
Abstract
The α-Gal (Galα1,3-Galβ1-4GlcNAc-R) epitope, the major xenoantigen, is the first barrier in a porcine-to-man tissue and organ xenotransplantation. The elimination or reduction of the α-Gal epitopes is therefore an important step for a successful xenotransplantation. The present study is to evaluate the α-Gal elimination in the porcine skin with α-galactosidase treatment, and to assess two methods (immunohistochemistry and inhibition ELISA) that may be used in quality control for quantifying the extent of the α-Gal elimination. Enzymatic cleavage in a single-step process is extremely efficient and affordable at eliminating the α-Gal epitope even in a tissue as dense as the porcine dermis. The cost of enzymatic cleavage is found to be less than US$7 for a 10 × 10 cm piece of porcine skin (0.5 mm thick) or about US$140 for 100 g of 3-dimensional soft tissues. After enzymatic cleavage, the α-Gal-positive immunostaining was essentially undetectable in enzyme-treated porcine skin. The inhibition rate constant of the monoclonal anti-Gal antibody M86 binding to α-Gal-bovine serum albumin in ELISA was reduced from 15.0 ± 4.3 (n = 10) to 6.1 ± 2.6 (n = 7) after enzyme treatment, in comparison to 4.4 ± 1.8 (n = 9) background inhibition of decellularized human skin (the ultimate negative control), which demonstrates ∼84% elimination of α-Gal epitopes in treated porcine skin. To examine the suitability of two detection methods for the routine quality control application, comparative studies were made with control and enzyme-treated porcine skin, porcine skin from the α-Gal knockout animal, as well as decellularized human skin. The data show that the traditional immunohistochemistry and, to a less extent, the inhibition ELISA with further modifications can be used as quality control tools in the production and selection of biocompatible bioprosthetic devices. The biological evaluation of enzyme-treated porcine skin is ongoing with a small animal model and a nonhuman primate model.
Introduction
T
Therefore, the presence of the α-Gal epitope is the first barrier in the animal-to-man tissue and organ xenotransplantation. In recent years, another promising approach for functional tissue and organ replacement has emerged, where decellularized three-dimensional animal tissue and organ scaffolds are seeded with selected human cell populations in an attempt to make functionally transplantable tissue/organ devices.5–8 While the earlier results are promising, significant challenges and critical issues remain to be resolved, including a better understanding of the components and functional properties of decellularized tissue/organ scaffolds. Decellularization removes the α-Gal epitope on the cell surface, but not these on the extracellular matrix scaffolds.
Since the clinical use of α-Gal-positive bioprosthetic materials can induce an anti-Gal antibody response, several approaches have been made to ensure the biocompatibility and safety when animal-based tissue devices are transplanted into humans. The strategies for inactivation, removal, and/or reduction of the α-Gal epitope include chemical crosslinking or shielding, 9 chemical degradation with oxidative and caustic agents, decellularization, genetic engineering using the knockout (KO) expression technology,10,11 as well as specific enzymatic cleavage using purified α-galactosidase.12–14 The prolonged survival of GalT-KO (α1,3-galactosyltransferase gene-knockout) porcine skin and heart valves in nonhuman primates, relative to the Gal-positive tissues, confirms that bioprosthetic devices with this major xenoreactive antigen eliminated, reduced the potential of immune-mediated degeneration and failure of transplanted grafts.15,16
Considering the commercial manufacturing of nonimmunogenic animal-based tissue and organ grafts for human clinical uses, the enzymatic cleavage with α-galactosidase treatment has a number of advantages. Various chemical methods that are mentioned above for reducing immunogenicity typically cause structural and functional damages in extracellular matrices due to their nonspecificity, resulting in the inferior grafts. The genetic engineering approach using the KO technology still faces tremendous regulatory and quality control issues in animal production, not to mention the prohibitive cost. The biology of α-galactosidase has been well studied, it is already commercially available, and the enzymatic treatment step can be integrated with ease into the existing processes used in tissue engineering. The incorporation of specific enzymatic cleavage into a manufacturing process would be less damaging to tissues and organs than other existing nonspecific chemical treatments, yielding high-quality products, and yet much more cost-effective than the KO genetic engineering methods with many regulatory and quality control issues.
In the present study, we have evaluated the α-Gal epitope elimination in the porcine skin with α-galactosidase treatment, and to assess two methods (immunohistochemistry and inhibition ELISA) that could be used in the quality control for quantifying the extent of the α-Gal elimination. This article provides methods and necessary data that can be used to optimize the α-Gal epitope elimination in animal tissue/organ decellularization processes and to measure the extent of the antigen removal from the tissue scaffold.
