the cells located on the inside during in vivo incubation. After harvesting, the two sheets were separated and embedded face down, so the cells of interest were derived from implanted donor cells rather than migrated host cells. Implanted crypts were alive after 1 day of in vivo incubation (Fig. 4A), whereas most of the implanted villi died as confirmed by the disappearance of nuclei (Fig. 4E). On day 3, implanted crypts were still alive and formed a sprout of mucosal architecture (Fig. 4B) as indicated by the presence of PAS-positive goblet cells (dark red), whereas the implanted villi revealed merged cell membranes with only the outline of individual villi remaining (Fig. 4F). On day 7, implanted crypts formed flat mucosa (Fig. 4C), whereas implanted villi demonstrated merged villi with large voids and cellular debris at the implantation site (Fig. 4G). On day 14, implanted crypts produced neomucosa with villi and crypts (Fig. 4D), whereas implanted villi disappeared and were replaced with fibrous tissue at the implantation site (Fig. 4H). On postimplantation days 21 and 28, crypt-seeded scaffolds demonstrated multiple neomucosal cysts in which the intestinal morphology was very similar to the appearance of crypt-seeded tubular scaffolds on day 28 postimplantation (as described in the next section), while villi-seeded scaffolds retained fibrous tissue as demonstrated at 14 days postimplantation (data not shown).
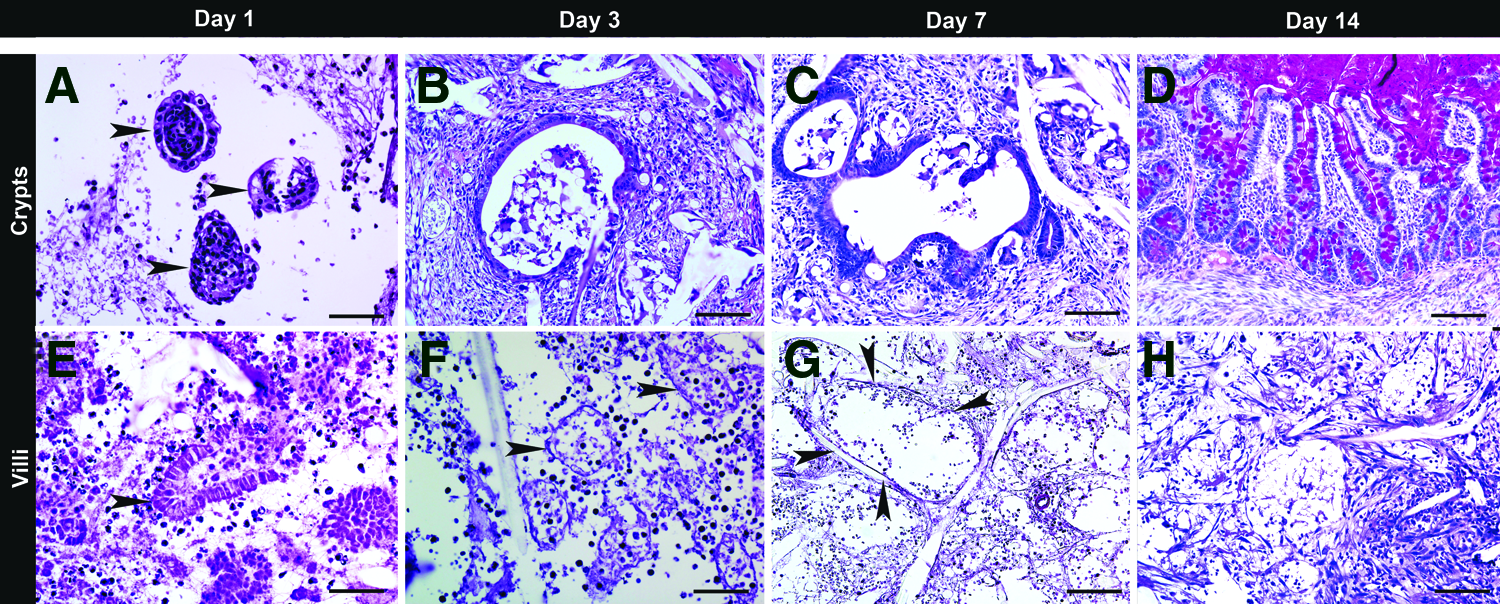
Cell viability and time course development of TEI. Flat sandwiched scaffolds were seeded with either the fraction of ISC-containing crypts (25–70 μm)
In vivo cell viability and production of TEI
The morphology of crypt- or villi-seeded tubular scaffolds at early time points (days 1, 3, 7, 14, and 21 postimplantation) was identical to that of flat scaffolds as detailed in the previous section, and therefore, the data shown here are from crypt- or villi-seeded tubular scaffolds at 28 days postimplantation. After 28 days of in vivo incubation, crypt-seeded tubular PGA scaffolds formed a mucosal layer on the interior luminal surface with a similar morphological appearance to native small intestine with long villi and dense crypts, whereas TEI produced