
Abstract
Functional reconstruction of lost tissue by regenerative therapy of salivary glands would be of immense benefit following radiotherapy or in the treatment of Sjogren's syndrome. The purpose of this study was to develop primary cultures of human salivary gland cells as potential regenerative resources and to characterize their acinar/ductal phenotype using electrophysiological measurements of ion transport. Human salivary gland cultures were prepared either from adherent submandibular gland cells (huSMG) or from mixed adherent and nonadherent cells (PTHSG) and were cultivated in Hepato-STIM or minimum essential medium (MEM). Expression of key epithelial marker proteins was determined by quantitative reverse transcription polymerase chain reaction (RT-PCR). Transepithelial electrical resistance (TER) was monitored following seeding the cells on Transwell membranes. Transepithelial ion transport was estimated by short-circuit current (Isc) measurements in an Ussing chamber. Both huSMG and PTHSG cells showed epithelial characteristics when cultivated in Hepato-STIM, while fibroblast-like elements dominated in MEM. Compared to intact tissue, cultivation of the cells resulted in substantial decreases in AQP5 and NKCC1 expression and moderate increases in claudin-1 and ENaC expression. Both cultures achieved high TER and transepithelial electrolyte movement in Hepato-STIM, but not in MEM. The Isc was substantially reduced by basolateral Cl− and bicarbonate withdrawal, indicating the involvement of basolateral-to-apical anion transport, and by the blockade of apical ENaC by amiloride, indicating the involvement of apical-to-basolateral Na+ transport. An almost complete inhibition was observed following simultaneous ENaC block and withdrawal of the two anions. Isc was enhanced by either apical adenosine triphosphate (ATP) or basolateral carbachol application, but not by forskolin, confirming the expected role of Ca2+-activated regulatory pathways in electrolyte secretion. Inhibition of basolateral NKCC1 by bumetanide reduced the response to ATP, indicating the active involvement of this transporter in Cl− secretion. In conclusion, we have demonstrated that both PTHSG and huSMG primary cultures cultivated in Hepato-STIM form two-dimensional monolayers in vitro on permeable supports and achieve active vectorial transepithelial electrolyte transport. The presence of both basolateral-to-apical anion fluxes and an apical-to-basolateral Na+ flux indicates both acinar and ductal characteristics. With further refinement, this model should provide a firm basis for new interventions to correct salivary gland dysfunction.
Introduction
T
Certain drugs and systemic diseases can also cause dry mouth; however, the two major causes are Sjogren's syndrome, an autoimmune disease, and radiation-induced salivary hypofunction following radiotherapy of oral and pharyngeal cancer patients. 3 In these cases, it is mainly acinar cell function that is lost, while the ductal system remains intact.1,3,6,7 In human salivary glands, the acinar cells are normally the main source of fluid, electrolyte, and protein secretion, while the ducts have primarily an absorptive role.4,5,8 Functional acinar cells are therefore essential for maintaining healthy oral conditions. When acinar cells are damaged or lost, active electrolyte transport and the resulting flow of fluid into the mouth are also lost.1–3,6,7 To restore fluid flow, the vectorial electrolyte transport mechanisms have to be reconstructed.2,3
Currently, there are no effective pharmacological tools for the treatment of salivary hypofunction, but various promising therapeutic avenues are emerging. 9 These include salivary gland regeneration from stem cells,3,10 development of tissue-engineered artificial salivary glands (secretory devices surgically implanted under the oral mucosa), development of new pharmacological treatments, and gene transfer-based treatments.11–13 The key requirement for these strategies is to create functional salivary secretory units, which are amenable to pharmacological and/or gene transfer manipulations. These will ultimately serve as grafts in an artifical salivary gland, providing both vectorial electrolyte transport from blood to saliva and the consequent secretory flow of water.
The HSG human submandibular adenocarcinoma cell line was the first to be studied for this purpose; however, these cells were unable to form tight junctions and were incapable of controlled fluid transport. 14 Recently, several methods have been developed to obtain human primary salivary gland cell cultures such as using nonattaching epithelial cells (huSMG - human submandibular gland; and huSG - human submandibular or parotid gland)1,15,16 and a mixture of nonattaching and surface-attaching cells (PTHSG, primary total human salivary gland). 17 The cells in these cultures are able to form tight junctions and build up a functional epithelial barrier, but all have the same limitation: most of the cells in the culture are of ductal phenotype.16–18
The use of Matrigel to promote acinar cell differentiation can help to create in vitro 3D salivary gland models. However, Matrigel is derived from a mouse sarcoma and for safety reasons it cannot be used clinically as part of an implantable device.18–20 Consequently, recent studies have attempted to apply other scaffolding materials to promote acinar cell differentiation such as the human extracellular matrix (ECM) peptide fragment, perlecan domain IV (PlnDIV), 21 fibrin hydrogel,22,23 and polyacrilamide gels. 24 These studies have met with varying success, but none of them has investigated the electrolyte transport capabilities of the salivary cells. There have also been successful attempts to cultivate primary cultures on permeable supports, but measurements were restricted to measuring transpepithelial resistance without any functional assessment of transepithelial electrolyte transport.16,18,20
The aim of the present study was to compare two different primary cultures of human salivary gland cells (huSMG and PTHSG) when cultivated in Hepato-STIM and minimum essential medium (MEM) and to characterize their acinar and ductal characteristics using electrophysiological measurements of ion transport.
