
Abstract
The establishment of adipose tissue test systems is still a major challenge in the investigation of cellular and molecular interactions responsible for the pathogenesis of inflammatory diseases involving adipose tissue. Mature adipocytes are mainly involved in these pathologies, but rarely used in vitro, due to the lack of an appropriate culture medium which inhibits dedifferentiation and maintains adipocyte functionality. In our study, we showed that Dulbecco's Modified Eagle's Medium/Ham's F-12 with 10% fetal calf serum (FCS) reported for the culture of mature adipocytes favors dedifferentiation, which was accompanied by a high glycerol release, a decreasing release of leptin, and a low expression of the adipocyte marker perilipin A, but high expression of CD73 after 21 days. Optimized media containing FCS, biotin, pantothenate, insulin, and dexamethasone decelerated the dedifferentiation process. These cells showed a lower lipolysis rate, a high level of leptin release, as well as a high expression of perilipin A. CD73-positive dedifferentiated fat cells were only found in low quantity. In this work, we showed that mature adipocytes when cultured under optimized conditions could be highly valuable for adipose tissue engineering in vitro.
Introduction
B
Several substances are known to reduce or inhibit lipolysis and favor lipogenesis.17–23 Insulin, as an anabolic hormone, is known to play a multipotent role in glucose and lipid metabolism, where it is mainly responsible for the uptake and storage of glucose and fatty acids. 24 Dexamethasone activates transcription factors upregulating enzymes that promote lipid uptake and adipogenesis. 25 Biotin, also known as vitamin H, serves as a prosthetic group for carboxylases, which are involved in gluconeogenesis and fatty acid biosynthesis.26,27 Pantothenic acid, also known as Vitamin B5, is important for the synthesis of coenzyme A, which is needed for the synthesis of fatty acids.28,29 Fetal calf serum (FCS) has contrary effects on mature adipocytes, depending on the used concentration. On the one hand, serum contains some insulin-like factors that have comparable effects on glucose as insulin does. 17 On the other hand, a high amount of serum favors the dedifferentiation of mature adipocytes because it also contains several factors inhibiting lipogenesis, like tumor necrosis factor-alpha, growth hormones, and epidermal growth factor. 23
In this study, we evaluated two commercially available media concerning their ability to maintain the morphology and functionality of mature adipocytes for up to 3 weeks.
Materials and Methods
Human tissue samples
All research was carried out in accordance with the rules for investigation of human subjects as defined in the Declaration of Helsinki. Patients gave a written agreement according to the permission of the Landesärztekammer Baden-Württemberg (F-2012-078; for normal skin from elective surgeries).
Adipocyte isolation and measurement of cell size
Mature adipocytes were isolated from human fatty tissue of plastic surgeries received from Dr. Ziegler (Klinik Charlottenhaus). The isolation was based on the procedure previously described by Zhang et al. 30 Briefly, the connective tissue was removed and adipocytes were isolated by collagenase digestion (125 U mL−1 in Dulbecco's Modified Eagle's Medium [DMEM; Biochrom] containing 1% bovine serum albumin [BSA; Sigma-Aldrich]) for 1.5 h at 37°C under agitation. The cell suspension was filtered through a 500-μm meshed sieve, centrifuged (19 g, 1 min), and washed twice.
The mean size of mature adipocytes and DFAT was determined of at least 60 cells from three different donors, whereby the inner diameter of the adipocytes and the longitudinal axis of the DFAT were evaluated.
Encapsulation of mature adipocytes into collagen type I hydrogels
Mature adipocytes were seeded in a collagen type I hydrogel (9 mg mL−1 from rat tail; Fraunhofer IGB) in 24-well plate inserts (Brand). A neutralization buffer consisting of 10 × DMEM/Ham's F-12 (Biochrom) and 50 mM NaOH in Aqua dest (1:1) with 0.2 M NaHCO3 and 0.225 M HEPES (Roth) was prepared. Four portions of collagen gel were mixed with four portions of packed mature adipocytes and one portion of neutralization buffer. Three hundred microliters of the mixture was pipetted into each well and gelled for 10–20 min at 37°C.