Materials and Methods
Materials
Alpha-galactosidase (recombinant) from Bacteroides fragilis was produced by our laboratory. 17 The α-Gal-bovine serum albumin (BSA) was from Dextra. M86, a monoclonal antibody to α-Gal epitope was purchased from Enzo Life Sciences. Alpha-galactosidase from green coffee bean and FITC-labeled GSI-B4 were purchased from Sigma-Aldrich, Inc. Decellularized porcine skin was obtained from Chongqing Datsing Yicheng Bio-Tech Co., Ltd. GalT-KO porcine skin was a gift from Dr. Deng-Ke Pan at the Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing, China. Decellularized human skin (AlloDerm) was a gift of LifeCell Corporation.
Alpha-galactosidase treatment of porcine skin
Porcine skin was cut to the dimension, 5 × 5 cm × 0.5 mm. After cleansing with sterile saline, skin samples were treated with purified recombinant α-galactosidase in phosphate-buffered saline (PBS, pH 7.4) to eliminate the α-Gal epitopes in the tissue for 20 h at 20 units/L and 26°C. Skin samples that were incubated in PBS without the enzyme were used as the control for comparison. To assess the extent of α-Gal epitope removal, all skin samples were divided into three groups randomly. One group was imbedded with paraffin for immunostaining, other was cryosectioned for immunofluorescence, and the third for ELISA detection. In another experiment, porcine skin samples (0.6–4.0 mm thick) were incubated with the commercially available α-galactosidase from green coffee beans (Sigma-Aldrich, Inc.) at 50 and 150 units/L for various durations up to 24 h at 23°C to evaluate the efficacy and the cost of α-galactosidase treatment for scale up production (∼600–800 g, or 12,000–16,000 cm2).
Immunohistochemical analysis of paraffin sections
After fixation in 4% paraformaldehyde for 72 h, skin samples were dehydrated in a graded series of ethanol and embedded in paraffin. Histological sections (3 μM) were deparaffinized. The endogenous peroxidase activity was quenched by incubation with 0.3% H2O2 at room temperature for 15 min. Nonspecific binding was blocked by incubating sections in 5% BSA in PBS at room temperature for 30 min. Tissue sections were incubated with the primary antibody of M86 (1:5) overnight at 4°C. After washing, sections were incubated with a secondary biotinylated goat anti-mouse IgM antibody working solution (ZSGB-BIO) for 30 min. Horseradish peroxidase (HRP)-labeled streptavidin working solution (ZSGB-BIO) was reacted with the second antibody for 30 min. Finally, sections were washed three times with PBS and the slides developed in diaminobenzidine (DAB) solution. Additionally, Hematoxylin was used to counterstain the nuclei of the cells in the specimens.
All the sections were visualized and photographed by a light microscope (Olympus IX71). Photographs were taken at a magnification of 200× for analysis. As a negative control, sections were stained with PBS replacing M86 antibody. Alpha-Gal expression was analyzed with Image-Pro Plus software (IPP 6.0, Media Cybernetics). Brown staining on the skin represented positive staining, and staining density indicated the expression level of α-Gal. Ten images of each sample were captured randomly at 400 × magnification. The parameters measured included mean density, area sum, and integrated optical density.
Immunofluorostaining of cryosections
Skin samples were embedded in an optimum cutting temperature compound for cryosections. Briefly, sections were submerged in cold acetone for 5 min and blocked with 5% BSA for at least 1 h at room temperature. The cryosections were stained with two probes, respectively. One group was stained with FITC-labeled GS-IB4 lectin (20 μg/mL), and another group was stained with monoclonal antibody M86 (1:5) overnight at 4°C. Control sections were incubated overnight at 4°C with PBS. After washing, secondary antibody (Alexa Fluor 488 goat anti-mouse IgM, 1:100) was incubated in blocking solution for 1 h at room temperature. Then, the sections were washed to remove nonspecific fluorescence and cover slipped with glycerin. All sections were observed using a fluorescence microscope (Leica) (Olympus).
In the efficacy study, sample slides were first incubated with the biotinylated GS-IB4 lectin before development in DAB. Using the untreated porcine skin as the positive control ( = 100% staining) and the decellularized human skin as the negative control ( = 0% staining), images of various samples at the 100× magnification were digitized and quantified for the presence of residual α-Gal epitopes. Images in a RGB format were loaded in MATLAB and converted to grayscale images. Morphological top-hat filtering was performed on the grayscale images to correct uneven illumination. Morphological bottom-hat filtering was performed to enhance contrast and subtract the background from the images. Edges in the image were detected using the Sobel edge finding method, 18 where the threshold was defined using Otsu's method. 19 The final edge image was dilated using a line structured element in 0° and 90° directions. Smaller holes of a predetermined size in the dilated edge image were filled.
Using connected component analysis, 20 the tissue area in the image was segmented. The percentage of stain for each image was computed using the corresponding complemented grayscale image. The minimum grayscale value for a pixel to be considered positively stained was determined from the lowest pixel value in the segmented positive control image. This value served as a threshold to determine the number of positive pixels in the segmented tissue area of any image. The percentage of stain was defined as the ratio of the number of positive pixels over the total number of pixels in the segmented tissue area. Percentage of stain of the negative control image were subtracted from all images and then normalized to the result from the positive control image.