from villi had mucosa with short disorganized villi (Fig. 5). Importantly, TEI produced from crypts contained all elements of native small intestine, including enteroendocrine cells as demonstrated by Chromogranin A staining, Paneth cells as demonstrated by lysozyme staining, absorptive enterocytes as demonstrated by villin staining, smooth muscle cells as demonstrated by Desmin/α-SMA double staining, ISEMFs as demonstrated by positive α-SMA and negative Desmin staining, and actively proliferating crypt cells as demonstrated by Ki67 staining (Fig. 6). The villous height of TEI produced from crypt-seeded scaffolds was comparable to that of native small intestine, whereas the villous height of TEI produced from villi-seeded scaffolds was significantly less (Fig. 7). The percent of circumferential mucosal engraftment of TEI produced from crypts was 80.9%, whereas that of TEI produced from villi was only 21.7% (p<0.0001) (Fig. 8).
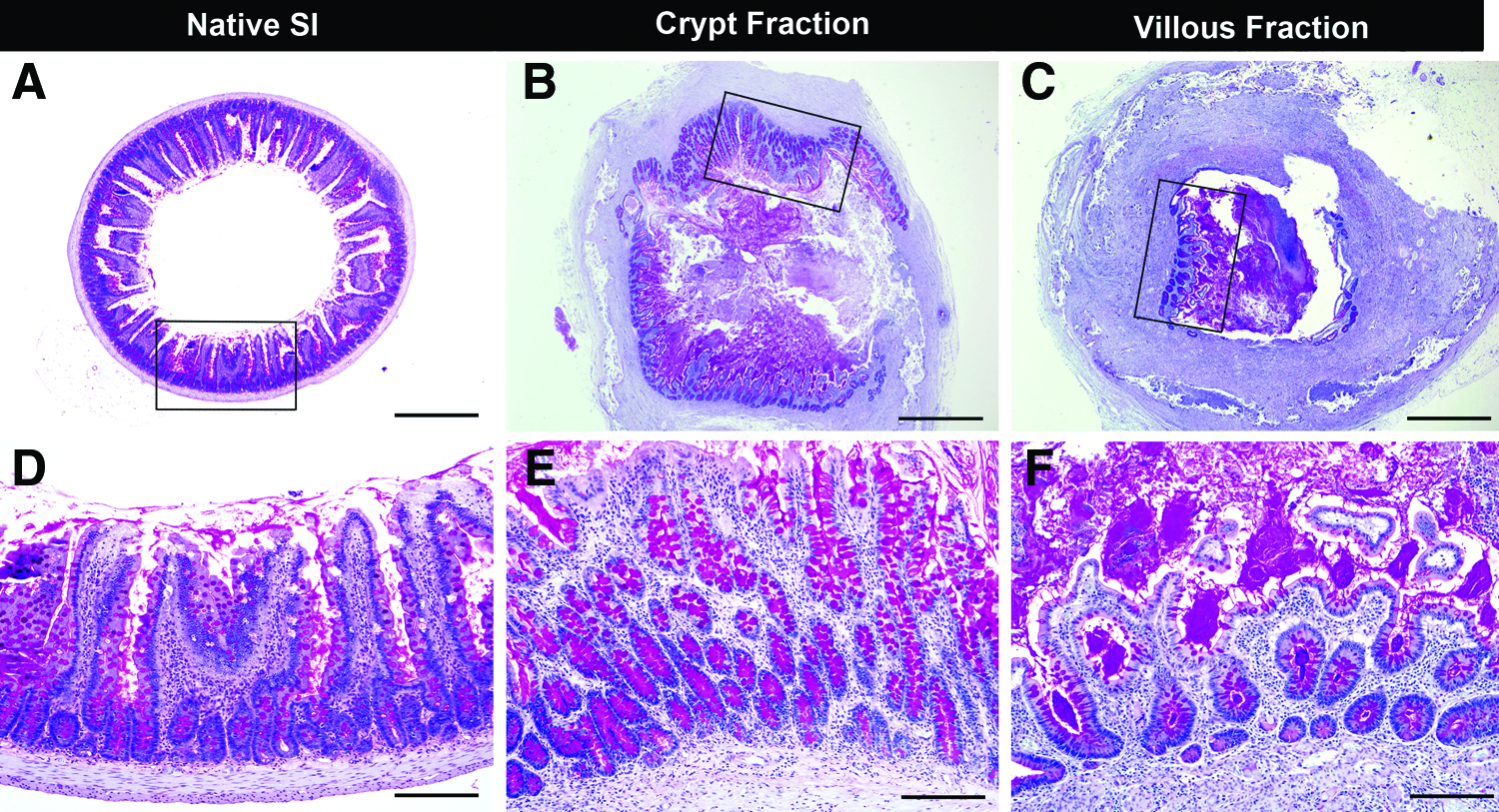
Comparison of TEI produced from ISC-containing crypts and DEC-containing villi. Tubular scaffolds were seeded with either ISC-containing crypts (25–70 μm fraction)

Characterization of TEI compared to native small intestine. Crypt-seeded scaffolds harvested 28 days postimplantation, as well as native rat small intestine, were subjected to immunohistochemical staining.
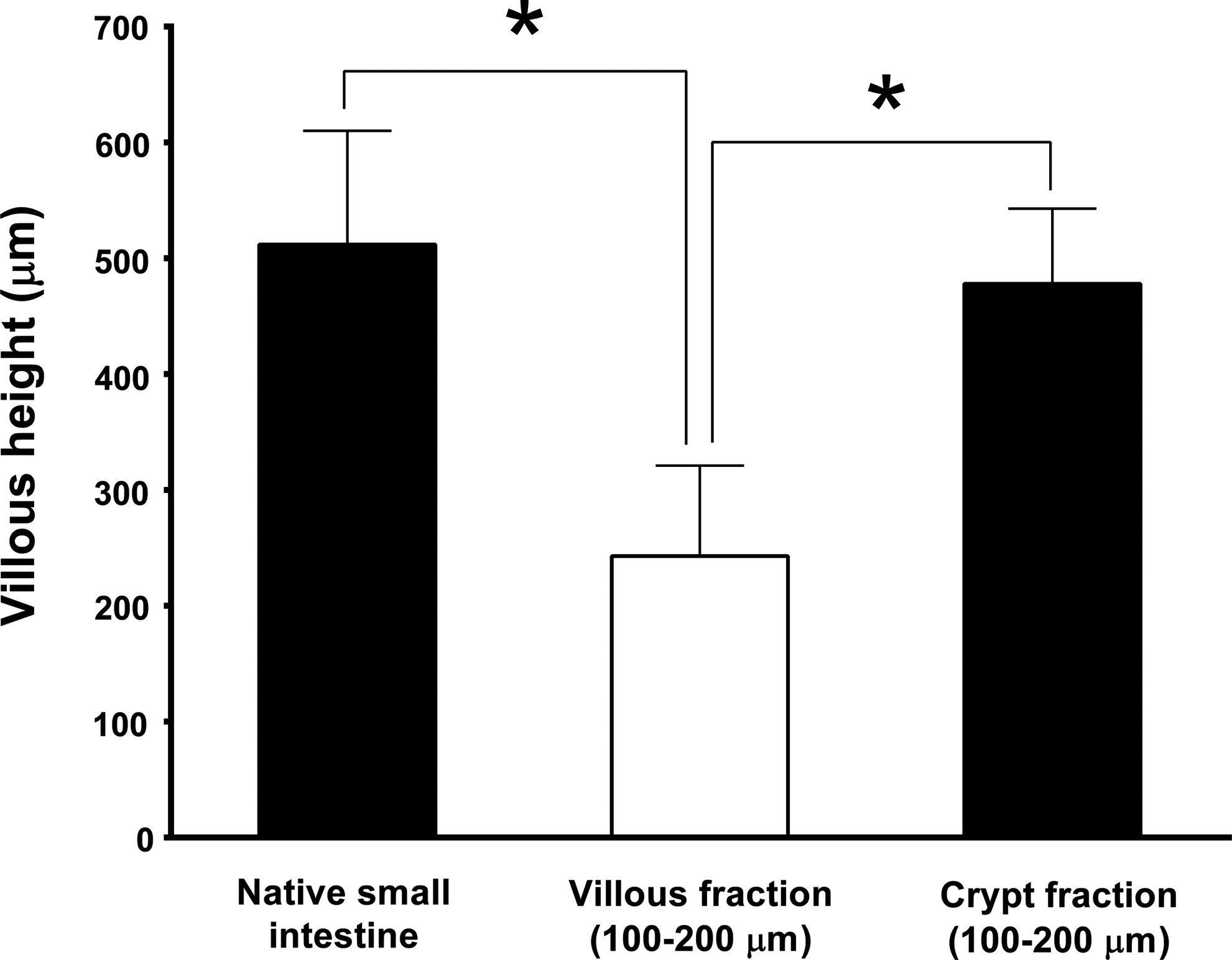
Quantification of villous height in tubular scaffolds seeded with the fraction of ISC-containing crypts (25–70 μm) or DEC-containing villi (100–200 μm) compared to native small intestine. Seeded scaffolds were incubated in the peritoneal cavity and harvested 28 days postimplantation. Asterisk indicates p<0.001.

Percent of circumferential mucosal engraftment in tubular scaffolds seeded with the fraction of ISC-containing crypts (25–70 μm) or DEC-containing villi (100–200 μm). Seeded scaffolds were incubated in the peritoneal cavity and harvested 28 days postimplantation. *Asterisk indicates p<0.001.
Discussion