Cells were then cultured for 3 weeks in three different media compositions: Medium 1 consisted of DMEM/Ham's F-12 and 10% FCS as a control medium. As medium 2, AM-1 (ZenBio) was used, consisting of DMEM/Ham's F-12 medium, HEPES pH 7.4, fetal bovine serum, biotin, pantothenate, human insulin, and dexamethasone. Medium 3 was the Adipocyte Nutrition Medium (PromoCell) supplemented with 0.03 mL mL−1 FCS (Gibco), 8 μg mL−1 d-biotin, 0.5 μg mL−1 human insulin, and 400 ng mL−1 dexamethasone (all purchased from Sigma-Aldrich).
Immunofluorescence staining
Hydrogels were fixed using Bouin's fixation (Sigma-Aldrich) for 1 h and watered with tap water for several hours. Gels were embedded into paraffin and slices of 5 μm were generated. Deparaffinization was done according to a standard protocol. Sections were heat demasked with a target retrieval solution, pH 9 (Dako), for 20 min in a preheated steamer (Braun). Cells were blocked with 3% BSA in PBS+ for 30 min. Primary antibody (perilipin A: 1:200; Sigma-Aldrich, CD73: 1:25; Santa Cruz) was diluted with an antibody diluent (Dako) and incubated overnight at 4°C. Tissues were then stained using the EnVision+ System-HRP (Dako) according to the manufacturer's protocol.
Live cell staining
Cytoplasm/lipid droplet/nuclei staining
Nonfixed hydrogels were stained with 1 μg mL−1 Hoechst 33342 in PBS+ for 15 min at 37°C. Thirty μL mL−1 Nile Red (Lonza) and 10 μg mL−1 fluorescein diacetate (FDA; Sigma-Aldrich) were added and incubated for another 15 min at 37°C. Images were taken in PBS+ using a laser scanning microscope (Zeiss).
Cytoplasm/nuclei staining
Cells were stained with FDA and Hoechst 33342 as described above. The total cell number was determined as the number of nuclei from 5 × magnified 100 μm z-Stack images using the Cell Profiler Cell Images Analysis Software (Broad Institute, United Kingdom). The ratio of univacuolar and multivacuolar cells and DFAT was analyzed manually. For every analysis day, at least three representative images of three hydrogels were analyzed per medium composition.
Functionality tests
Leptin enzyme-linked immunosorbent assay
An enzyme-linked immunosorbent assay (Pepro Tech) was performed according to the manufacturer's protocol. For color development, 100 μL tetramethylbenzidine (TMB) substrate (Sigma-Aldrich) for each well was used. Incubation took place at room temperature (RT) for ∼15 min. The enzymatic reaction was stopped with 50 μL 1 M sulfuric acid, and the color development was measured at 450 nm with a wavelength correction set at 570 nm (Tecan).
Glycerol release
Lipolysis was detected with a glycerol kit (Randox). Briefly, 5 μL samples were mixed with 100 μL reagent and incubated for 10 min at RT. The color development was analyzed at 520 nm.
Leptin and glycerol measurements were performed 24 h after the last media exchange. Values were normalized to the number of univacuolar and multivacuolar adipocytes, named as fat cells.
Statistics
All experiments were repeatedly performed, successively using cells from at least three different donors. Data were compared by a one-way analysis of variance with a Tukey post hoc test using Origin Pro 8.5 G. Statistic significances were stated as p ≤ 0.05.