Alpha-Gal inhibition ELISA test
To determine further the extent of α-Gal antigen removal in the porcine skin material treated by α-galactosidase, the inhibition ELISA assay was performed with a modified method that was based on the original Galili's method (Fig. 1). 21 In our method, tissue material was first chopped into small pieces and homogenized to tissue fragments about 5-100 μM. Tissue fragments, at the content from 0.5 to 200 mg/mL, were incubated overnight with M86 monoclonal antibody (1:50) at 4°C. After incubation, tissue fragments were removed by centrifugation (20,000 g). The binding interaction of M86 with α-Gal epitopes in tissue material was calculated by measuring the activity of free M86 left in the supernatant of samples. The ELISA method with α-Gal-BSA as a solid-phase antigen was used to measure the activity of the M86 antibody in the supernatant.
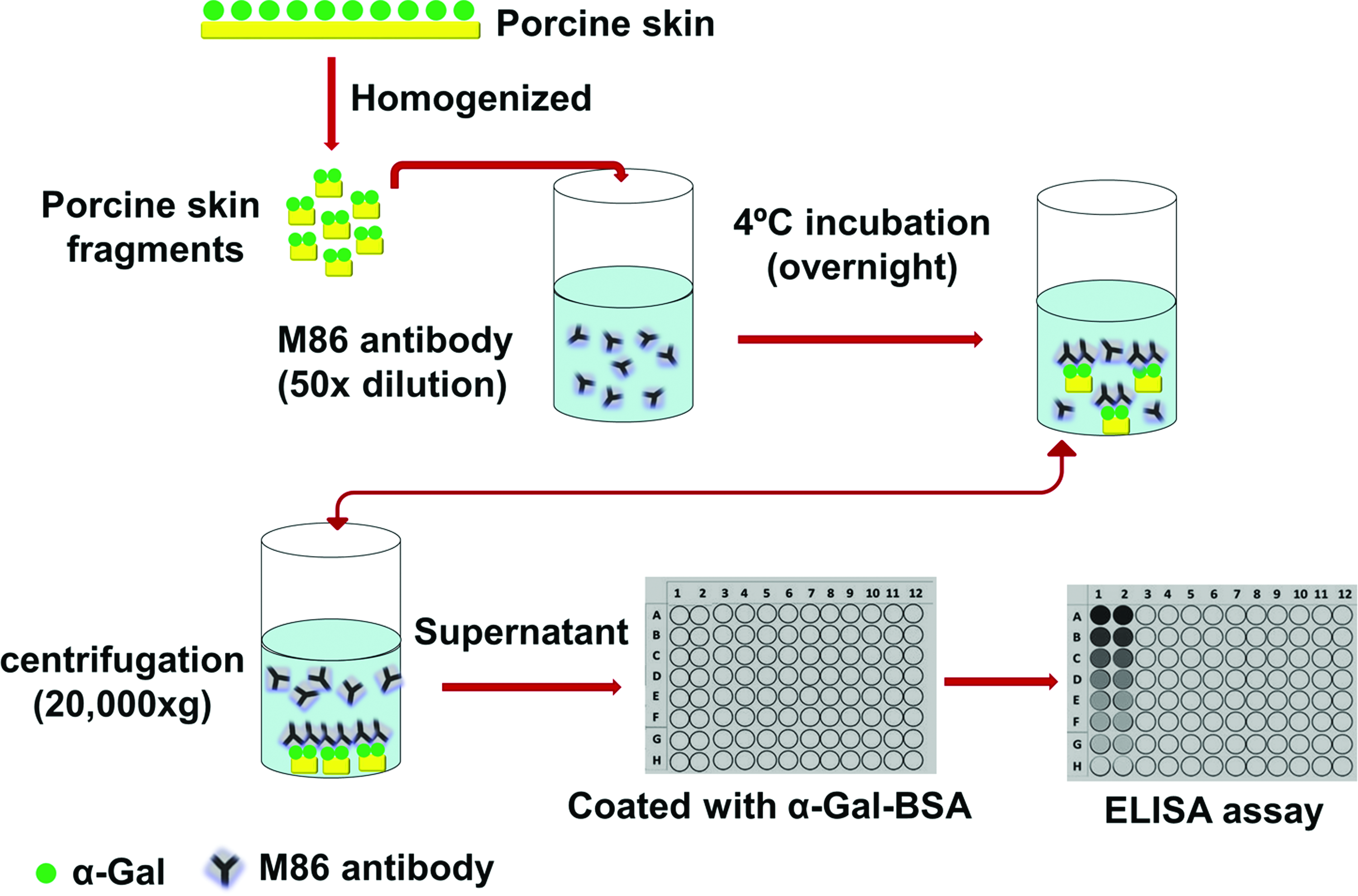
Schematic of the inhibition ELISA assay to measure the α-Gal epitope removal by α-galactosidase on porcine skin. The extent of the M86 antibody binding to the skin material is defined as a percentage inhibition of subsequent binding to α-Gal-bovine serum albumin (BSA). Color images available online at
Aliquots (100 μL) of α-Gal-BSA in PBS (10 μg/mL) was used to coat wells of microplates (Nunc) through an overnight incubation at 4°C. After three washes with PBS, the blocking procedure was performed using 250 μL per well of 2% BSA (Sigma-Aldrich) in PBS for 2 h at room temperature in darkness. Microplate wells were washed again three times with PBS solution. Aliquots of 100 μL sample supernatants were added into microwells and incubated for 1 h at 37°C in darkness. The secondary HRP-conjugated goat anti-mouse IgM antibody (diluted to 1:5000 in PBS) (Life technologies) was loaded for 30 min after washing. Finally, 100 μL TMB (Sigma-Aldrich) at 0.2 mg/mL was added to each well for 10 min incubation at 37°C in darkness and terminated with 50 μL of 2M sulfuric acid. The absorbance was measured by plate reader at 450 nm. The data were analyzed using GraphPad Prism 5.0.