Materials and Methods
Primary cell cultures
The method for preparation of cell cultures from the human submandibular gland is based on previously developed protocols of Szlavik et al. 17 for PTHSG and Tran et al. 200516,17 for huSMG, with several minor modifications. Portions of submandibular salivary gland were obtained from the Department of Oro-Maxillofacial Surgery and Stomatology, Semmelweis University (Budapest, Hungary). The samples were collected from patients undergoing neck dissection surgery with the permission of the Regional Human Research Ethics Comittee (permission number: 67/2005). Patients involved received no prior irradiation treatment or chemotherapy and none of the patients had salivary gland tumors or other pathological salivary gland conditions. Forty-nine salivary gland samples were obtained from 49 patients with ages ranging from 40 to 86 years. The histology of all gland samples was typical for normal human submandibular glands.
Dissected salivary gland samples (wet weight 1–2 g) were transferred on ice in RPMI-1640 medium(Sigma-Aldrich supplemented with 5% fetal calf serum) (FCS; Gibco Invitrogen) and antibiotic/antimycotic solution (100 U/mL penicillin, 100 mg/mL streptomycin, 2.5 mg/mL amphotericin-B; Sigma-Aldrich). Before and during mechanical tissue mincing, the samples were washed several times with the DMEM-F12 (Gibco) supplemented with antibiotic/antimycotic solution. The minced tissue was transferred into 20 mL of dissociation buffer and incubated in a water bath for 1 h at 37°C with vigorous vortexing every 20 min. The dissociation buffer contained 0.04% trypsin (Gibco) and 0.15 mg/mL collagenase (Sigma-Aldrich) in the DMEM-F12. After 1 h, the supernatant, that is, the fraction with already dissociated cells, was transferred into another Falcon tube containing 20 mL DMEM-F12.
The incubation of the rest of the tissue continued with a 20 mL fresh dissociation buffer, and the supernatant fraction was transferred again after an hour. At the end of the incubation, all dissociated cell fractions were collected into one dish. The whole dissociation process took approximately 2.5 h. This sequential digestion protocol minimized enzymatic damage to the epithelia, providing more viable epithelial cells and, most importantly, a significant number of acinar cells in the cell culture (21). One milliliter of 10 mg/mL Dnase I (Roche Diagnostic Corporation) was added, and the cell suspension was triturated for 2 min.
The cell suspension was centrifuged at 230 g for 5 min, the supernatant was discarded, and 20 mL cold Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich) with 10% FCS was used to resuspend the cell pellet. The cell suspension was centrifuged again (230 g, 5 min) and the supernatant discarded. The cell pellet was suspended in 10 mL of Hepato-STIM (BD Biosciences), supplemented with 10% FCS, with antibiotic/antimycotic solution and 1% glutamine (Sigma-Aldrich). This suspension was filtered through a 70 μm cell strainer (BD Biosciences), plated onto 60 mm Primaria tissue culture dishes (BD Biosciences), precoated with FCS, and incubated overnight at 37°C in 5% CO2.
Next morning, the supernatant was transferred into a second dish. This was to separate the quickly attaching fibroblast-like cells from nonattaching epithelial cells. The huSMG cell culture consisted solely of 1-day-old floating cell aggregates 16 containing only nonattaching cells, while the PTHSG cell culture included both quickly attaching and nonattached cells. 17 We examined two different media for the growth of cell cultures and for monolayer formation: Hepato-STIM supplemented with 10% FCS, antibiotic/antimycotic solution, 1% glutamine, and the Minimal Essential Medium (MEM; Sigma-Aldrich) with the same supplementation of FCS and antibiotic/antimycotic solutions. The culture medium was changed thrice a week: approximately one-third of the old solution was left on the cells and diluted with a fresh medium. The cells were harvested when the plate reached 80% confluence using 0.25% trypsin–ethylenediaminetetraacetic acid (trypsin-EDTA; Gibco).
In previous studies 17 and also in preliminary experiments, we changed the medium completely on both plastic culture dishes and Transwell filters thrice a week. However, using conditioned medium to retain secreted growth factors, 25 the cells reached confluence much faster, and on Transwell membranes, the transepithelial electrical resistance (TER) rose above 500–600 Ω cm2 in both PTHSG and huSMG cultures. For experiments, the cells from passages 1 to 3 were seeded onto permeable polyester supports (Transwell Clear, Corning Costar) with an area of 1.12 cm2 and 0.4 μm pore size to obtain polarized confluent monolayers. The Transwell membranes were coated with FCS before cell plating (15 min incubation, 37°C). Confluence was assessed by measuring TER with an epithelial voltohmmeter (EVOM, World Precision Instruments) at daily intervals after seeding.