Results
Mature adipocytes dedifferentiated during in vitro culture
Mature adipocytes were encapsulated in a collagen type I hydrogel and cells cultured in medium 1 dedifferentiated over time (Fig. 1). After encapsulation, they showed a round morphology with one huge lipid droplet filling the whole cytoplasm. The nucleus was compressed to the cell membrane. Over the culture period, cells were elongating and stored their lipids in several lipid droplets. The nucleus migrated to the center of the cell. When reaching a fibroblast-like morphology, these DFAT did not store any lipid vacuoles in their cytoplasm and started to proliferate. Mature adipocytes had a diameter of 79 ± 19.0 μm, whereas DFAT had a nonsignificantly smaller cell length with 70 ± 24.4 μm (Fig. 1D).

Mature adipocytes lose their lipid droplets during dedifferentiation. Cells were cultured in medium 1. Lipid vacuole was stained with Nile Red and is shown in red, nuclei were stained with Hoechst 33342 in blue, and cytoplasm was marked with fluorescein diacetate (FDA) in green.
Optimized media reduced the dedifferentiation of mature adipocytes
The total number of cells encapsulated in a collagen type I hydrogel was evaluated and shown in Figure 2 as a percentage of the control medium (medium 1) on day 1. A similar cell amount (112% ± 43.6%) could be found in medium 2, whereas in medium 3, significantly fewer cells were available (24% ± 4.4%). Only a slight decrease in the cell number could be seen in medium 1 and 2 on day 7, whereas the number of cells in medium 3 stayed constant up to day 21. In medium 1 and 2, cells proliferated on day 14, to a level of 125% ± 43.1% and 152% ± 23.8%, respectively. The cell amount further increased on day 21 to 133% ± 65% in medium 1 and 173% ± 62.7% in medium 2.
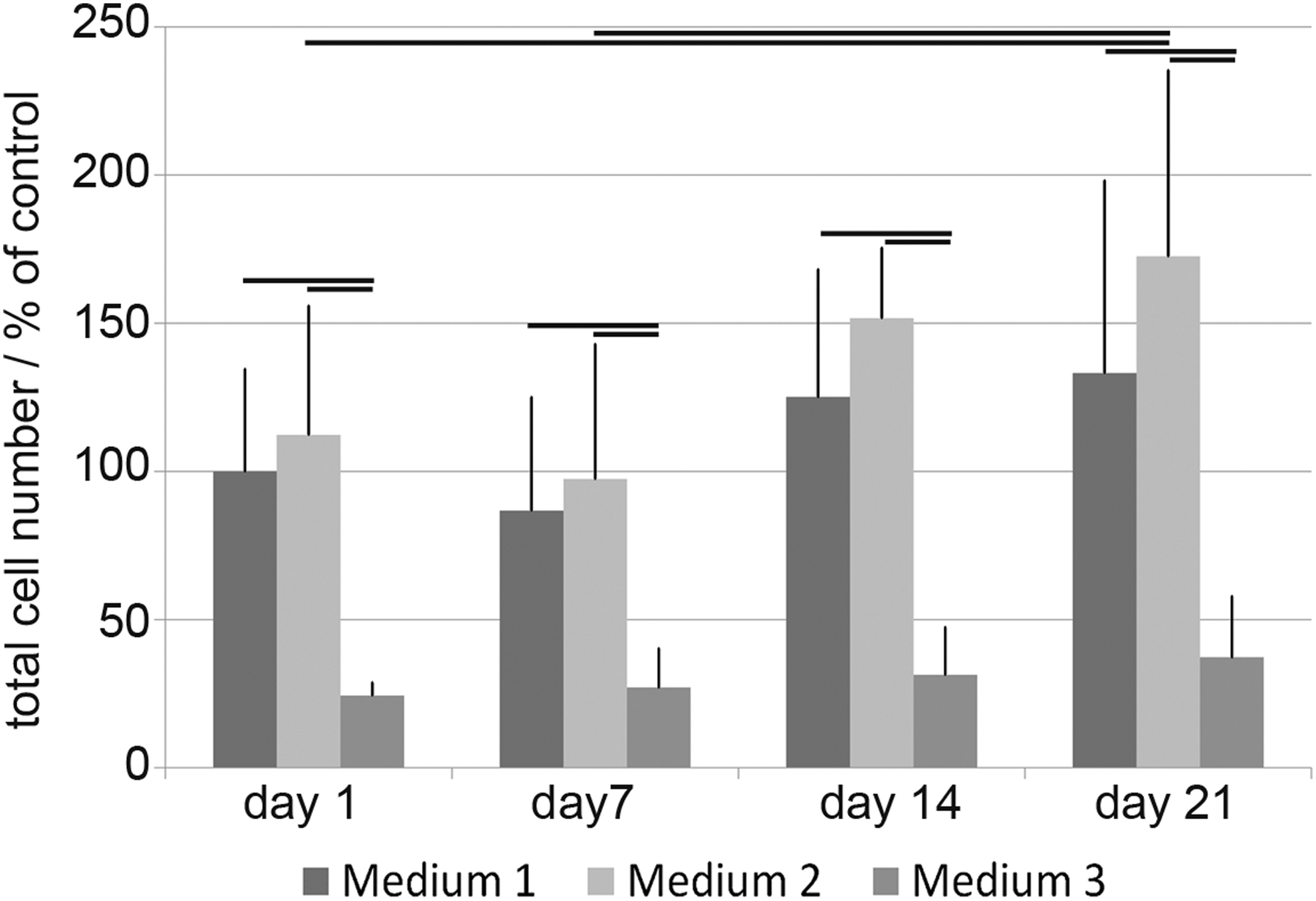