Statistical analysis
Statistical comparison was carried out with the unpaired Student's t-test among various groups. The GraphPad Prism 5.0 was used for analysis. The difference is considered to be statistically significant when the p-value was less than 0.05.
Results
Efficacy of α-galactosidase cleavage of α-Gal epitopes in porcine skin
This study uses one single step of α-galactosidase incubation to remove the major xenoantigen (i.e., α-Gal epitope) in animal tissue. Figures 2 and 3 have presented the results from the immunohistochemical analysis and immunofluorostaining analysis. The enzymatic cleavage at a low concentration of 20 units/L is highly efficient at eliminating the α-Gal epitope in porcine skin, as detected by two immunohistochemical methods. Immunostaining with the M86 monoclonal antibody in the paraffin-embedded sections shows strong and extensive brown stain (positive α-Gal epitope expression) on the positive control porcine skin sample (incubated in PBS only). No staining was found on the α-galactosidase-treated porcine skin sample, the GalT-KO sample (the first negative control), and the decellularized human skin sample (the second negative control) (Fig. 2).
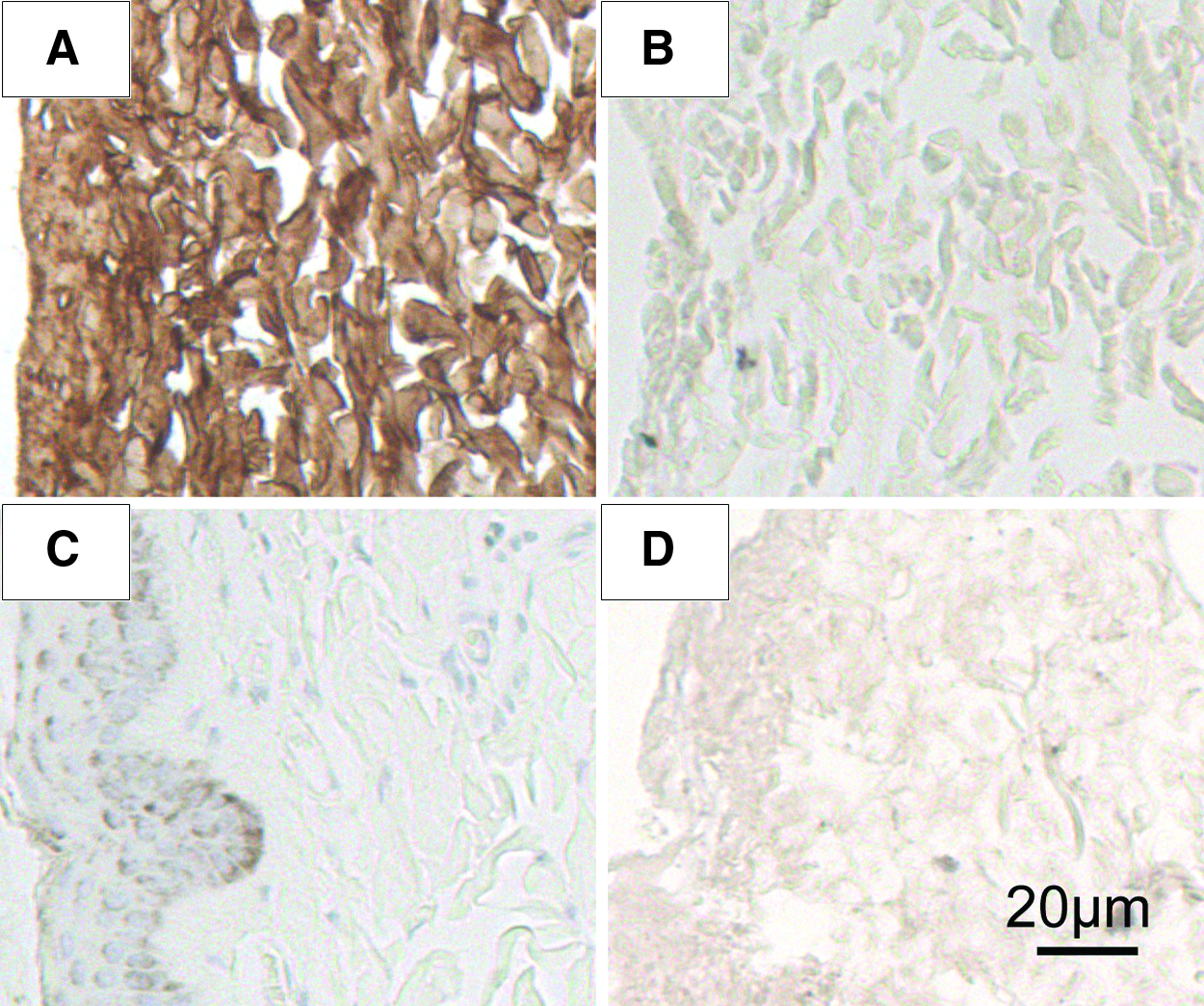
Representative M86 monoclonal antibody-stained immunohistochemical slides (magnification ×200).