Intestinal epithelium is composed of crypts and villi adjacent to a surrounding innervated muscularis that is responsible for peristalsis. Under normal conditions, the intestinal mucosa is very dynamic and regenerates its entire epithelial lining every 3–7 days. This is achieved by the proliferation of intestinal epithelial stem/progenitor cells located at the base of the crypts. As the daughter cells migrate along the crypt/villus axis, they differentiate into four specialized cell types: mucous-producing goblet cells, antibacterial protein-producing Paneth cells, hormone-producing enteroendocrine cells, and absorptive enterocytes. When they reach the top of the villus, they undergo apoptosis and are shed into the lumen of the intestine. 11 Several putative intestinal epithelial stem cell markers have been proposed, including Bmi1, 12 Lgr5, 7 CD133, 13 DcamKL-1, 14 and Musashi-1. 15 Lgr5 and CD133 label four to eight crypt base columnar cells located between the Paneth cells at the base of the crypts. In vitro studies with isolated Lgr5-positive cells have resulted in the generation of intestinal crypt/villus organoids with an epithelium containing all four differentiated intestinal epithelial cell types. 16 On the other hand, Bmi1, DcamKL-1, and Musashi-1 label cells are located in position+4, right above the Paneth cells. Subcutaneous implantation of DcamKL-1-positive cells forms spheroid-like structures with glandular epithelium. 17
Since DECs in villi no longer have the capacity to proliferate and are destined to undergo apoptosis, and since multipotent ISCs contribute to the intestinal growth, we felt that separating crypts from villi and seeding only the ISC-containing crypts would be key to the successful production of TEI. In addition, we felt that it was important to utilize intact crypts containing ISCs and their neighboring Paneth cells to maintain the mesenchymal niche as a contributor to the growth of TEI. 18
It is currently possible to separate and enrich ISCs using ISC marker (antibody)-based techniques, for example, cell sorting using flow cytometry or magnetic bead separation. There are several technical limitations to the use of those techniques for the production of TEI. First, these techniques may not efficiently yield the high numbers of cells required to meet the needs for TEI production. 19 Second, the cost of antibodies is significant. Third, a longer in vitro process may increase the chance of contamination and negatively impact cell viability. 19 Fourth, these techniques work better with individual cells rather than clusters of cells as in the ISC-containing crypts we sought to purify. 20