Total cell number per hydrogel. To evaluate the total cell number, the amount of nuclei was counted from 5× magnified 100 μm z-Stack images and given as a percentage of the control medium 1 on day 1 (p ≤ 0.05).
Figure 3A shows the percentile number of univacuolar and multivacuolar adipocytes and DFAT when culturing mature adipocytes for 21 days in three different media. On day 1, only mature adipocytes could be found (Fig. 3B–D). The number of mature adipocytes on day 7 in medium 1 with 62.3% ± 25.37% was significantly lower when compared with medium 3 with 85.7% ± 10.29%. In medium 2, about 83.2% ± 13.36% mature adipocytes could be found. The number of multivacuolar adipocytes increased only slightly in all three media. The number of DFAT was significantly higher in medium 1 (37.2% ±24.94%) compared with medium 2 (12.1% ± 11.58%) and medium 3 (13.3% ± 10.18%).

Evaluation of mature adipocytes cultured in different media for up to 21 days.
On day 14, only 49.0% ± 28.89% of the cells were mature adipocytes in medium 1, whereas medium 2 and 3 contained 72.8% ± 19.63% and 72.8% ± 17.01% mature adipocytes, respectively. The number of multivacuolar adipocytes was significantly higher in medium 2 (23.1% ± 17.85%) compared with medium 1 and 3. DFAT increased significantly on day 14 in medium 1 compared with medium 2 and 3: 48.4% ± 26.58% DFAT could be found in medium 1, 4.1% ± 3.22% were available in medium 2, and 20.8% ± 15.89% have been found in medium 3.
On day 21, the highest amount of mature adipocytes could be found in medium 3 with 71.5% ± 21.14%, followed by medium 2 with 45.6% ± 28.27% and medium 1 with 39.9% ± 27.51%. The number of multivacuolar adipocytes was highest in medium 2 with 47% ± 33.47%, but very few could be found in medium 1 and 3. The significantly highest amount of DFAT was seen in medium 1 on day 21 with 58.9% ± 27.64%, whereas in medium 2, only 7.4% ± 9.88% DFAT could be found. Medium 3 had 20.8% ± 16.94% DFAT. This is visualized in Figure 3B–D.
