
Abstract
Successful hair follicle (HF) neogenesis in adult life depends on the existence of both capable dermal cells and competent epidermal keratinocytes that recapitulate embryonic organogenesis through epithelial–mesenchymal interaction. In tissue engineering, the maintenance of trichogenic potential of adult epidermal cells, while expanding them remains a challenging issue. We found that although HF outer root sheath keratinocytes could be expanded for more than 100 passages as clonogenic cells without losing the proliferative potential with a 3T3J2 fibroblast feeder layer, these keratinocytes were unable to form new HFs when combined with inductive HF dermal papilla (DP) cells. However, when these high-passage keratinocytes were cocultured with HF DP cells for 4 days in vitro, they regained the trichogenic ability to form new HFs after transplantation. We found that the short-term coculture with DP cells enhanced both Wnt/β-catenin signaling, a signaling cascade key to HF development, and upregulated the expression of HF-specific genes, including K6, K16, K17, and K75, in keratinocytes, indicating that these cells were poised toward a HF fate. Hence, efficient production of trichogenic keratinocytes can be obtained by a two-stepped procedure with initial cell expansion with a 3T3J2 fibroblast feeder followed by short-term coculture with DP cells.
Introduction
H
Neogenesis of HFs in postnatal life requires appropriate epithelial–mesenchymal interaction that recapitulates the embryonic developmental process. Up to date, postnatal neogenesis of HFs can be generally categorized into three types. The first is wound-induced spontaneous de novo HF neogenesis. HF neogenesis can be observed following large-area full-thickness skin loss in both laboratory mice and wild rodents.2,3 However, this phenomenon was rarely documented in humans. Second, HF neogenesis can be achieved by the combination of embryonic or newborn dissociated epidermal and dermal cells.4–6 These embryonic or newborn cells still preserve the ability to initiate the epithelial–mesenchymal cross talks that enable HF neogenesis. However, this may not be feasible for clinical application because of the lack of autologous embryonic tissues in adult life. The third method for HF neogenesis is to use the adult HF mesenchymal cells, that is, dermal papilla (DP) cells, for HF induction. Both intact DP and cultured low-passage DP cells are able to interact with keratinocytes to generate new HFs.7–10 Newborn keratinocytes are often used due to their high competency to react to the inductive mesenchymal cells.
Generally, adult interfollicular keratinocytes, either freshly isolated or expanded in culture, do not respond well to the inductive signals from adult DP cells, leading to a low efficiency of HF neogenesis.11,12 Successful development of new HFs, therefore, depends on using freshly isolated fetal or newborn epidermal cells.5,13 While culture-expanded adult keratinocytes originated from clonogenic stem cells of outer root sheath have high proliferative ability and self-renewal capacity, they were unable to form new HFs efficiently under the inductive cues from adult DP cells.14–16 This limitation poses challenges in achieving HF organogenesis with adult keratinocytes.
Attempts to gain or restore the capability of certain cell types for regeneration of specific tissues have been tried either through direct conversion with transcriptional factors4,17,18 or by serial induction with defined growth factors or chemicals. 19 Direct cellular reprogramming is an attractive approach to convert specific somatic cells to desired cell types. However, the requirement of viral transfection can be a safety concern in clinical application. Although to expand and maintain cells with preservation of specific functions have been achieved in various cell types with defined growth factors, currently there is a lack of a culture method that can maintain or enhance the competency of adult keratinocytes to form HFs under the inductive cues of adult DP cells. Conventionally, HF stem cells can be extensively expanded and passaged without losing the proliferative ability under an appropriate fibroblast feeder layer in vitro.20–23 However, whether these cultured stem cells can still respond to the inductive cues of cultured DP cells has not been systemically examined.
To prepare sufficient numbers of competent epidermal cells for HF regeneration, it will be necessary to establish methods without losing their differentiation ability in serial passages. Identifying suitable conditions and factors required to maintain the inductive property is therefore crucial for investigation. In the present study, dissociated cells used for HF regeneration were prepared by traditional feeder-supported expansion from adult cells, followed by short-term in vitro contact with inductive dermal cells. This two-stepped cultivation first quickly expanded adult keratinocytes and then conferred them trichogenicity. The cells were assessed for markers of hair differentiation and for the factors that regulate development of the HF. This method allows fast scalable expansion of keratinocytes for HF neogenesis.
Materials and Methods
Animals
All animal experiments in this study were carried out according to the guidelines that were approved by the Institutional Animal Care and Use Committee (IACUC) of the National Taiwan University (Approval No: 20080227). DP cells and keratinocytes were isolated from 6-week-old female Wistar rats (BioLASCO Taiwan Co., Ltd.). Hair neogenesis patch assay was performed on 5-week-old female nude mice (BALB/cAnN-Foxn1nu/CrlNarl, National Laboratory Animal Center, Taiwan). The animals had free access to water at any time and were maintained under light–dark cycles.