In our study, intestine from transgenic Lgr5-EGFP mice was used to measure the size of crypts and villi. The significant size difference between crypts and villi leads to the possibility of separation of crypts from villi by means of size filtration. Currently, commercially available cell strainers have limitations in this regard. First, the selection of pore sizes is not very broad, with only 40, 70, and 100 μm pore sizes available. Second, the bottom working area is 6.2 cm2, which is too small to filter the 3–5-mL cell pellet suspended in 30–50 mL of cell culture medium required to seed multiple scaffolds. Third, the recovery of cells from the strainer is not convenient or efficient. A commonly used method reported by others to separate crypts from villi is by sedimentation.21–25 Although we have tried this method in our laboratory, there is no clear interface between the layers of crypts and villi after the mixed cell suspension settles down, thereby contaminating the desired crypt cell population with villi.
In the current study, a novel cell filtration system was developed to overcome these disadvantages. With a larger working surface area (12.4 cm2) and a bigger volume of the top cup, the filtration system can easily filter 30–50 mL of cell suspension. Moreover, the application of a gentle vacuum reduces the chance of cell clogging in the sieves. The removable sieves can be transferred to a cell Petri dish for convenient and efficient cell recovery. Additionally, the removable and replaceable sieve membranes provide a broad range of pores sizes from 8 to 200 μm. Note that since the villous height and crypt depth of mouse pups is very close to that of rat pups, we showed that the same protocol can be used to separate mouse pup crypts from villi (data not shown). Although optimized for mouse and rat pup intestine, this protocol can be adapted for adult mouse and rats, as well as for human intestine, using the same strategy.