Specific marker expression is shown in Figure 4. On day 1, perilipin A was expressed from adipocytes cultured in all media, where there was a lower expression in medium 3. Only slight CD73 staining was found on day 1. On day 21, cells expressed perilipin A to a lower extent than on day 1. Perilipin A could also be found in multivacuolar fat cells. A high amount of CD73-positive cells was detected in medium 1 on day 21, whereas the level was lower in medium 2 and 3.
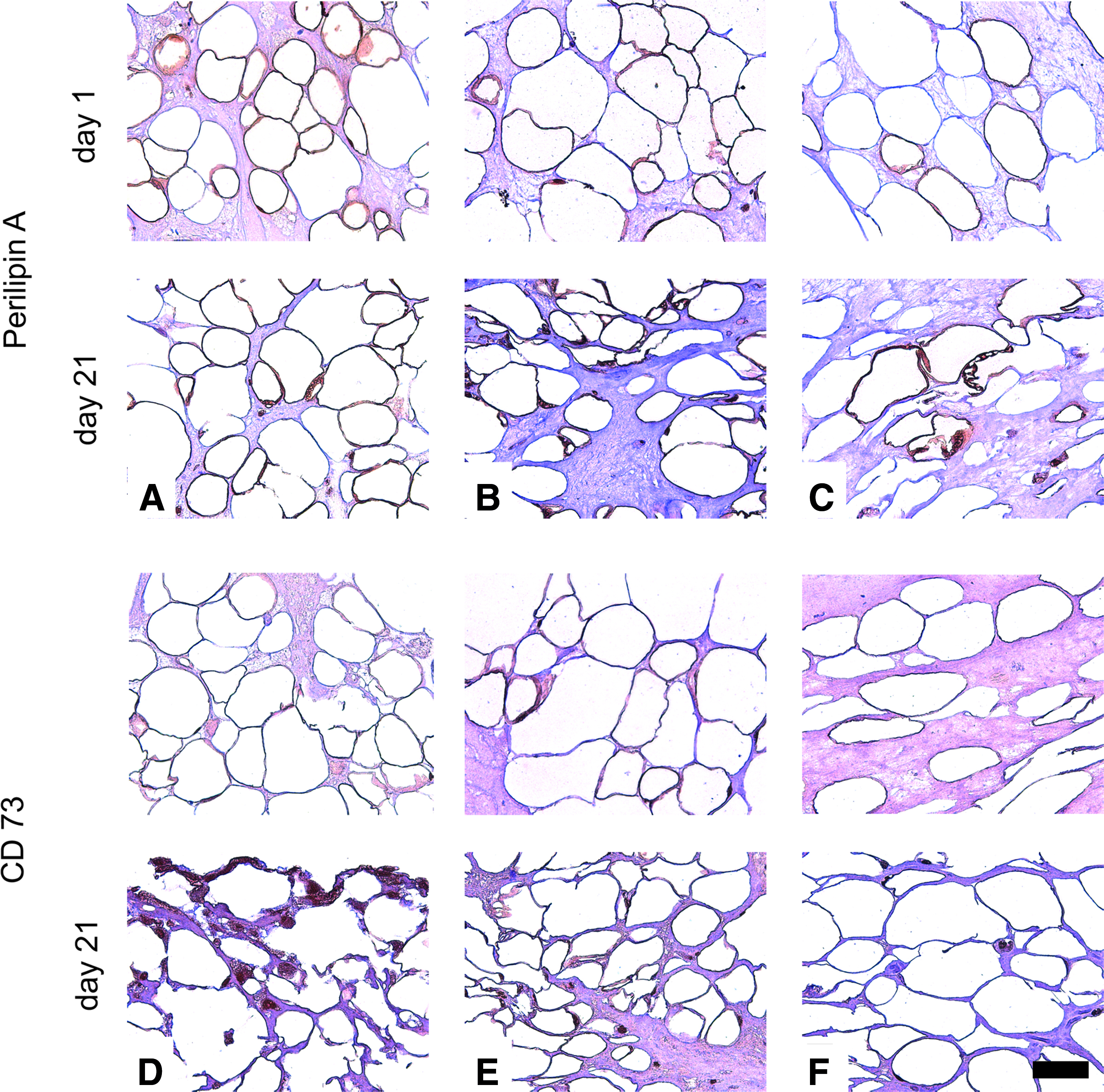
Perilipin A and CD73 expression of cells in different media.