Cell culture
DP cells were isolated from vibrissae of rats as described previously. 24 DP cells at passage 3 (p3) were used in the following coculture experiments and patch assays for HF neogenesis. Keratinocytes were isolated from the outer root sheath of rat vibrissae using the following steps. Using fine forceps, the vibrissae were dissected out from the pad individually after removing the subcutaneous fat and connective tissue around the capsule. Two transversal cuts were made to each of the dissected vibrissae. The first cut removed the upper part, including the superior blood sinus and sebaceous gland. A second cut was made in the middle of the remaining part. The middle part was reserved in Hank's buffered salt solution (Gibco). All reserved fragments were then transferred into the medium containing 1.25 U/mL dispase (Gibco) and 0.5 mg/mL collagenase I (Sigma-Aldrich) at 37°C for 30 min. After the incubation, the shaft and the epithelial cells distributed on outer root sheath (ORS) inside the capsule could be easily pulled out and incubated in a solution of 0.05% trypsin/EDTA (Gibco) for 1 h at 37°C. During this period, pipetting the solution gently every 20 min was done till the process of dissociation was completed. The keratinocytes were obtained by filtering the cell suspension through a sterile 40-μm cell strainer (BD Biosciences). Cells were resuspended in a 10-mL keratinocyte growth medium (KGM) in a 10-cm culture dish (Nest). The 3T3J2 fibroblast cells, kindly provided by Prof. Howard Green (Harvard Medical School, Cambridge, MA), were treated with γ-irradiation of 60 Gy from the cesium source (Cesium-137 gamma irradiator) and seeded as a feeder layer with keratinocytes simultaneously. The keratinocytes were serially subcultured with 3T3J2 cells and we used passage 40 (p40) to passage 50 (p50) generations in the following experiments.
KGM is a 3:1 mixture of Dulbecco's Modified Eagle's Medium (DMEM) and Ham's F12 media (Gibco), containing 10% fetal bovine serum (Biological Industries), 5 mg/mL insulin (Sigma-Aldrich), 0.18 mM adenine (Sigma-Aldrich), 0.4 mg/mL hydrocortisone (Sigma-Aldrich), 0.1 nM cholera toxin (Sigma-Aldrich), 2 nM triiodothyronine (Sigma-Aldrich), 10 ng/mL epidermal growth factor (Gibco), and 1% antibiotic–antimycotic liquid (Gibco). 25 The 3T3J2 was cultured in DMEM containing 10% bovine calf serum (Thermo Scientific HyClone) and 1% penicillin–streptomycin solution (Gibco).
Preparation of cocultured cells
DP cells and keratinocytes were cocultured in KGM for 4–5 days. The ratio of DP cells and keratinocytes for coculture was 1: 2. Approximately 3×105 DP cells and 6×105 keratinocytes were seeded in a 10-cm culture dish (Nest). The cells were cocultured for up to 5 days, and the medium was changed on day 3 and 4.
Patch assay for HF neogenesis and histological examination
Cultured DP cells (p3) were combined with high-passage outer root sheath keratinocytes (p40–50). Approximately 1 million DP cells and 1 million epidermal cells were mixed at a concentration of 104 cells/μL in DMEM/F12 and were injected into the subcutis of nude mice (BALB/cAnN-Foxn1nu/CrlNarl). 5 Three weeks after injection, the skin was harvested and processed for histological analysis. We inserted a rough glass disc into the hypodermis of anesthetized nude mice 7 days before patch model cell injection to enhance the granulation tissue bed and blood vessel formation. 10 The disc was removed right before the patch assay injection procedure. Each patch assay using either untreated high-passage keratinocytes or cocultured keratinocytes, or neonatal epidermal cells was repeated for at least 10 times.
Histology and immunostaining
DP cells and keratinocytes were cocultured in the chamber slide (Lab-Tek) and were collected on day 0 through day 4. Cells were rinsed with phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde for 5 min, permeabilized with 0.3% Triton X-100 for 10 min, and then washed thoroughly with PBS. The cells were then blocked with 5% bovine serum albumin (BSA) in PBS for 30 min, followed by incubating with the mouse anti-β-catenin antibody (1:500 in1% BSA/PBS) (BD Biosciences) overnight at 4°C. This primary antibody was washed off with 1% BSA in PBS for 5 min twice and then incubated with mouse IgG DyLight 488 (GeneTex) for 1 h at room temperature at a dilution of 1: 300 (diluted in 1% BSA/PBS). This secondary antibody was washed off with 1% BSA in PBS for 5 min twice, and nuclei were labeled with 1 μg/mL 4′,6-diamino-2-phenylindole (DAPI) (Sigma-Aldrich) for 5 min. Slides were rinsed with PBS for 5 min twice and then mounted with a fluorescence mounting medium (Dako). Fluorescence was detected by a confocal microscope (Meta510; Carl Zeiss), and images were analyzed by Zeiss ZEN confocal software (Carl Zeiss Microscopy GmbH).
Quantitative polymerase chain reaction
Quantitative real-time PCR (qPCR) was performed to analyze specific gene expression in cocultured keratinocytes. After coculture, 0.02% EDTA (Invitrogen) was used to remove DP cells to collect the keratinocytes. The cells were further processed for qPCR analysis as described. 12 The following program was designed for all the qPCRs: (I) 95°C for 15 min ×1 cycle and (II) 95°C for 15 s, 60°C for 1 min×40 repeat cycles. The data were calculated with the 2−ΔΔCt method, with Gapdh as the reference gene for normalization.
Genes for analysis include the DP signature gene of Vimentin (Vim), epidermal differentiation markers of keratin 1 (Krt1), maker for basal cells in the epidermis and HF of keratin 14 (Krt14), outer root sheath marker of keratin 6 (Krt6) and keratin 17 (Krt17), follicular differentiation marker of keratin 16 (Krt16), companion layer marker of keratin 75 (Krt75), and the Wnt signaling pathway marker for HF development of β-catenin (Ctnnb1) and were analyzed.