Expression of key marker proteins
We studied the mRNA expression of typical salivary gland transporters and chose five acinar and ductal markers (Na+/K+/2Cl− cotransporter NKCC1, epithelial Na+ channel ENaC, claudin-1, aquaporin-5, amylase) for further quantitative analysis. Total RNA was isolated from the salivary gland tissue samples as well as from cultured PTHSG and huSMG cells grown both on plastic Petri dishes and on Transwell membranes. The salivary gland samples were homogenized using a Potter homogenizer, and the RNA was isolated using TRI Reagent (Life Technologies). Total RNA from cell cultures was extracted by a spin column method, using the NucleoSpin RNA II kit (Macherey-Nagel), according to the manufacturer's protocol. The RNA concentration was measured by a NanoDrop spectrophotometer, and RNA quality was checked by gel electrophoresis. One microgram of each high-quality RNA sample was then reverse transcribed using the High-Capacity RNA-to-cDNA Master Mix (Life Technologies). For conventional reverse transcription polymerase chain reaction (RT-PCR), we used Diamond Mix (Bioline) and the specific primers listed in Table 1. In this study, we investigated the expression of ENaC, NKCC1, Na+-HCO3− cotransporter NBC1, Na+/H+ exchanger NHE1, and Cl−/HCO3− exchanger AE2, all of them are regarded as key proteins required for normal salivary gland electrolyte transport.4,5,8,26
RT-qPCR amplification was performed using the ABI StepOne System with TaqMan Universal Master Mix (Applied Biosystems) and the predesigned primers (Applied Biosystems) listed in Table 2. We used RPLP0 as a housekeeping control 17 and the ΔΔCt method to quantify gene expression.
Solutions for functional studies
The standard HEPES-buffered solution contained (in mM) 137 NaCl, 5 KCl, 1 CaCl2, 1 MgCl2, 10 D-glucose, and 10 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and was equilibrated with 100% O2. The standard HCO3−-buffered solution contained (in mM) 116 NaCl, 25 NaHCO3, 5 KCl, 1 CaCl2, 1 MgCl2, 10 D-glucose, and 5 HEPES and was equilibrated with 5% CO2/95% O2. Na+- and Cl−-free solutions contained equimolar N-methyl-D-glucamine (NMDG+) and gluconate in the place of Na+ and Cl−, respectively. In solutions containing NH4+, the concentration of Na+ was reduced to maintain the osmolarity. All solutions were adjusted to pH 7.4 at 37°C, and all reagents were purchased from Sigma-Aldrich.
Measurement of short-circuit current
Confluent monolayers of PTHSG and huSMG cells, grown for at least 4 days on FCS-coated, permeable polyester supports (Transwell, 12 mm diameter, 0.4 μm pore; Corning Costar), were mounted in a modified Ussing chamber. The apical and basolateral compartments were each perfused continuously at a flow rate of 2 mL/min. The transepithelial potential difference (Vm) was measured with agarose-KCl bridges and Ag-AgCl electrodes connected to an epithelial voltage clamp amplifier (EC-825, Warner Instrument Co). The short-circuit current (Isc) was measured at 30 s intervals by voltage clamping to 0 mV for 5 s with Ag/AgCl wire current electrodes mounted directly in the bath fluid. A positive current can represent either a negative charge transfer from the basolateral to the apical side or a positive charge transfer from the apical to the basolateral side.
Amylase release measurements
Samples of media from PTHSG and huSMG cultures were collected to measure the secreted amylase activity. Cells grown either on a plastic surface or on Transwell filters were incubated in Hepato-STIM for 72 h. Samples were taken of media bathing both the apical and basal surfaces of the cell cultures. Amylase activity was measured by Phadebas Amylase Test (Magle AB), which is based on starch hydrolysis. Values were normalized to 1 m2 of cultured monolayer and are given in U/L in 1 h.
Statistics
Averaged values are presented as the mean ± SEM. Statistical comparisons were performed using analysis of variance followed by a Tukey post hoc test or using a t test, where only two groups were to be compared. Where data were not normally distributed (e.g., HEPES data in Fig. 5E), nonparametric tests were used.
Results
Primary culture
When established, both PTHSG and huSMG cell cultures showed similar growth characteristics, although the huSMG cultures contained more floating and semiattached cell aggregates than the PTHSG preparations initially after plating. The attached cells in both cultures formed epithelium-like patches separated by thin lines of fibroblast-like cells. With increasing passage number, the number of semiattached cell aggregates diminished. In the present study, the cells were used between passages 1 and 3. Cell cultures reached appoximately 80% confluence 5–6 days after seeding and grew as monolayers. Initially, the attachment and proliferation properties of the cells were similar in MEM and in Hepato-STIM. However, the cultures in MEM contained considerably higher numbers of fibroblast-like cells from early time points and these cells grew over the epithelium-like cells (Fig. 1C, D). Micrographs showed that fibroblast-like cells became dominant and were mixed with sporadic islands of epithelial cells even before the first passage in MEM.

Morphology and transepithelial electrical resistance (TER) of PTHSG and huSMG cultures. Phase-contrast micrographs of PTHSG and huSMG cultures grown in Hepato-STIM
In Hepato-STIM, both PTHSG and huSMG cultures formed mainly epithelium-like patches suggesting that Hepato-STIM acts as a selection factor for epithelial cell promotion over mesenchymal cells. Cobblestone-like patches formed a tight monolayer with very few elongated fibroblast-like cells (Fig. 1A, B). From passage 4, even in Hepato-STIM, fibroblast-like cells became dominant and the cultures were discarded.
Monolayer and tight junction formation
The formation of functionally polarized and confluent monolayers is essential for epithelial fluid and electrolyte secretion. The effects of different culture media on epithelial barrier formation were observed by TER measurements on cells plated onto Transwell permeable supports. Although the cells were capable of growing in MEM, they did not attain high TER values: the values were typically below 200 Ω cm2. In Hepato-STIM, the cells reached confluence in 7–10 days, and TER levels increased to 595 ± 93 Ω cm2 (PTHSG, n = 55) and 622 ± 117 Ω cm2 (huSMG, n = 45).