Mature adipocytes remained functional when cultured in an optimized media
The lipolytic activity of cells cultured in different media was analyzed by measuring the glycerol release and is shown in Figure 5. The values were normalized to 10,000 fat cells, whereas univacuolar as well as multivacuolar cells were considered since both cell types are able to undergo lipolysis.

Lipolytic activity of cells cultured in different media by measurement of the glycerol release. The values were normalized to 10,000 fat cells, where univacuolar and multivacuolar cells were considered to undergo lipolysis (p ≤ 0.05).
On day 1, only a slight release of glycerol occurred in all tested media, whereas the amount of glycerol in medium 3 was significantly higher than in medium 1 and 2. On day 7, a significantly higher amount of glycerol was measured from cells cultured in medium 1 with 57 ± 19.4 nmol. In medium 2, 13 ± 6.4 nmol glycerol could be measured in the supernatants. A higher amount of glycerol of 27 ± 14.6 nmol could also be found in medium 3 on day 7. On day 14, the glycerol release was reduced again. Glycerol release decreased further on day 21 to 14 ± 5.0, 8 ± 5.4, and 19 ± 11.7 nmol in all tested media, respectively.
Cell functionality was analyzed by measuring the leptin release, which is seen in Figure 6; again, the data were normalized to 10,000 fat cells. On day 7 to 21, a significant higher release of leptin was found in medium 2 and 3 compared to medium 1. A similar release of leptin was seen in medium 1, 2, and 3 on day 1 with 0.7 ± 0.30, 0.3 ± 0.31, and 0.2 ± 0.13 ng leptin, respectively. In medium 1, the leptin release stayed constantly on this low level. Cells cultured in medium 2 released 3.3 ± 2.08 ng leptin on day 7, which only decreased slightly up to day 21. In medium 3, 4.0 ± 2.0 ng leptin was released on day 7 and on day 14 and 21, a slightly lower amount with 3.2 ± 1.48 and 2.8 ± 1.47 ng leptin, respectively, were measured.

Leptin release from cells cultured in different media for up to 21 days. The values were normalized to 10,000 fat cells, where univacuolar and multivacuolar cells were considered to produce leptin (p ≤ 0.05).
Discussion
Mature adipocytes are involved in various diseases, such as obesity or cancer. To date, mainly differentiated preadipocytes have been used for investigations in adipocyte-related cross talks, since mature adipocytes dedifferentiate under in vitro conditions. In this study, two media compositions were evaluated in comparison to a standard medium for their suitability to culture mature adipocytes.
Mature adipocytes dedifferentiated during in vitro culture
It was previously demonstrated that mature adipocytes dedifferentiate in vitro.10,14,31,32 DFAT are then able to proliferate.33,34 Adipocytes are thought to be very plastic cells and depending on the nutrition status, they can store or release triglycerides, which has a drastic impact on their cell size. 35 It is thought that by means of lipid droplet fragmentation, and therefore, surface enlargement, lipases have a facilitated contact to triglycerides, which gives a direct link to lipolysis.36,37
Optimized media reduced the dedifferentiation of mature adipocytes
Triggers for the dedifferentiation of mature adipocytes are still mainly unknown, and only a few hypotheses are available. Some authors suggested the susceptibility of adipocytes against hypoxia.23,38 Others proposed that a high cell density is important to decelerate the dedifferentiation process. 39 Therefore, we used the maximal number of fat cells possible to incorporate into a stable collagen type I hydrogel. It is also believed that the culture of human fat cells in the absence of stimulatory factors results in spontaneous basal lipolysis. 40 We were now able to show that the morphological and functional stability of mature adipocytes could be largely maintained by using an optimized media in vitro.