Representative α-Gal expression on the skin as evaluated by immunofluorescence analysis. The sections are immunostained with FITC-labeled GS-IB4 and M86 monoclonal antibody in combination with FITC staining, respectively. Green fluorescence reveals the positive expression of α-Gal epitopes. Picture magnification is 200×. GalT-KO porcine skin and decellularized human skin are used as negative control materials. Color images available online at
Digital analysis of these immunostaining images indicates an average of 99.4% ± 0.6% reduction of antigen expression when it was scaled between the positive control porcine skin sample and the negative GalT-KO sample or an average of 104.3% ± 5.0% reduction when scaled between the positive control porcine skin sample and the negative decellularized human skin. The elimination of α-Gal epitope after enzymatic cleavage is further demonstrated by immunofluorostaining analysis with cryosections, using the FITC-labeled GSI-B4 lectin or M86 antibody followed by FITC-labeled anti-mouse IgM monoclonal antibody (Fig. 3). Specific green fluorescence is observed on the positive control porcine with both GS-IB4 lectin or M86 antibody. In contrast, there is no obvious immunofluorostaining in the enzyme-treated porcine skin sample or decellularized human skin sample, again indicating the complete elimination of α-Gal epitopes in the porcine skin.
In the efficacy study with the commercially available α-galactosidase from green coffee beans at 50 units/L for 24 h treatment, the presence of residual α-Gal epitopes was reduced to ∼10% for treated porcine skin at 0.5 mm thickness and <25% for skin samples at up to 4.0 mm thickness. The enzyme α-galactosidase from green coffee beans was less efficient than the recombinant α-galactosidase from B. fragilis, but still highly effective for the α-Gal antigen elimination in the dense porcine skin.
A proper method to quantify the antigen removal is required for process control and product quality evaluation. Traditionally, the extent of immunohistochemical staining is scored positive and negative, or ranked from 0 to 4. This approach, depending on the trained eyes of the pathologists, is less reproducible and severely handicapped when it is used for routine quality analysis at a commercial scale. However, the advance in automatic digital image analysis has rendered this classic method a new life. The efficacy study tested the effects of enzyme source, enzyme concentration, duration of enzyme incubation, and skin thickness on the α-Gal removal. Samples with different amounts of remaining α-Gal epitopes were used to assess the utility of immunohistochemical digital analysis as a quantitative quality control tool.
Figure 4 shows representative immunohistochemical images of enzyme-treated porcine skin samples along with the untreated porcine skin (positive control) and decellularized human skin (negative control). As described in the section of Materials and Methods, the slides of samples were digitized and the % of positive staining pixels for each image was computed by using the negative and positive control samples as reference points. The digital analysis of immunohistochemical slides was able to differentiate, within ±3–4% uncertainty, samples with GS-IB4 lectin stain in the range between 2% and 99%. This resolution is sufficient for the use as a routine quality analysis (say, to meet the threshold of positive staining ≤20%). The digital analysis is able to resolve a small difference among samples, which the eyes and mind of a pathologist cannot.

Representative immunohistochemical images of enzyme-treated porcine skin samples along with the untreated porcine skin (positive control, 100%) and decellularized human skin (negative control, 0%). The line curve shows the differentiation among 20 samples with different amounts of residual α-Gal epitopes in tissues. Horizontal bar in the image is 200 μm. Vertical bars in the line graph represent the uncertainty of digital analysis (c.a., standard deviation, ±3–4%). Color images available online at
Quantitative M86 inhibition ELISA assay
The mouse M86 monoclonal antibody is highly specific for α-Gal epitopes and binds to synthetic α-Gal epitopes linked to bovine serum albumin (α-Gal-BSA). In the present study, tissue homogenates at various contents were incubated with a solution containing the M86 antibody. The α-Gal epitopes in tissue fragments immobilize the M86 antibody before the ELISA assay, resulting in an inhibition in the subsequent M86 binding to the substrate α-Gal-BSA. The extent of M86 binding to the tissue fragments, a measure of α-Gal epitope expression, is proportionally related to the percent inhibition of subsequent M86 binding to the substrate α-Gal-BSA. In other words, the ELISA inhibition values correspond to the expression of α-Gal epitopes in tissue samples.