Expression of marker proteins
To investigate the expression of marker protein characteristic of salivary epithelial cells, we performed two sets of experiments. The first served to detect quantitative changes in key salivary cell markers compared with the whole human salivary gland: amylase, the aquaporin water channel AQP5, and the cation-chloride cotransporter NKCC1, which are primarily acinar markers, and the tight junction protein, claudin-1, and the epithelial sodium channel, ENaC, which were presumed to indicate ductal phenotype. mRNA expression of these five proteins could be detected in both PTHSG and huSMG preparations (Fig. 2A).

Expression of salivary acinar and ductal cell markers and transporters.
Our data show practically no difference between the two preparations, but we found a striking three orders of magnitude decrease in the acinar marker amylase and AQP5 and a similar, but much less pronounced, decrease in NKCC1 expression in the cultured cells compared to the whole human salivary gland.In contrast, culturing of the salivary cells induced a substantial increase in claudin-1 expression and a much less pronounced increase in ENaC. Interestingly, culturing the cells on permeable supports, which enables them to polarise with distinct apical and basolateral membrane domains accessible to fluid, did not significantly modify the expression patterns of either of the two cell cultures (Fig. 2A).
Amylase expression was significantly higher in cells grown on Transwell supports than in those cultivated on a plastic surface, indicating a moderate enhancement of the acinar phenotype in the polarized cells. Furthermore, amylase was released into the medium in both culture conditions [huSMG: 742 ± 214 U/L on plastic and 5022 ± 1682 U/L on Transwells (apical surface medium); PTHSG: 568 ± 193 U/L on plastic and 3043 ± 882 U/L on Transwells (apical surface medium); (n = 9–11)]. Salivary cell polarization was clearly shown by the fact that in cells cultivated on Transwells, no amylase was detectable in the medium bathing the basal surface of the cultures, whereas significant amounts were detected in the medium bathing the apical surface.
We also applied semiquantitative RT-PCR to assess the expression of a number of other important electrolyte transporters in PTHSG cells. As expected, ENaC, NKCC1, NBC1, NHE1, and AE2 mRNAs were all identified in the native human submandibular gland tissue homogenate. We observed no substantial changes in ENaC, NHE1, and AE2 in PTHSG cultures, while decreased expression of NKCC1 and NBC1 was obvious in cells grown either on plastic or on Transwell membranes (Fig. 2B). These observations support the quantitative PCR data described above.
Basal short-circuit current
In the HCO3−-buffered solution, a significant Isc of approximately 6 μA/cm2 was detectable in the absence of stimulation in both PTHSG and huSMG cells (Fig. 3). This basal current could be partially inhibited by substitution of Cl− and HCO3− on both sides of the epithelium (Fig. 3A). The solution change evoked a brief transient increase in Isc, but in the steady state, the effect was to significantly inhibit the current (Fig. 3B).
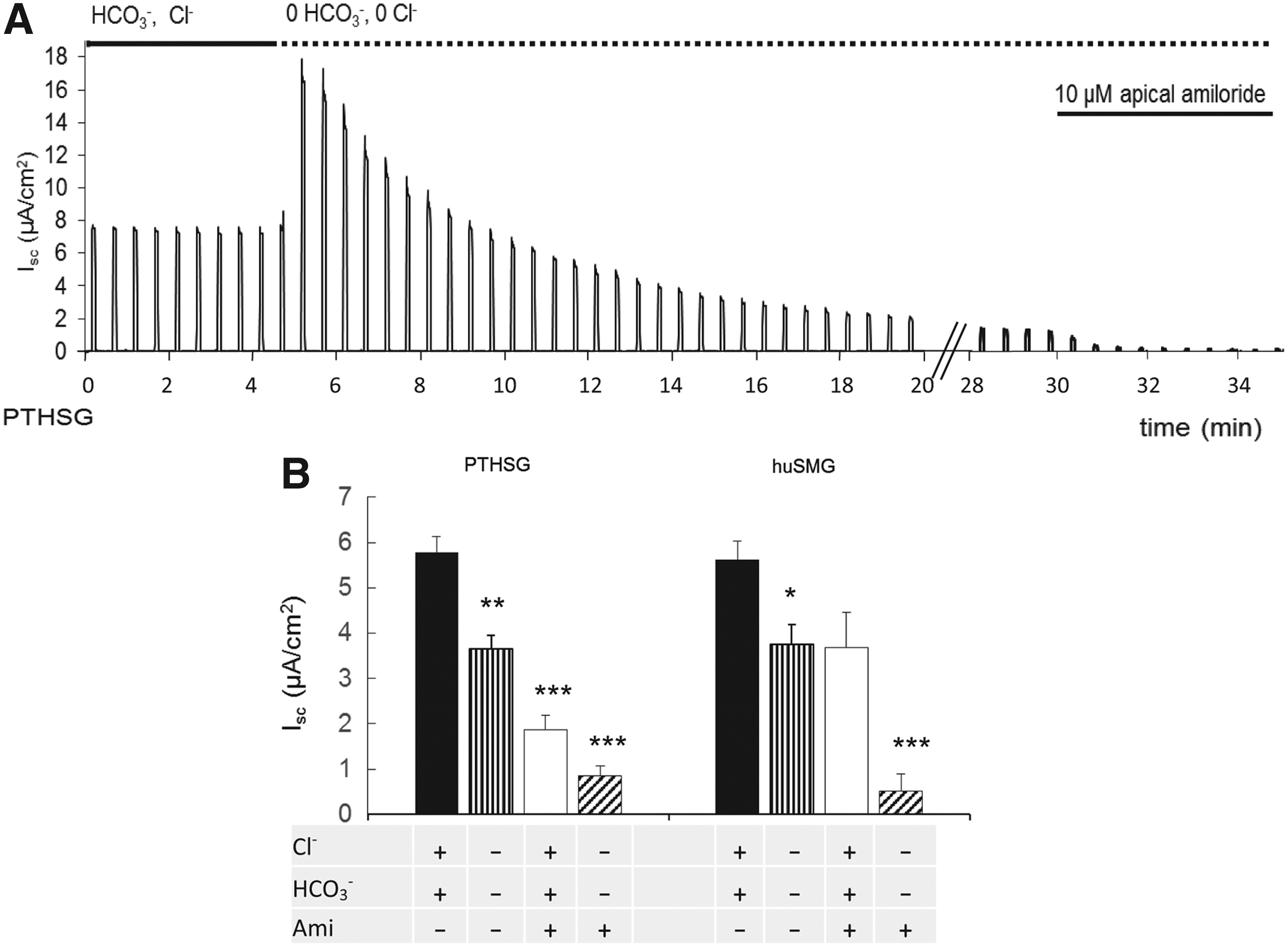