The sets of primers and probes were designed as follows: Gapdh: 5′-tcaacgaccccttcattgac-3′ and 5′-ttcccattctcagccttgac-3′; Vim: 5′-tttgaagaagctgcacgatg-3′ and 5′-acatcgatctggacatgctg-3′; Krt1: 5′-gaagccgagaccttttacca-3′ and 5′-tccgtgtttaccagcagtga-3′; Krt14: 5′-tgagagcctcaaggaagagc-3′ and 5′-cctctcatggaagccatttc-3′; Krt6: 5′-gcgttggaccagtcaacatc-3′ and 5′-ctgcactgccatagccact-3′; Krt17: 5′-ccagagctgacctggagatg-3′ and 5′-actgatcccgcatctctgac-3′; Krt16: 5′-aggacttggtggtggattt-3′ and 5′-gtaccagtcacggatcttca-3′; Krt75(K6hf): 5′-ctgtctgtccctctggaagc-3′ and 5′-tgatctgttcccgctcttct-3′; Ctnnb1: 5′-gaccacaagcagagtgctga-3′ and 5′-ctgactcgggtctgtcaggt-3′
Western blotting
For cell fractionation into nuclear and cytoplasmic content, the untreated and cocultured cells were pelleted and lysed with the NE-PER Nuclear and Cytoplasmic Extraction Kit (Thermo Scientific) supplemented with Protease Inhibitor Cocktails (Thermo Scientific) according to the manufacturer's instructions. The proteins were stored at −80°C until use. Protein lysates were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrotransferred onto a Hybond-ECL nitrocellulose membrane (Bio-Rad Laboratories). The membrane was blocked with 5% nonfat milk in phosphate-buffered saline with Tween® 20 (PBST-20X) and incubated with anti-β-catenin (1:1000) overnight at 4°C. After washing with PBST-20X for 10 min thrice, the membrane was incubated with HRP-conjugated goat anti-mouse IgG (CHEMICON) (1:1000) for 1 h at room temperature. After that, the membranes were thoroughly washed with PBST-20X and developed with the WesternBright ECL HRP substrate (Advansta). The signal intensity was detected and measured by ImageJ software (National Institutes of Health [NIH], Bethesda, MD).
Statistical analysis
Student's t-test was used to analyze keratinocyte gene expression. qPCR data were analyzed after being normalized for actin loading control. Western blotting quantitative analysis was carried out by the fold change of β-catenin intensity of the keratinocytes before and after coculture with DP cells. Statistical significance was determined if two-sided p<0.05.
Results
Expansion of keratinocytes with a 3T3J2 fibroblast feeder and the loss of trichogenic ability
To expand the population of epidermal cells for HF neogenesis, culturing primary cells in serial passages provides an efficient method for large-scale production. We took keratinocytes from the vibrissae of 6-week-old adult Wistar rats, cultivated these cells, and passaged them as previously described.12,21,22 Lethally irradiated 3T3J2 cells provided a supporting environment for keratinocyte attachment and growth. Under this condition, the keratinocytes replicated as colonies and were tightly packed and adhesive to the culture plate (Fig. 1a). The colonies were surrounded by feeder 3T3J2 cells, and most cells in the colonies remained small and uniform. The rare presence of large flat (squame-like) cells inside or surrounding the colonies indicated that most of the keratinocytes did not undergo terminal epidermal differentiation.20,26 Colonies from the low passage to high passage (up to passage 100) preserved almost a uniform morphology and exhibited a high proliferative activity (Fig. 1a–g), suggesting that these colonies were most likely to be holoclones with stem cell activity rather than meroclones or paraclones that have only limited replicative potential. 20 Hence, under such culture conditions, keratinocytes from both the HF outer root sheath could be serial passaged and expanded and the high-passage keratinocytes still preserved the reproductive capacity and retained their growing characters such as holoclones, indicating that they are more primitive or progenitor like.