Therefore, the M86 antibody assay should be a promising method for quality control application in the production and selection of animal tissue-based bioprosthetic devices. A comparison was made among tissue homogenates of the positive control porcine skin, α-galactosidase-treated porcine skin (from B. fragilis), and decellularized human dermis (Fig. 5). The porcine skin homogenate (positive control) readily interacts with the M86 antibody and registers the ELISA inhibition value about 92.8% ± 11.3% (n = 9) at a tissue content of 150 mg/mL. The inhibition of α-galactosidase-treated porcine skin is about 38.5% ± 12.6% (n = 5), as compared with 27.6% ± 12.1% (n = 6) background inhibition of decellularized human skin dermis (the negative control) at the same tissue content. This result indicates a 83.3% elimination of α-Gal epitopes: (92.8–38.5)/(92.8–27.6) = 83.3%. Using the logarithmic response model, the rate constant of the M86 binding kinetics (Fig. 5) was calculated to be 15.0.5 ± 4.3 (n = 10), 6.1 ± 2.6 (n = 7), and 4.4 ± 1.8 (n = 9) in the porcine skin homogenate (positive control), the α-galactosidase-treated porcine skin homogenate, and the decellularized human skin dermis (the negative control), respectively. The latter calculation method also demonstrates ∼84% elimination of α-Gal epitopes in porcine skin after incubation in 20 units/L α-galactosidase solution for 20 h [(15.0–6.1)/(15.0–4.4) = 84.0%].

ELISA inhibition assay performed with tissue homogenates of untreated porcine skin, α-galactosidase-treated porcine acellular skin and decellularized human dermis, using the monoclonal anti-Gal antibody M86. Data are presented as % inhibition of M86 binding to α-Gal-BSA in ELISA, after overnight incubation of M86 with skin homogenate at various contents. Individual data point is the mean ± standard deviation (n = 10, 6, and 9 for untreated porcine skin, α-galactosidase-treated porcine acellular skin, and decellularized human dermis, respectively).
However, the standard deviation of the inhibition values is very large, as shown in Figure 5. Figure 6 shows the M86 binding inhibition as a function of normalized tissue content used in the ELISA inhibition assay, where the tissue contents of α-galactosidase-treated porcine skin samples were normalized, based on the average inhibition values of individual samples over repeated measurement experiments, to the tissue contents equivalent to the positive control (untreated) skin samples with the same amount of α-Gal epitopes. For example, the normalized tissue content of an enzyme-treated sample with an inhibition value of 50% is calculated to be: the actual tissue amount used in the assay × 50%. All data points from 10 experiment series for untreated porcine skin and 6 experiment series for α-galactosidase-treated porcine dermis were superimposed onto the same scatter plot.
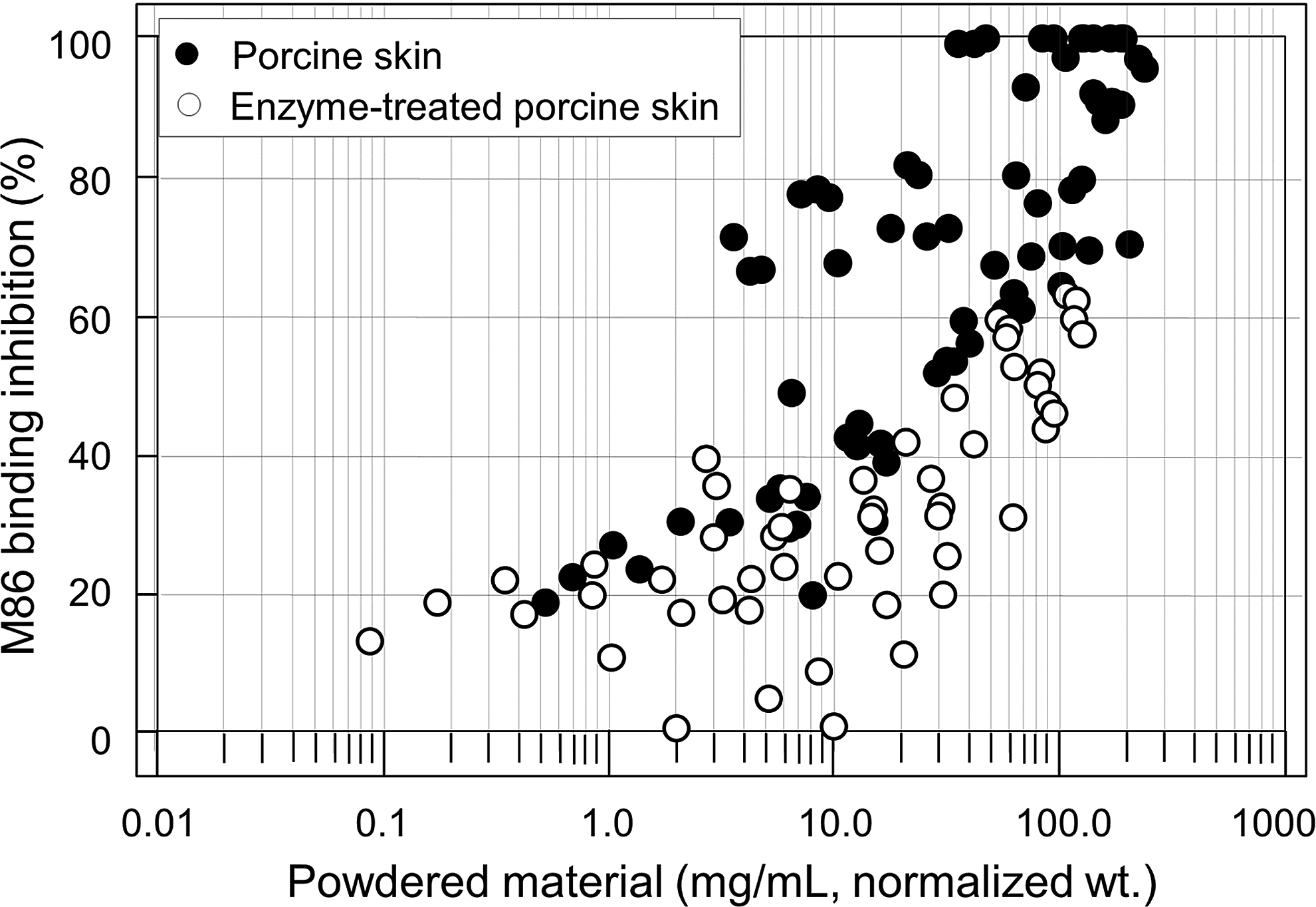