Effects of anion substitution and ENaC block on the basal short-circuit current (Isc) generated by unstimulated PTHSG and huSMG monolayers. Changes in the basal Isc of PTHSG cells following bilateral substitution of Cl− and HCO3− and subsequent application of 10 μM amiloride at the apical membrane
In the absence of Cl− and HCO3−, the remaining current was most likely due to Na+ absorption. In the salivary duct, the epithelial sodium channel, ENaC, plays a key role in Na+ absorption and it can be selectively blocked by low concentrations of amiloride. With Cl− and HCO3− present in the bathing solutions, application of 10 μM amiloride to the apical surface reduced the basal Isc by 40–60% (Fig. 3B). In the absence of Cl− and HCO3−, Isc was almost completely abolished by apical amiloride (Fig. 3A). These data suggest that the basal Isc may be attributed partly to Cl− and/or HCO3− secretion and partly to Na+ absorption, a combination of both acinar and ductal transport activities.
Response to stimulation
Stimulation of apical purinergic receptors with 50 μM adenosine triphosphate (ATP) caused Isc to increase by approximately 5 μA/cm2 in both PTHSG and huSMG monolayers (Fig. 4A). The time course of the change is shown in Figure 5A. The response was transient, rising over a period of 2 min to a peak and then falling more slowly to the basal level despite the sustained stimulation. The response to ATP was similar in the absence of HCO3− (Fig. 5B). However, the increase in Isc (ΔIsc) was significantly smaller in the absence of both Cl− and HCO3− (PTHSG: ΔIsc = 2.8 ± 0.8 μA/cm2, n = 5; huSMG: ΔIsc = 1.6 ± 0.7 μA/cm2, p < 0.05, n = 8).

Peak changes in short-circuit current (ΔIsc) following stimulation of PTHSG and huSMG monolayers with ATP, carbachol, and forskolin.
Effects of NKCC1 inhibition and ENaC block on the changes in short-circuit current (Isc) following stimulation of PTHSG monolayers with ATP in the presence and absence of HCO3−.Changes in Isc evoked by 50 μM ATP applied to the apical membrane in the presence
Blockade of the ENaC channels with 10 μM amiloride also significantly reduced the Isc changes evoked by ATP (PTHSG: ΔIsc = 3.4 ± 0.6 μA/cm2, n = 9; huSMG: ΔIsc = 2.4 ± 0.3 μA/cm2, p < 0.05, n = 5). These results suggest that both anion secretion (Cl− and/or HCO3−) and Na+ absorption increase following purinergic stimulation.
When we explored the possible relationship between the TER values (measured immediately before short-circuit experiments) and the Isc responses to ATP stimulation in individual Transwell preparations, we found only a very weak association between the two values (Fig. 4B). These data indicate that Isc activity is not strongly related to TER values determined by tight junction formation.
Basolateral application of the muscarinic agonist carbachol CCh, 100 μM, which acts through the IP3/Ca2+ signaling pathway, caused a smaller but still significant increase in Isc in both PTHSG and huSMG cells (Fig. 4A). In contrast, apical application of forskolin (10 μM), which elevates intracellular cAMP by stimulating adenylyl cyclase, had no significant effect on Isc (Fig. 4A).
The PTHSG and huSMG cells showed no significant difference in their basic functional and molecular characteristics. However, the huSMG cells were much more difficult to handle, and unexplained cell death occured more often than in the PTHSG cell cultures. Therefore, subsequent functional experiments were performed only on the PTHSG cells.
