
Abstract
The prolonged ischemia after myocardial infarction leads to a high degree of cardiomyocyte death, which leads to a reduction of normal heart function. Valuable lessons can be learnt from human myocardium and stem cell biology that would help scientists to develop new, effective, safe, and affordable regenerative therapies. In vivo models are of high interest, but their high complexity limits the possibility to analyze specific factors. In vitro models permit analyzing specific factors of tissue physiology or pathophysiology providing accurate approaches that may guide the creation of three-dimensional (3D) engineered cell aggregates. These systems provide a simplistic way to examine individual factors as compared to animal models, and better mimic the reality than 2D models. In this sense, the objective of this work is to better understand the behavior of a human mesenchymal stem cell-like cell line (subcutaneous adipose tissue-derived progenitor cells [subATDPCs], susceptible to be used in cell therapies) when they are embedded in the 3D environment provided by RAD16-I self-assembling peptide (SAP). Specifically, we study the effect in subATDPCs viability, morphology, proliferation, and protein and gene expression of matrix composition (i.e., RGD motif and heparin polysaccharide modifications) in RAD16-I matrix under different media conditions. Results demonstrated that the 3D environment provided by RAD16-I SAP is able to maintain subATDPCs in this new milieu and at the same time its cardiac commitment. Additionally, it has been observed that chemical induction can induce upregulation of cardiac markers, such as TBX5, MEF2C, ACTN1, and GJA1. Therefore, we propose this 3D model as a promising platform to analyze the effect of specific cues that can help improve cell performance for future cell therapy.
Introduction
A
Understanding the underlying biology is the first step to a successful breakthrough in the development of new and efficient therapies. Thus, in vitro models, where the complex cellular microenvironment is deconstructed into simpler and predictable systems, are nowadays being widely studied. Disassemble three-dimensional (3D) microenvironments and the associated processes into adhesive, mechanical, and chemical components would aid to understand the fundamental mechanisms that define the behavior of the cells, developing tissues, and organs.3–6 Additionally, it would help researchers to identify and analyze the role of specific critical factors to regulate stem cell fate and improve the physiological relevance of cell-based assays. 7 These approaches have already provided useful tools for controlling cell differentiation with potential clinical application and could also be important to accelerate translational research.3,8
An overwhelming number of biomaterials have been developed to study as well as to direct cellular interactions in a 3D environment, 6 but the discovery and development of self-assembling peptides (SAP) revolutionized the tissue engineering community.9,10 The nanofibers of these peptides are 1000-fold smaller than synthetic polymer microfibers and similar in scale to natural extracellular matrix (ECM), which better mimics the 3D environment found in vivo. In this work, RAD16-I SAP, which has been shown to support attachment and/or differentiation of a variety of cell types, was used.10–14 This environment does not contain any specific peptide signaling motif and can be defined as a noninstructive milieu from the point of view of cell receptor recognition/activation. 15 This fact allows us to include specific signaling cues to the matrix to analyze their effect on cell behavior.
Several peptide motifs have been described to improve myogenic activity with the aim to maintain phenotype, allow cell–cell contact, establishment of gap junctions, as well as cellular viability. 10 The cell-adhesion motif RGD (R: arginine, G: glycine, D: aspartic acid) that derives from fibronectin is, by far, the most characterized and often employed functionalization system. This recognition motif has shown to display high biological activity on cell anchoring, growth, migration, survival, and differentiation of encapsulated cells. 16 On the other hand, heparin is also an interesting molecule to analyze, since it has been reported to allow better control of local gradients of matrix-binding molecules, such as growth factors (GFs), chemokines, or others, which can be maintained in the matrix and later released.17–22 The sequestering of GFs, externally added or synthesized by cells, helps to localize their activity, protect them from degradation, and in some cases enhance their binding to cell surface receptors. 23 Moreover, heparin can participate in the modulation of various cellular processes such as cell growth, differentiation, morphology, and migration.20,24
In the present work, we present the development of a 3D model based on RAD16-I SAP for the analysis of the cardiogenic potential of cells before implantation in animal models. Specifically, we worked with human subcutaneous adipose tissue-derived progenitor cells (subATDPCs), which are mesenchymal stem cell-like cells (have similar surface markers and have been demonstrated to differentiate into adipogenic and osteogenic phenotypes). 25 These cells are easily obtained in large quantities and have been applied in clinical trials with promising results.26,27 In this study, we test the effect of RGD motif and heparin polysaccharide molecules on subATDPCs behavior by their addition in a 3D RAD16-I matrix. Additionally, the chemical cardiogenic induction was studied to analyze the capacity of subATDPCs to undergo cardiac lineage commitment.