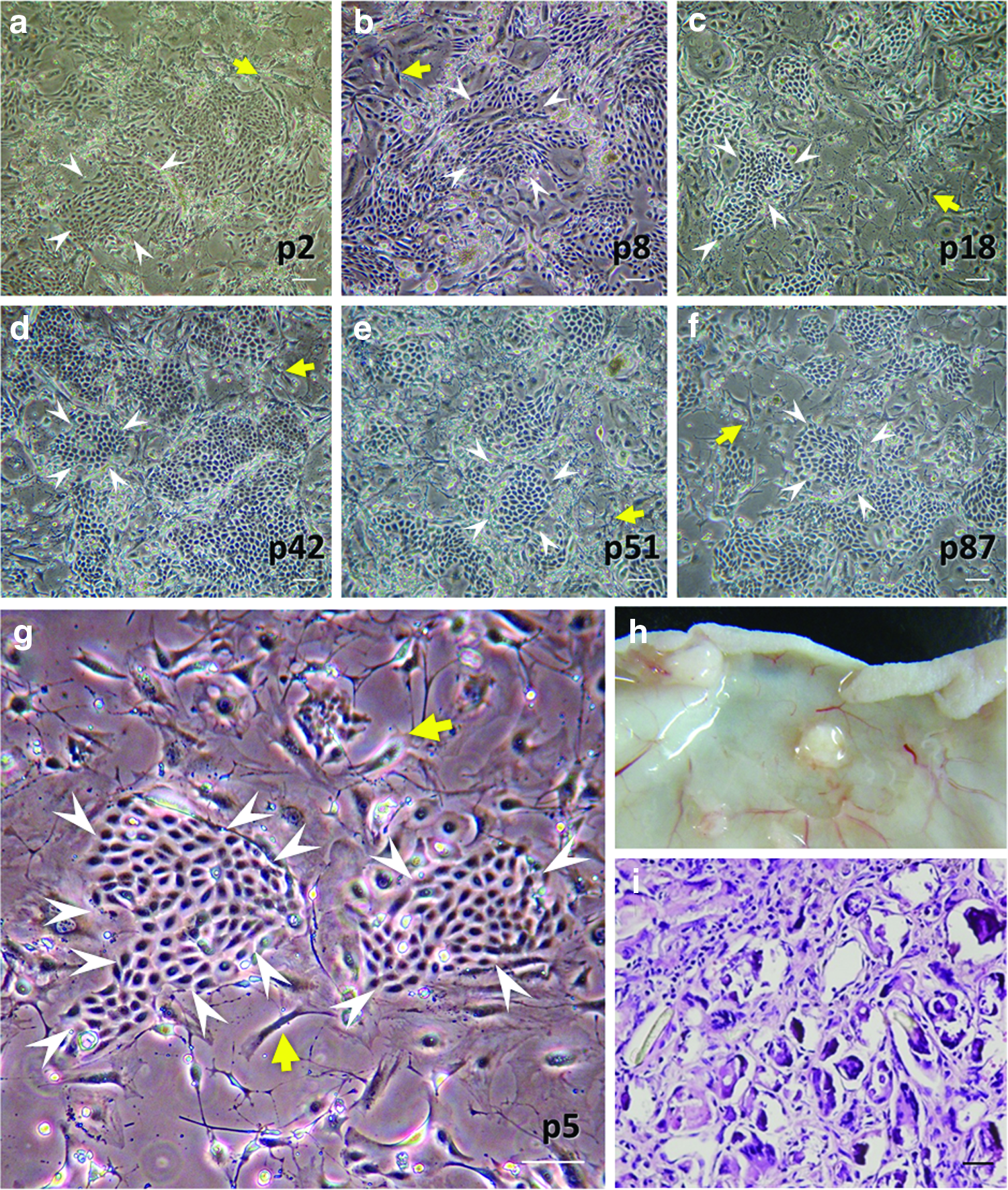
High-passage keratinocytes maintained proliferative potential, but not hair follicle (HF)-forming capability.
We then tested whether the cultured keratinocytes could form new HFs in response to adult DP cells. We found that further in vivo follicular cell implantation animal experiments combining hair-inducible cultured DP cells with these high-passage keratinocytes were not able to form new hair (Fig. 1h, i).
Coculturing with DP cells conferred high-passage adult keratinocyte HF forming ability
Our previous report showed that when freshly isolated adult foot pat keratinocytes were in contact with cultured adult DP cells in a spheroid, they were able to form new HFs. 12 It suggests that intimate interaction with DP cells may confer adult keratinocyte trichogenic ability. Hence, we tested whether trichogenesis of high-passage clonogenic keratinocytes can be enhanced through interaction with DP cells. Coculture with DP cells did not overtly alter epidermal cell morphology (Fig. 2a). When tested in the patch assay, we found that the cocultured high-passage keratinocytes exhibited hair-forming capability (Fig. 2b). Further examination of the hair fibers by the scanning electron microscope confirmed that these fibers were true hair shafts with typical overlapping cuticles (Fig. 2c). Multiserial ladder medullation of the shafts that frequently exists in rodent hairs was observed in the histological examination (Fig. 2d). Compared with control high-passage keratinocytes, these cocultured high-passage keratinocytes showed a rather high hair induction rate (Table 1). Three out of four experiments yielded positive hair neogenesis.