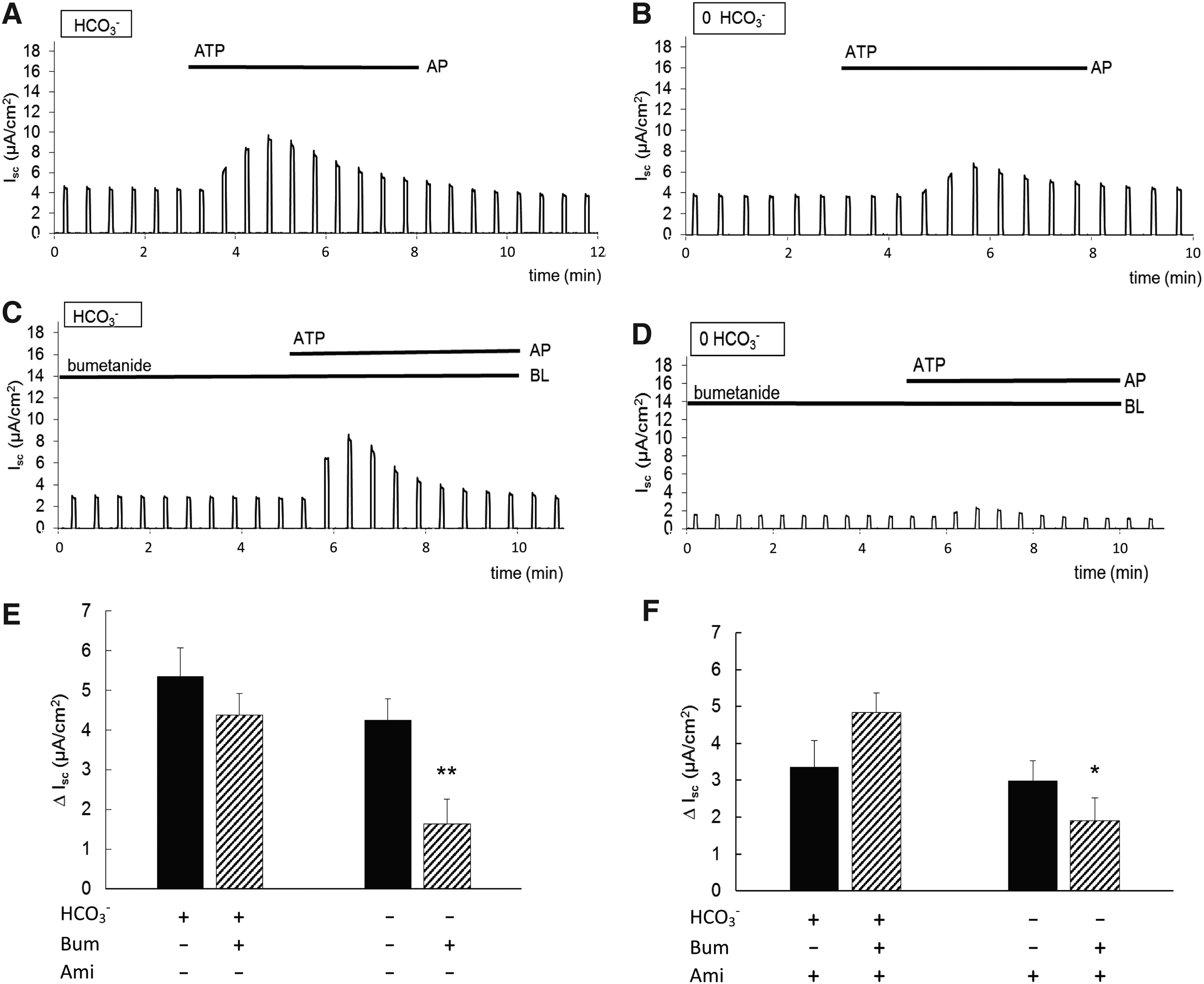
Inhibition of basolateral transporters
Uptake of Cl− across the basolateral membrane, by secondary active transport, is the main driving force for fluid and electrolyte secretion by salivary acinar cells. To evaluate the contribution of the basolateral Na+-K+-2Cl− -cotransporter (NKCC1), which is expected to be the major pathway of Cl− uptake on the basolateral side, we applied bumetanide (100 μM), a selective inhibitor of NKCC1, to assess its contribution to the Isc generated by PTHSG cells. When bumetanide was added to the HCO3−-buffered solution on the basolateral side of the monolayer, there was no significant change in the basal Isc (data not shown), nor in the increase in Isc following stimulation with apical ATP (Fig. 5C). However, in the absence of HCO3−, pretreatment with bumetanide resulted in a ∼60% decrease in the response to ATP (Fig. 5D). These results are summarized in Figure 5E. They suggest that a substantial fraction of the current generated in presence of HCO3− is mediated by a HCO3−-dependent process that does not involve NKCC1 activity. Only when this is blocked by omission of HCO3− from the solutions do we see a significant contribution from NKCC1.
This experiment was then repeated in the presence of apical amiloride (10 μM) to eliminate the contribution of ENaC-mediated Na+ absorption. When the monolayer was bathed with the HCO3−-buffered solution, amiloride had no significant effect on the ATP response in the presence or absence of bumetanide. However, in the absence of HCO3−, and with amiloride present in the apical solution, bumetanide once again caused a significant decrease in the ATP response (Fig. 5F). This indicates that the HCO3−-dependent component of ΔIsc derives from anion secretion rather than Na+ absorption. It could be due either to secretion of HCO3− itself or HCO3−-dependent secretion of Cl−, driven perhaps by a basolateral Cl−/HCO3− exchanger.