The total cell number was increasing slightly in medium 1 and 2 over the culture period. Since mature adipocytes are in a terminal differentiation status, it is likely that in medium 1, DFAT started to proliferate and therefore increased the cell number. In medium 2, only a small amount of DFAT were available. We propose that multivacuolar adipocytes were able to divide, which was already shown by Sugihara et al. 10 In medium 3, there was an initial drastic decrease in the cell number, which then stayed stable over the culture period. This led us to the assumption that for the initial cell survival, either a high FCS content (medium 1) or the addition of pantothenate, the supplement that distinguished medium 2 and 3, might be indispensable. Further research is needed to elucidate the roles of FCS and pantothenate in initial mature adipocyte cell survival.
We have seen that cells cultured in medium 3 had the highest percentage of mature adipocytes on day 21. In medium 1, mature adipocytes were mainly replaced by DFAT, which could be confirmed by immunofluorescence staining of CD73. In medium 2, however, only a small amount of DFAT could be detected, and most of the cells were multivacuolar. The decelerated dedifferentiation in medium 2 and 3 is thought to rely on the content of FCS, biotin, insulin, dexamethasone, and pantothenate (only in medium 2).
Justesen et al. showed that mature adipocytes were dedifferentiating in the presence of 10% FCS, which could be confirmed by us in medium 1. 41 It is thought that FCS contains many insulin-like factors, and Gliemann proposed that 2% serum has the same effect as insulin on the intake and metabolism of 0.1 mg mL−1 glucose in mature adipocytes. 17 This supports our results that a high FCS content of 10% in medium 1 favored dedifferentiation, whereas a low content in medium 3 promoted the maintenance and viability of mature adipocytes. We also suggest that medium 2 has a low FCS content. Other groups have performed their studies with a low serum content between 1% and 3% FCS, but did not evaluate the long-term culture of adipocytes.42–45 Sugihara et al. reported that 20% FCS showed the best adhesion and proliferation of DFAT. 8
Several researchers could show that insulin had an antilipolytic effect on mature adipocytes in short-term culture in vitro, where they defined a wide range of concentrations as optimal.17–19,46,47 Generally, the glycerol release in medium 2 and 3 was lower compared with medium 1 on all analyzed days (except medium 3 on day 21), which led us to the assumption that insulin had a positive effect on the lipolytic activity of adipocytes.
An enhanced effect could be seen by the combination of supplements. It was shown that the combination of insulin and dexamethasone resulted in an increased incorporation of glucose into lipids compared with insulin alone. 20 Also, medium 2 and 3 in our study contained insulin and dexamethasone, which suggests a synergistic effect.
The effect of biotin on mature adipocyte behavior has not yet been examined intensively, but was used in several studies for the differentiation of preadipocytes into adipocytes. Kuri-Harcuch et al. have shown that 3T3 adipocytes did not accumulate triglycerides without biotin during differentiation. 21 Others showed that lipid accumulation was increased using biotin-supplemented media.22,48,49 Janke et al. cultured mature adipocytes for a short term while adding 4 μg mL−1 biotin to the culture medium. 50 Medium 2 and 3 were also supplemented with biotin, of which we used 8 μg mL−1 in medium 3, but the concentration for medium 2 was unknown. According to these data, a positive effect of biotin on the stability of mature adipocytes was seen.
The effect of pantothenate and dexamethasone on mature adipocytes is hardly known. Many investigations have been performed with a coapplication of the two supplements. Wang et al. reported a synergistic effect of dexamethasone with insulin on glucose incorporation and lipogenesis. 20 Furthermore, insulin and dexamethasone were responsible for an increased leptin release, but were not thought to be the main modulators. 51 Also, biotin combined with panthothenate improved lipogenesis. 23
Mature adipocytes remained functional when cultured in an optimized media
To evaluate cell functionality, the release of glycerol and leptin was measured. We observed that cells cultured in medium 1 had a high release of glycerol and expressed leptin only in very small amounts. Cells cultured in medium 2 produced lower amounts of glycerol, but higher amounts of leptin at all analyzed days compared with medium 1. The glycerol release of cells in medium 3 stayed on an average level from day 7 to 21, whereas the leptin production was higher compared with medium 1 and 2.