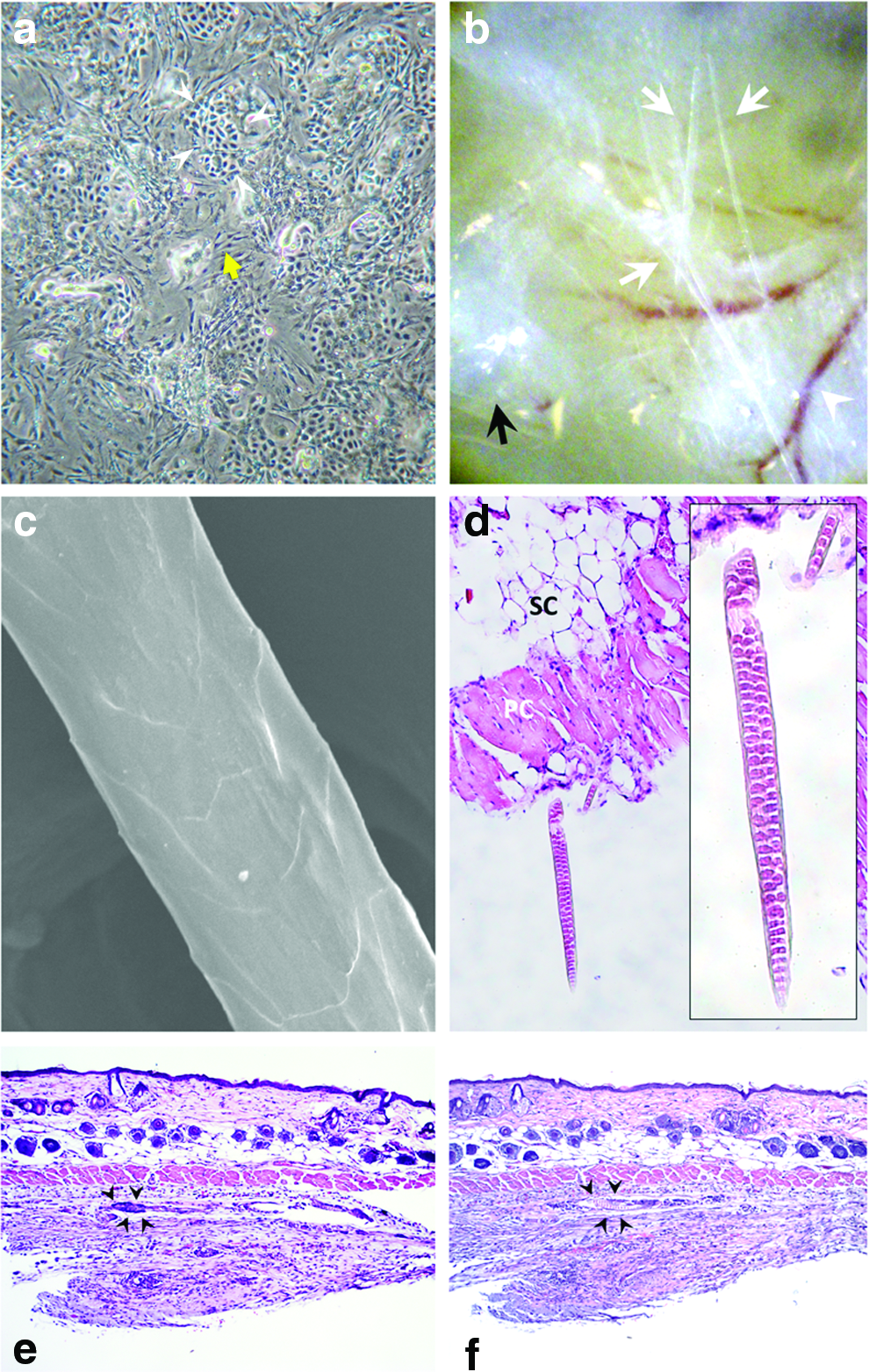
Short-term coculturing of high-passage keratinocytes with DP cells enhanced HF-forming capability.
DP, dermal papilla.
Expression of follicle-specific genes were upregulated in cocultured keratinocytes
The findings above showed that coculture of keratinocytes with adult DP cells explicitly changed the HF-forming capability of the high-passage keratinocytes. To understand how these cells became poised for HF formation, we analyzed the changes of gene expression (Fig. 3). Generally, epidermal keratinocytes have two fates during embryogenesis: interfollicular fates and follicular fates. We found coculture with DP cells increased the expression of HF-specific genes. The gene expression of HF differentiation markers for the outer root sheath (K6, K16, K17) and companion layer (K75/K6hf) showed a coculture time-dependent increase in the mRNA levels.27–31 Induction of K16, K17, and K75/K6hf expressions indicated the differentiation of cultured epidermal cells into follicular lineages.31,32 Among these genes, the hair-specific keratin K75/K6hf exhibited a rather high-fold increase in the gene expression level, corresponding to an earlier study demonstrating that DP may enhance HF differentiation of epidermal stem cells by paracrine factors. 28
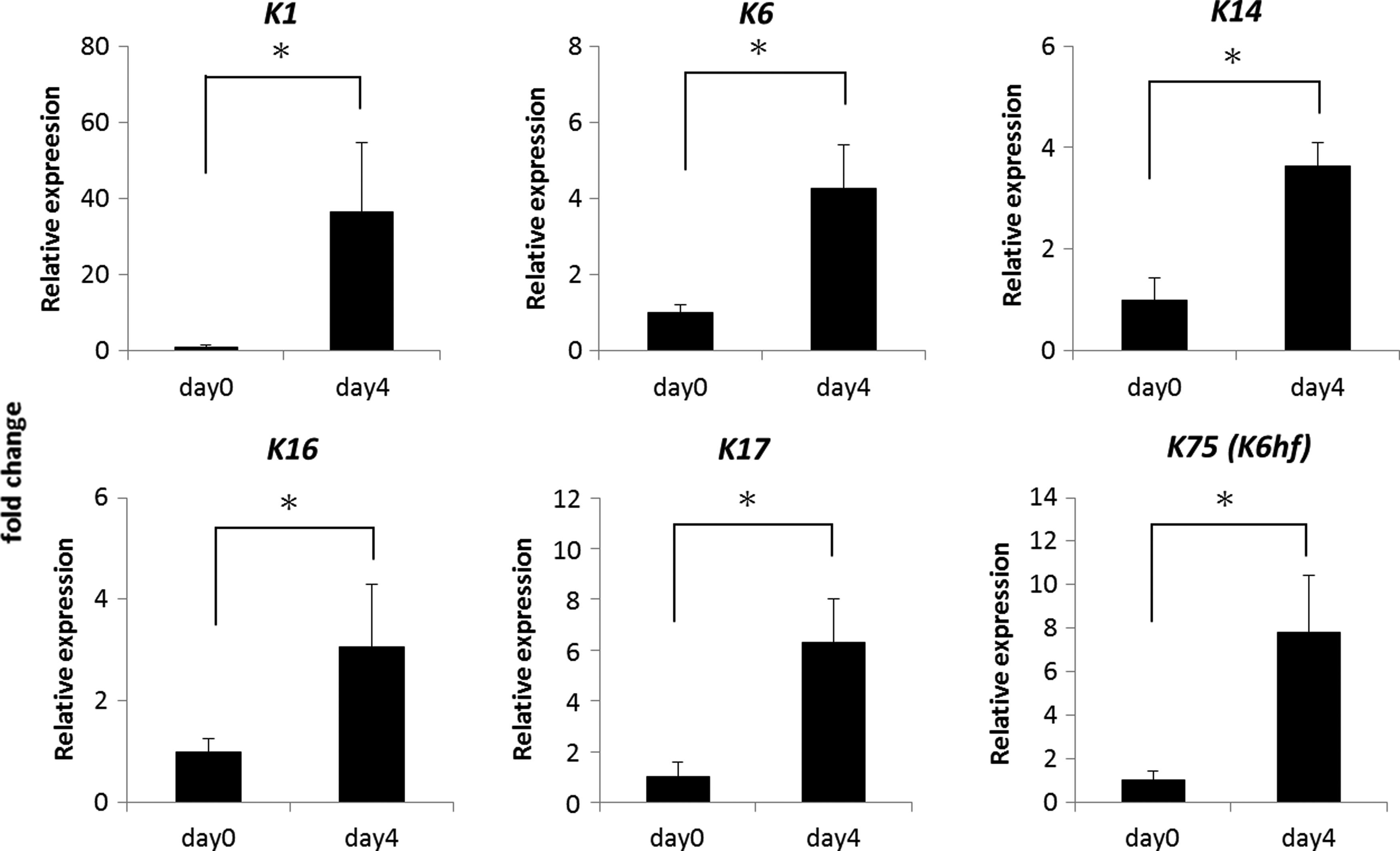
Upregulation of hair differentiation genes of cocultured epidermal cells. Follicular differentiation gene expression for ORS (K6, K16, K17) and companion layer (K75/K6hf) demonstrated a synchronously increasing trend. The nonfollicular epidermal differentiation marker, K1, was also elevated during coculture. The basal keratinocyte marker of K14 was also upregulated. *p<0.05.
We also noted that the interfollicular epidermal-type differentiation marker, K1, was also increased during coculture. This finding suggests that while cocultured epidermal cells received signals from the DP cells to initiate hair differentiation, differentiation potential toward an epidermal fate was still maintained. These cocultured cells were bipotent in generating an epidermal and follicular structure.
Increased β-catenin expression and activated Wnt/β-catenin signaling in cocultured epidermal cells
The canonical Wnt/β-catenin signaling pathway plays a key role in HF morphogenesis and development.33–35 Without Wnt/β-catenin signaling, HF development was abolished. 36 Postnatal activation of β-catenin signaling in epidermal keratinocytes has been shown to increase folliculogenesis. 33 We wonder whether Wnt/b-catenin signaling was enhanced in cocultured epidermal cells.