Crypt-seeded scaffolds formed a neomucosa that almost covered the entire circumferential surface of the scaffolds by 28 days postimplantation. The morphology of the neointestine was very similar to native intestine with a fully differentiated epithelium as confirmed by specific epithelial markers, containing absorptive enterocytes, mucus-producing goblet cells, hormone-producing enteroendocrine cells for control of the digestion process, 26 and Paneth cells, which produced antibacterial proteins and contribute to the regulation of the intestinal flora. 27 Moreover, it contains ISEMFs, which are key mesenchymal cells supporting ISCs, and a major source of instructive signals to the intestinal epithelium.28–30 In contrast, scaffolds seeded with DEC-containing villi produced TEI with mucosa that only covered a small portion of the circumference of the scaffolds, with mucosa containing shorter disorganized villi. Mucosal engraftment in crypt-seeded scaffolds was significantly higher than that in villi-seeded scaffolds. The in vivo data confirmed that the novel filtration system we have described can efficiently separate crypts from villi and can provide concentrated ISC-containing crypts, which contribute to the improved quality of TEI.
In summary, these results support the use of purified crypts in the production of TEI. In future we will perform side-by-side comparisons of the quality of TEI produced with our filtration method to that of published sedimentation protocols. In addition, we will focus on the functionality of TEI after implantation in continuity with native intestine in an effort to treat short bowel syndrome. Furthermore, TEI may provide a useful tool for studying stem cell biology, cell proliferation, migration and differentiation, and general intestinal epithelial cell physiology in the future.
Footnotes
Acknowledgments
This study was supported by grants from the Research Institute at Nationwide Children's Hospital and from Ohio Third Frontier. The authors thank Cynthia Mcallister, Patricia Craig, Makenzie Eisnaugle, Spencer Dunaway, and Melanie Herring for their assistance in sample preparation for histological assessment. The authors also thank Palak Painter for her breeding of LGR5-EGFP mouse pups.
Disclosure Statement
No competing financial interests exist.