Glycerol is thought to be a product of lipolysis.39,52 Therefore, it is likely that the number of mature adipocytes in medium 1 decreased because of lipolytic activities, which was shown in a high release of glycerol from these cells and appearance of a fibroblast-like morphology. We could see that dedifferentiation also occurred in cells cultured in medium 2, but mainly to the level of multivacuolar adipocytes. The trigger to hold these cells in a multivacuolar state instead of completing the dedifferentiation process still needs to be determined. The glycerol release on day 1 in medium 3 was significantly higher compared to medium 1 and 2. This might be a result of elevated lipolysis during cell death. In this study, a still higher glycerol content was expected; we propose that initial cell death further resulted in disintegration of the cells and nonactive release of triacylglycerides from the lipid vacuoles, which cannot be measured with the method used here. In general, cells in medium 3 seemed to have a general turnover of fatty acids and glycerol because the total number of mature adipocytes decreased less compared with medium 1 and 2.
In general, we were able to detect a higher release of leptin from cells cultured in medium 2 and 3. It was previously described that the expression and secretion of leptin correlated proportionally with body fat mass and adipocyte size.53–55 This supports our findings that cells cultured in medium 2 and 3 were able to keep a higher fat mass and adipocyte typical morphology compared with cells cultured in medium 1. Furthermore, insulin and dexamethasone are thought to be potent stimulators for the leptin expression.44,53,56–58 The effect of insulin is thought to rely on a higher glucose transport and metabolism and less on a direct insulin effect. 59 Another study proposed that insulin regulates the release of leptin from intracellular storages. 56 The higher leptin release from cells cultured in medium 2 or 3 could therefore be an additional result of insulin and dexamethasone in the culture medium.
On the contrary, a high leptin expression is thought to stimulate basal lipolysis.60–62 Siegrist-Kaiser et al. have shown in sugar rats that leptin increased lipolysis in white adipose tissue dosis- and time-dependent. They concluded that leptin has an auto-/paracrine effect on adipocytes, which might be the weight- and fat-reducing activity of leptin. 61 These data show that there might be a cascade of insulin–leptin–lipolysis signalling controlling fatty tissue storage and release. Because of the high production of leptin from cells cultured in medium 2 and 3, this might be another explanation for the elevated glycerol release in these media.
Conclusion
Mature adipocytes were cultured in two commercially available media compared with a control culture medium used in the literature and known to promote dedifferentiation. It was shown that the media supplemented with adipocyte-specific factors prevent cells from dedifferentiation and had a greater effect on cell viability, stability, and functionality compared with the nonsupplemented medium. For the composition and culture of an in vitro adipose tissue equivalent, cell survival and cell functionality are both of great importance. Since medium 2 favors high initial adipocyte survival, it is superior to medium 3. Mature adipocytes can now be used in a wide range of studies and also on a long-term basis to answer questions involving adipocyte metabolism and function. These cells might be highly valuable for the implementation of test systems to investigate cell–cell cross talk in monocultures and cocultures. In the future, mature adipocytes could be used for the composition of implants because of their promising properties concerning their high cell yield during isolation, easy handling, and stability under in vitro conditions. 63
Footnotes
Acknowledgments
The authors thank Dr. Ziegler (Klinik Charlottenhaus, Stuttgart, Germany) for the kind provision of human fatty tissue samples from plastic surgery. We gratefully acknowledge the European Commission for funding the ArtiVasc 3D project under the Seventh Framework Program (Grant Agreement No. 263416).
Disclosure Statement
No competing financial interests exist.