To study the changes of β-catenin levels, the principal mediator of the canonical pathway of Wnt signaling, cocultured epidermal cells were isolated for determination of gene and protein expression of β-catenin. As shown in Figure 4a, β-catenin mRNA was upregulated in a time-dependent manner in epidermal cells from day 0 to 4. Consistent with this, we also found that the total β-catenin protein was also increased during coculture (Fig. 4c). The results showed that coculture with DP cells increased the expression of β-catenin expression both in the RNA and protein level in high-passage keratinocytes.

Enhancement β-catenin expression and Wnt/β-catenin signaling pathway by coculture with DP cells.
We also determined whether Wnt/β-catenin signaling was activated by coculture. Accumulated and stabilized β-catenin translocates from the cytosol to the nucleus where it associates with the transcription factors LEF/TCF to form a transcriptional complex that promotes the expression of Wnt-regulated target genes responsible for cell proliferation and differentiation.37–41 Therefore, retention of β-catenin in the nucleus is direct evidence of an activated Wnt/β-catenin signaling pathway. To examine the nuclear translocation of β-catenin, the cytoplasmic and nuclear extracts from the epidermal cells were subjected to Western blot analysis. We found that the ratio of nuclear β-catenin increased after coculture with DP cells (Fig. 4b, d). This was also supported by the immunofluorescence detection of increased localization of β-catenin (Fig. 4e). The results showed that coculture with DP cells not only increased the expression of β-catenin but also activated Wnt/β-catenin signaling in high-passage keratinocytes.
Discussion
Hair loss due to aging or other pathological changes can now be treated with surgical hair transplantation by redistributing remaining HFs to the hairless area or with medications that target causative underlying pathological pathways such as oral finasteride in male baldness. 42 These methods do not increase HF numbers and are only effective for mild to moderate hair loss. In patients with severe hair loss, neogenesis of new HFs can be a potential treatment alternative. In patients with a large area of skin defects due to injuries, such as burn and trauma, although a skin equivalent derived from autologous epidermal cells and fibroblasts can be used to repair the skin defects, no skin appendages, including HFs, can be regenerated. Hence, the ability to regenerate HFs through tissue engineering can help to meet the current need. Hence, in both hair loss and injury-induced skin defects, how to reconstitute the fetal epithelial–mesenchymal interaction by use of adult cells is the key to the neogenesis of HFs.