Salivary acinar cells express Cl−/HCO3− and Na+/H+ exchangers (AE2 and NHE1) and Na+/HCO3− cotransporters (NBC1) at the basolateral membrane. These mediate Cl− and HCO3− uptake and contribute to the driving force for secretion across the apical membrane. To evaluate the contribution of HCO3− secretion to Isc, the following experiments were performed in the absence of Cl−. Simultaneous application of 30 μM EIPA and 500 μM H2DIDS, to inhibit basolateral HCO3− accumulation through NHE1 and NBC1, respectively, resulted in a decrease of approximately 20% in the basal Isc (Fig. 6A). The current was reduced further, but not completely, by addition of 10 μM amiloride at the apical membrane. The origin of this remaining component of the basal current is unknown, since all of the known pathways for Na+ and HCO3− transport were blocked by the inhibitors and Cl− was absent from the solutions.

Effects of NHE1 and NBC1 inhibition and ENaC block on the basal and ATP-evoked changes in short-circuit current (Isc) recorded from PTHSG monolayers in the absence of Cl−. Basal Isc values
The basolateral transport inhibitors had a greater effect on the increase in Isc evoked by ATP stimulation. In the absence of Cl−, EIPA and H2DIDS reduced ΔIsc by approximately 50%, and the remaining current was abolished by the addition of apical amiloride (Fig. 6B). This result suggests that a significant component of the ATP-evoked current can be due to Cl−-independent HCO3− secretion. This could account for some or all of the bumetanide-insensitive component of the anion current noted earlier.
Discussion
An important requirement for the development of an artificial salivary gland is to construct a secretory epithelium that is able to achieve basolateral-to-apical electrolyte transport followed by the passive osmotic movement of water.3,9,12 The major finding of the present work is that it is indeed possible to form an epithelial monolayer of human salivary cells that is capable of transporting anions from the basolateral to the apical surface. We report in this study that we succeded in growing polarised salivary epithelial cells, which not only formed tight junctions but also achieved vectorial transepithelial electrolyte transport.
Isolation and cell culturing protocols may serve as important selection factors in achieving optimal epithelial cell differentiation, which is essential for successful epithelial gland tissue engineering. 27 Hepato-STIM, a specialized medium originally designed for the culture of hepatocytes, has been successfully used to grow both isolated human primary salivary cells15–17 and rabbit lacrimal acinar cells. 28
Our data confirm that Hepato-STIM and the widely used culture medium, MEM, are both suitable for maintaining primary cell cultures from salivary glands. However, only Hepato-STIM was able to direct both PTHSG and huSMG cells toward confluent, epithelium-like cell monolayers. When cells were cultivated in MEM, fibroblast-like elements dominated the cell population early in cultivation. Futhermore, high TER, consistent with the formation of functional tight junctions, developed only in the presence of Hepato-STIM and not in MEM. In this respect, it is worth mentioning that Hepato-STIM is a Williams' E medium with dexamethasone, insulin, transferrin, selenium, EGF, and 1.8 mM Ca2+. These components are all important factors in maintaining an epithelial phenotype.16,17,29–31
Active ion transport, assessed by Isc measurements, was only weakly related to TER values under our experimental conditions. This is because transepithelial resistance is primarily determined by tight junction formation, which does not directly affect active transcellular electrolyte transport. Widely varying TER values have been measured in previous studies on confluent monolayers of CFPAC, Capan 1, ParC-10, and HPAF cells (∼100, 200, 500, and 2000 Ω cm2, respectively), despite them showing very similar Isc values.32–35
Besides the medium used, the origin of the cells seems to be crucial as well. In the original description, the PTHSG cell culture method omits a step that in the huSMG protocol was intended to select for epithelial cells. PTHSG cultures consequently also contain quickly attaching fibroblast-like cells.16,17 When cultured in Hepato-STIM, the PTHSG and huSMG cell populations show similar morphology, mRNA expression patterns, and basic functional features (monolayer formation, short-circuit current generation). However, in addition to the already reported difficulties with huSMG cell culture (slow growth, insufficient cell number, failure of survival on basement membrane extract [BME]15,17,20), we observed unexplained cell death and poor monolayer formation (low TER levels) more frequently in huSMG than in PTHSG cultures.
These findings suggest that separation of the 1-day-old floating cells from the other cell types is not only unnecessary for producing an epithelium-enriched cell culture but may also have some disadvantages. In line with our observation, recently it has been reported that isolated mesenchymal progenitors gained epithelial phenotype when cocultured with salivary epithelial cells. 15 This study, together with the present one, suggests that epithelial–mesenchymal interactions may result in better epithelium formation in optimized conditions.
A suitable graft cell for the artificial salivary gland should be capable of regulated fluid and electrolyte secretion similar to that achieved by acinar cells in the human salivary gland.11,12,14,36,37 The huSMG cells were orginally reported as duct cells after morphological examination (by transmission electron microscopy) and functional (fluid movement) experiments. 16 We previously showed that PTHSG cultures exhibit relatively moderate acinar characteristics, but considerable ductal phenotype. 17 In the present study, both acinar (NKCC1, AQP5, and amylase) and ductal (ENaC and claudin-1) markers4,8 have been detected in PTHSG and huSMG cultures, and the short-circuit current measurements have provided further evidence for both ductal and acinar characteristics.
During culture, the expression of acinar cell markers progressively declined compared to the intact submandibular gland, suggesting that there is some dedifferentiation, a feature often reported in other primary cell cultures (e.g., ref 38 38 ). Expression levels of amylase and claudin-1 increased, while NKCC1, AQP5, and ENaC levels were sustained after seeding on to Transwell membranes. This suggests that monolayer formation and polarization do not automatically lead to acinar differentiation in either PTHSG or huSMG cell cultures, but significantly shift the cultured cells toward the acinar phenotype.