M86 binding inhibition as a function of normalized tissue content used in the ELISA inhibition assay. The tissue contents of individual α-galactosidase-treated porcine skin samples were normalized to the values equivalent to the positive control skin samples with the same amount of α-Gal epitopes. The plot shows a general trend of inhibition with the increasing tissue content, but the large extent of data point scattering makes the method less useful as a quality control tool.
Such a plot shows the reproducibility and repeatability of the M86 inhibition assay. The scatter plot demonstrates a clear quantitative trend in tissue content between 0.5 and 200 mg/mL with very large variation, suggesting that the M86 method needs further optimization when used for quantitative quality analysis of solid tissue fragments. As the assay is in the current form, statistically it cannot be used to resolve a difference of <20% between samples with five repeated experiment series.
Discussion
The present study has demonstrated that enzymatic cleavage at low concentration is extremely efficient at eliminating the major xenoantigen α-Gal epitope even in a tissue as dense as the porcine dermis (Figs. 2–4). The enzyme α-galactosidase is commercially available in several forms, and some of them are used as a drug to treat the Fabry disease (a deficiency of α-galactosidase A in plasma and leukocytes of affected males). The economics of the tissue treatment by α-galactosidase before clinical uses of animal tissues or organs is calculated as follows: The Sigma-Aldrich's retail price for α-galactosidase from green coffee bean is US$419 for 50 units, which can be used to eliminate the α-Gal epitopes in ∼300 g porcine skin tissue if the enzyme is used at the concentration of 50 units/L, a concentration similar to the one used in the present study.
Assuming that the porcine skin is harvested at an average of 0.5 mm thickness for burn treatment, the enzyme material costs less than US$7 per 100 square centimeters (10 × 10 cm). For other 3-dimensional tissues or organs, the α-galactosidase treatment costs less than US$1.4 per gram tissue, a price that is inexpensive and really affordable. Porcine skin was chosen because of its high tissue density and low porosity, with the perception that it would be even easier for other animal soft tissues and highly vascular organs. Decellularized tissue matrices are currently used in various surgeries for tissue repairs, such as hernia, breast argumentation and reconstruction, as well as abdominal wall reconstruction. The α-Gal epitope is also present in the extracellular matrix of animal tissues. It is conceivable that treatment by α-galactosidase would be much more efficient with decellularized tissue matrices, because decellularized tissue matrices are looser than untreated animal tissues.
In addition, the biology of the α-galactosidase is well known, and its production can be scaled up with ease. Therefore, the α-galactosidase process is affordable for commercial manufacturing of bioprosthetic devices from animal tissues. The recombinant form of α-galactosidase from B. fragilis that was developed in our laboratory is even more efficient at removing the α-Gal epitopes and can be produced at a lower cost (Figs. 2 and 3).