Considering the large numbers of new HFs needed, expansion of inductive dermal cells and competent epithelial cells is required. For dermal cells, it has been shown that DP cells quickly lose the inductive ability after expansion by serial passages in vitro.6,43,44 Hence, only low-passage DP cells can be used to regenerate HFs, and culturing low-passage DP cells into multicellular aggregates can help to enhance the inductive ability.12,13,24,45,46 To maintain the inductive property of DP cells while expanding them, it has been shown that enhancing Wnt/β-catenin signaling from the start of in vitro culture can help to maintain the inductive property of DP cells. 47 On the contrary, once the HF-inducing property is lost in high-passage DP cells, reintroduction of Wnt/β-catenin signaling is unable to convert them back into inductive cells. 48
Physiologically, DP cells are in close contact with HF matrix germinative cells in vivo, and maintenance of the interaction of DP and HF matrix germinative cells is important for proper hair growth. It is intriguing to test whether these matrix germinative cells are able to maintain or enhance the inductive property of DP cells in vitro. It was demonstrated that high-passage DP cells that were unable to induce HF neogenesis regained the inductive property after coculture with freshly isolated germinative HF cells in vitro. 49 It implies that reconstitution of the in vivo heterotypic intercellular interaction may help to restore the original properties of high-passage cultured cells. In line with this, it has also been shown that reconstitution of proper heterotypic intercellular interaction can help to maintain progenitor cell characters while expanding them in vitro. 50
In this work, we hypothesized that DP cells can enhance the competency of high-passage HF outer root sheath keratinocytes to form new HFs. The ability of organ-specific mesenchymal cells to instruct the growth and differentiation epithelial cells has been shown. Organ-specific mesenchymal cells have also been shown to be able to convert epithelial cells of other organs into epithelial cell types of the organs where the specific mesenchymal cells are derived. 51 For example, corneal mesenchymal cells are able to instruct skin keratinocytes to differentiate toward corneal keratinocytes. 52 We showed that short-term coculture with DP cells increases the competence of high-passage keratinocytes to form new HFs. In our method, keratinocytes were first expanded with the feeder layer of 3T3J2 fibroblasts. This fibroblast cell line has been widely used in culturing the epithelial of skin and nonskin origins.21,22,53–55 It supports the growth of colony-forming cells from both HFs and the interfollicular epidermis. In addition, it also supports the growth of corneal epithelial keratinocyte stem cells that have been used to repair corneal epithelial defects clinically.54,56 It enables quick and long-term expansion of HF outer root sheath keratinocytes. Compared with 3T3J2 fibroblasts, DP cells were unable to support fast and long-term expansions of HF outer root sheath keratinocytes (S.J. Lin, unpublished data). However, 3T3J2 fibroblasts do not maintain the competency to form HFs. High-passage keratinocytes generated under this feeder layer lost the ability to respond to the inductive activity of DP cells (Fig. 1). This two-stepped culture allows first a quick expansion of keratinocytes. Once the desired cell number is obtained, these cells are then cocultured with DP cells to enhance their competency to form HFs.
The interaction between high-passaged keratinocytes with DP cells can involve dynamic molecular interaction through secreted factors and intercellular contact. Compared with cells expanded under the feeder layer of 3T3J2 fibroblasts, we found that Wnt/β-catenin signaling in high-passaged keratinocytes was enhanced by the coculture with DP cells. Although 3T3J2 fibroblasts maintained clonal growth of outer root sheath keratinocytes, they did not promote Wnt/β-catenin signaling. Wnt/β-catenin has been shown to be essential for the morphogenesis of HFs, activation of HF stem cells, and differentiation of matrix cells into hair shafts. However, Wnt/β-catenin is not required for the maintenance of HF stem cell. 57 3T3J2 fibroblasts may be capable of supporting the clonal growth of HF stem cells, but they are unable to promote HF differentiation. However, this unique property helps to maintain the stemness of the cultured keratinocytes, enabling long-term expansion and preserving the proliferative potential.
After short-term coculturing with DP cells, high-passage keratinocytes exhibited differentiation toward HFs that was evidenced by the expression of HF-specific keratins, including K6, K16, K17, and K75 (Fig. 3). We also found that epidermal differentiation markers of K1 were still maintained. It is possible that part of the cells still maintained epidermal phenotypes, while other cells started differentiating toward a HF fate. Another possibility is that coculture with DP cells brought these cells to an intermediate bipotent state that exhibited differentiation toward both the epidermal and follicular fates. Such changes increase the competency to cross talk with inductive DP cells after transplantation in vivo, enabling the formation of new HFs.
Conclusions
Taken together, this study showed that coculture of high-passage epidermal cells with DP cells enables epidermal cells to undergo trichogenesis, and this process is associated with activated Wnt/β-catenin signaling. This simple two-step method allows an initial quick expansion of clonogenic keratinocytes. Then, the short-term in vitro contact with DP cells shifts the keratinocytes to a state that is competent to receive inductive signals from adult DP cells for HF neogenesis (Fig. 5). This enables scalable production of competent epidermal cells for cell-based treatment of hair loss.