However, it is worth mentioning that we were able to measure the amylase release from both PTHSG and huSMG cultures. Amylase discharge was higher in cells cultured on Transwells than on cells cultivated on a plastic surface. Furthermore, salivary amylase was discharged exclusively at the apical surface, once again showing salivary cell polarization under these conditions on Transwell membranes. Based on our previous studies, additional activation of the cells, such as pathways activated by Matrigel extracellular matrix application, could lead to further increases in amylase expression and a decrease in claudin-1 expression as we and others have reported previously.17,18
Our finding that the expression of the tight junction protein claudin-1 is increased in cells cultured both on plastic and on Transwell membranes is not unexpected. Although qualitative studies suggested that claudin-1 is expressed in both salivary acini and ducts,16,39 quantitative analyses have shown that its expression in ductal elements is much more abundant than in acini.40–42 In other words, claudin-1 can be regarded as a quantitative rather than a qualitative marker to differentiate between the acinar and ductal phenotype. Data from the present investigation showed that the expression of this protein was increased in both PTHSG and huSMG cultures compared to intact salivary tissue, and the formation of polarised monolayers on Transwells did substantially modify this expressional level compared to plastic surfaces.
In our work, transepithelial electrolyte secretion was detected by short-circuit current measurements. The ability to block the major ductal ion movement (Na+ reabsorption mediated by ENaC) with low concentrations of amiloride allowed us to characterize the acinar function of the monolayers in isolation.30,43 We observed that electrolyte secretion is Cl− and HCO3− dependent. We also found that the HCO3−-dependent component of the secretion is bumetanide insensitive. These data are consistent with the presence of an alternative Cl− uptake pathway that, in addition to NKCC1, requires the coactivity of basolateral NHE1 and AE2.26,29,31,44 The presence of Na+/HCO3− cotransporters (NBC1) at the basolateral side of salivary acinar cells has been described in several studies, but it was unclear whether the HCO3− taken up this way recirculates through AE2, or whether NBC1 provides a HCO3− uptake pathway for transepithelial bicarbonate transport.45–47 In our work, even in the absence of Cl− from the bathing solution (thus inhibiting Cl− uptake/secretion), a considerable amount of anion secretion remained. This residual secretion decreased significantly after applying the NBC1 inhibitor, DIDS, suggesting that there is a net transepithelial HCO3− secretion through this pathway. Taken together, our data show that in both PTHSG and huSMG preparations, the transepithelial electrolyte movement is made up of three principal active components: basolateral-to-apical HCO3− and Cl− movements, characteristic of acinar cells, account for approximately 50%, while apical-to-basolateral Na+ reabsorption, characteristic of ductal cells, accounts for the rest.
In salivary glands, an increase in the intracellular Ca2+ concentration is known to be the primary signal for fluid secretion, while cAMP elevates protein secretion.4,5,8,26 In our work, the Ca2+-mediated agonists, ATP and carbachol, both increased transepithelial anion secretion, while forskolin, used to elevate intracellular cAMP, had no effect. Under physiological conditions, parasympathetic stimulation results in a massive fluid secretion with little protein in the saliva. This activation is associated with the M3 muscarinic receptor.4,5,8,48 As expected, not only carbachol but also ATP increased transepithelial electrolyte current, an effect that has been characterized previously in various salivary cell preparations from diverse species.49–51 In our experiments, ATP stimulation actually produced the largest increases in short-circuit current.
This is a feature shared with many epithelial tissues, including respiratory and renal epithelia. In these tissues, ATP has been reported to work as a paracrine agent activating members of the P2 purinergic receptor family on either the apical or basolateral side of the epithelium. P2 receptors can cause ENaC inhibition or stimulation, depending on the receptor subtype, and may thereby regulate sodium reabsorption. 52 The purinergic activation of ENaC might therefore account for the additional increase in Isc compared with the muscarinic stimulation with carbachol that we observed in our experiments.
The currently accepted model of salivary fluid secretion is based on a large number of molecular and functional studies performed on standard laboratory species such as mice, rats, and rabbits. 44 In recent years, studies using monolayers grown from cell cultures and cell lines have taken over from duct isolation techniques. However, human salivary cells, capable of forming polarized monolayers, have not previously been available for this purpose. 14 Our present study provides evidence that PTHSG and huSMG are not only capable of forming polarized confluent monolayers but are also capable of regulated transepithelial anion secretion.
In conclusion, the greatest challenge in engineering an artificial salivary gland remains to find suitable graft cells that are not only compatible with the human body but also capable of performing salivary electrolyte and fluid secretion. In the present study, we have demonstrated that both PTHSG and huSMG primary cultures cultivated in Hepato-STIM can form two-dimensional monolayers in vitro on permeable supports. These monolayers are indeed capable of performing vectorial transepithelial electrolyte transport, but the outcome currently is a culture exhibiting both acinar and ductal characteristics. Nonetheless, further refinement of the culture methods should ultimately lead to a safe, human-compatible, and more fully differentiated cell source for engineering artificial human salivary glands for therapeutic use.
Footnotes
Acknowledgments
Our research was supported by the Hungarian National Development Agency (TÁMOP-4.2.1/B-09/1/KMR-2010-0001, TÁMOP-4.2.2/B-10/1-2010-0013) and the Hungarian Scientific Research Fund (OTKA-NKTH CK-80928).
Disclosure Statement
No competing financial interests exist.