Porcine skin overlay is an effective way to treat extensive burn wounds in early stages and reduces scar formation during healing. 22 However, changes have been observed in the human body to the anti-α-Gal antibody titers, indicating that α-Gal antigen immune risk potential cannot be ignored.23,24 A number of attempts have been made to reduce or inactivate the antigen from tissues to ensure the safety of animal-derived medical products, but only a few have resulted in a practical clinical use. For example, glutaraldehyde-fixed porcine skin or acellular dermal matrix (ADM) has been utilized as a temporary cover for third-degree burns. However, glutaraldehyde treatment does not eliminate α-Gal immunogenicity. It was reported that α-galactosidase treatment of porcine valve can reduce the α-Gal epitope and the immune response. 25
Furthermore, another study indicated that no or minimal anti-Gal activity was detected in the primates implanted with Gal−/− and α-galactosidase-treated porcine ADM. 14 Our data confirmed that α-Gal epitope was greatly reduced when porcine skin was treated with α-galactosidase. The enzymatic removal of the α-Gal epitopes in porcine skin is feasible and commercially viable for clinical use.
The assessment of α-Gal expression on various cells, tissues, and organs is important for the prediction of anti-Gal-mediated immune rejection of xenografts. In the study, we have examined the suitability of two detection methods for the quality control application. Alpha-Gal epitopes on isolated cells can be detected by flow cytometry analysis and immunohistochemistry. However, for the tissue and organs, only qualitative immunohistochemistry and immunofluorescence exist. The ELISA inhibition assay developed originally by Galili's measures the number of α-Gal epitopes on cells with reference to a standard α-Gal source (i.e., mouse myeloma SP2/0 cells or rabbit red blood cells). 21
The ELISA assay should be also useful in assessing the removal efficiency of xenoantigens by chemical shielding, genetic engineering, or enzyme treatment. Because this method uses mouse myeloma SP2/0 cells or rabbit red blood cells as a reference, it can only provide a semiquantitative estimate of α-Gal epitope expression. We have adopted the method and modified the assay for solid tissue materials. Solid tissue material is first homogenized to particulates at 5–100 μm. A homogenate sample with α-Gal epitopes would remove the M86 antibody before the ELISA assay, reducing subsequent M86 binding to the substrate α-Gal-BSA (inhibition). The direct relationship between the amount of α-Gal epitopes in the homogenate sample and reduction of subsequent M86 binding to the substrate α-Gal-BSA was explored for the determination of α-Gal epitope elimination rate by α-galactosidase (i.e., the % subsequent binding to α-Gal-BSA or 100% inhibition).
Theoretically, the ELISA inhibition of untreated porcine should reach 100% and decellularized human skin should have no inhibitory activity to the M86 antibody. But our data showed some inhibitory activity with decellularized human skin homogenate (Fig. 5), demonstrating certain nonspecific reaction with the M86 antibody. The M86 antibody was considered to be the most suitable probe to identify and quantify the residual α-Gal epitopes in bioprosthetic devices, 9 according to earlier data tested with cells, where the α-Gal epitopes exist on the cell surface. 1 We have examined the reproducibility and repeatability of this method for the solid tissue material such as micronized porcine dermis particles in the present study, and observed a very large variation in testing tissue homogenates (Figs. 5 and 6).
A statistical calculation based on the data of the present study indicates that the reproducibility and repeatability of this M86 binding method is low, and cannot resolve a difference of <20% between samples with five repeated experiment series. The high cost of test material and labor, as well as the low throughput render the method less valuable for quality control applications as it is in the current form. It was possible that the expression of α-Gal epitopes in solid tissue fragments were affected by the accessibility due to the limited diffusibility of M86 antibodies into the tissue particles and/or the size uniformity of tissue fragments. The range of tissue content was shown to be 1–200 mg/mL for the ELISA inhibition assay as illustrated in Figure 6. The ELISA inhibition method may be useful, but remains to be optimized further as a quantitative assay for solid tissue materials. Digital analysis of immunostaining images was also evaluated in an attempt to establish a more convenient method for the detection of α-Gal epitope removal from solid tissues (Fig. 4). The image analysis method is normalized with the positive control (e.g., untreated animal tissues) as 100% and the negative control (e.g., human tissues) as 0%. The image analysis method is straightforward and can be used for full-range detection.
In summary, the study has demonstrated the efficacy and affordability of α-galactosidase treatment to remove α-Gal xenoantigen in porcine skin. The enzymatic cleavage of α-galactosidase can be integrated into a typical animal tissue process with ease. The results also show that both the traditional immunohistochemistry and the inhibition ELISA with further optimization can be used as reliable quality control methods for achieving a greater safety of animal tissue-based grafts.
Footnotes
Acknowledgments
The work was supported by the National Science and Technology Major Project of the Ministry of Science and Technology of China (No.2013ZX09J13110). The authors thank Dr. Hussain Fatakdawala for assisting the digital analysis of immunohistochemical images.
Disclosure Statement
No competing financial interests exist.