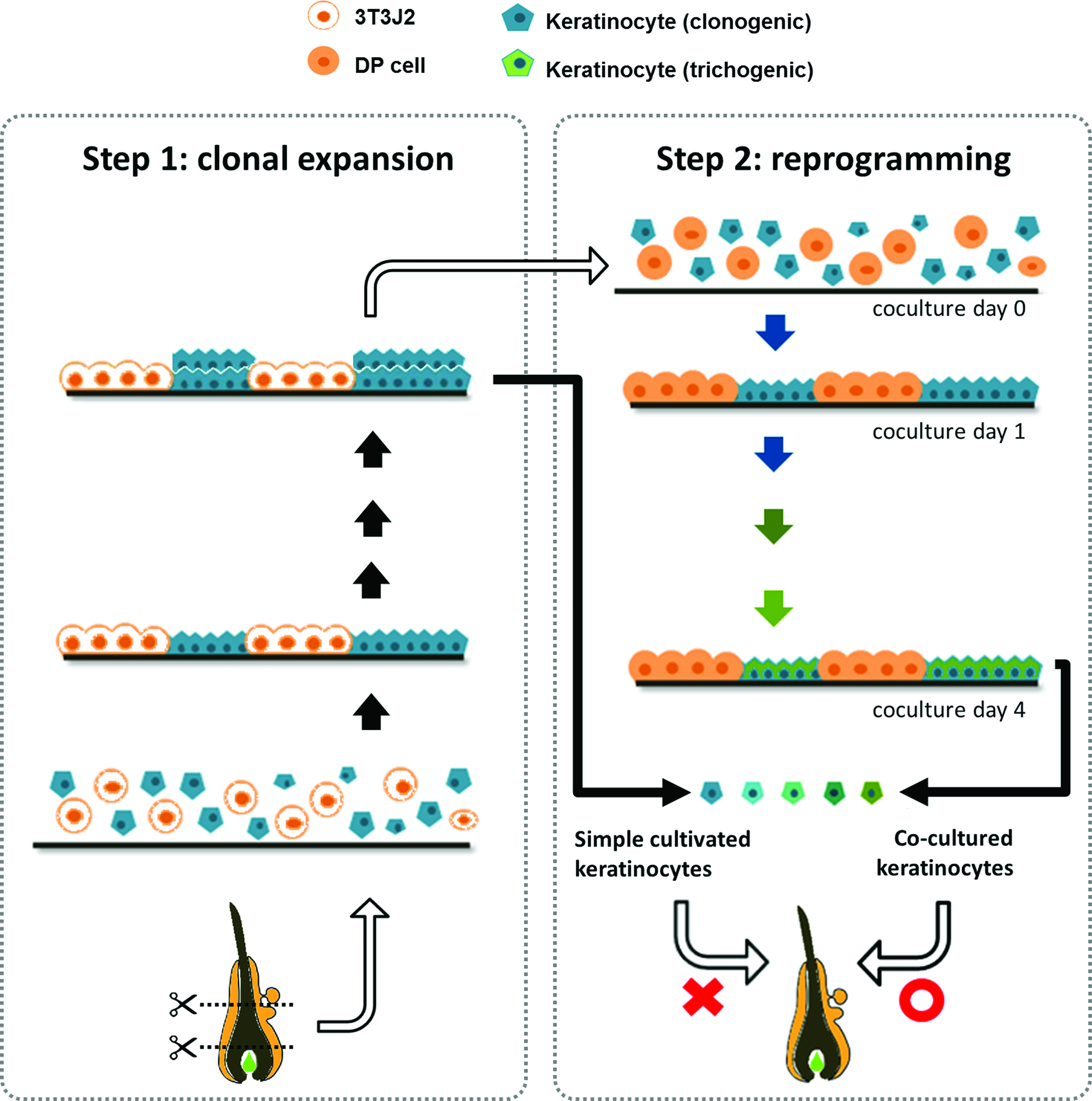
Two-step preparation of trichogenic cultured adult keratinocytes. Step 1 (left panel): population expansion by means of traditional feeder supporting cultivation from clonogenic ORS keratinocytes. Step 2 (right panel): reprogramming takes place, while cultured DP cells enable high-passage epidermal cells to adopt follicular fate with successful expression of follicular-specific genes. These educated and reprogrammed epidermal cells show their ability of folliculogenesis in hair regeneration models. Color images available online at
Footnotes
Acknowledgments
The authors would like to thank Ms. Yi-Fan Chan and Chien-Mei Yen for their technical support on cell culture and animal experiments. They thank the staff of the Eighth Core Laboratory, Department of Medical Research, National Taiwan University Hospital, for their technical support. The authors also acknowledge Dr. June-Tai Wu and Dr. Chia-Yu Chu for fruitful scientific discussions during all phases of this study, and Mr. Pei-Kai Hung for his inspirational support. This work was supported by grants from the Taiwan Ministry of Science and Technology (NSC98-2314-B-002-027-MY3, NSC99-2320-B-002-004-MY3, MOST-103-2325-B-002-008, MOST-102-2314-B-002-025-MY3), the National Taiwan University Hospital (103-S2437, 100-FTN11, 102-M2279, 103-M2583, 104-M2902), and the Taiwan National Health Research Institutes (PS9803).
Disclosure Statement
No competing financial interests exist.