Materials and Methods
Cell culture
Human subcutaneous adipose tissue-derived progenitor cells were isolated from fat pads between the skin and sternum from patients undergoing cardiac surgery in Hospital Germans Trias i Pujol (IGTP). Informed consent was obtained from all subjects, and the study protocol conformed to the principles outlined in the Declaration of Helsinki. Adipose tissue biopsy samples were processed as previously described.28,29 Culture flasks obtained for each patient were maintained for two passages to assure viability, correct growing, and mycoplasma contamination. At passage 2, when cell density was about 20,000 cells/cm2, they were detached using Trypsin-EDTA 0.05% −0.02% (25300-062; Invitrogen) and pools from five different patients were prepared. Each passage was performed when a high level of confluence was reached (90% approx.). The medium was changed on alternate days.
RAD16-I SAP preparation
Four different peptide variations of RAD16-I were analyzed: RAD16-I 0.3% (w/v) without modifications, RAD16-I/RADRGD (95:5) 0.3% (w/v), RAD16-I 0.3% (w/v) with heparin (570:1), and RAD16-I/RADRGD (95:5) 0.3% (w/v) with heparin (570:1). Shortly, 1% RAD16-I nanofiber peptide (PuraMatrix ™, 354250; Corning), was diluted to obtain a solution of 0.3% RAD16-I in sucrose 10% (S1888; Sigma). In parallel, RADRGD (synthesized by Peptide 2.0) 1% water solution was diluted to obtain a solution of RADRGD 0.3%. Afterward, RAD16-I 0.3% and RADRGD 0.3% were mixed in a blend ratio of 95:5. To obtain the peptide with heparin (H3149; Sigma), RAD16-I 0.3% and RAD16-I/RADRGD 0.3% were mixed with heparin 0.01% in a blend ratio of 95:5. Table 1 presents the sequence of the used peptides.
Bold aminoacids represent the minimal recognition motif of laminin for αvβ3 integrin.
Sequences of RAD16-I and RAD16-I modified with RGD.
RGD, arginine, glycine, aspartic acid.
3D cultures
Assembly
SubATDPCs pools were expanded until passage 7, when cells were resuspended in sucrose 10% to render 1.6 × 105 cells in 40 μL. Three-dimensional constructs for each peptide variation, previously described, were prepared mixing an equal volume of the cell suspension and nanofiber peptide obtaining a final cell suspension containing 2 × 106 cells/mL. Three-dimensional cultures were performed as previously reported.11,30 Cells were incubated at 37°C and 5% CO2.
Maintenance of 3D cultures
All 3D constructs were maintained in a control medium during 4 days, when the medium was completely replaced. Half of the 3D constructs were induced with cardio medium (used to test the potential of these cells to differentiate toward a cardiac phenotype) while the other half of the samples was refilled with control medium. Samples were picked 2 and 4 weeks after induction for their analysis, and cells seeded in 2D cultures were taken as control. The composition of both (control and cardiac induction media) is described in Supplementary Data Fig. S2 (Supplementary Data are available online at
Macroscopic and microscopic analysis
SubATDPCs seeded on 2D culture plates were analyzed using Nikon Eclipse TE2000-1 microscope. Macroscopic structure of 3D cultures was analyzed under stereoscopic microscope Nikon Digital Sight DS-2M.
Live and dead staining
Cell viability in the constructs was determined using the LIVE/DEAD® Viability/Cytotoxicity Kit for mammalian cells (L3224; Invitrogen) as previously described. 31
DAPI and phalloidin staining
Cells were double stained using DAPI (D9542; Sigma) and Phalloidin–tetramethylrhodamine B isothiocyanate (7418; Sigma). Briefly, the 3D constructs were fixed for 30 min using 2% paraformaldehyde (PFA) (158127; Sigma) diluted in phosphate buffered saline (PBS) (D8537-500 mL; Sigma) followed by three washes with PBS. Cell membranes were permeabilized in 0.1% Triton X-100 (X100–500 mL; Sigma) solution in PBS with soft shaking during 30 min, followed by three PBS washes. The samples were incubated with Phalloidin-TRITC at a final concentration of 1 μg/mL in PBS for 25 min. DAPI solution (1 μg/mL in PBS) was added into the previous solution and incubated for 5 min to counterstain the nucleus. Samples were carefully washed three times with PBS to eliminate the background and examined under Zeiss AxioVert 200M/ApoTome Microscope.
MTT viability assay
MTT assay is a colorimetric test based on the reduction to formazan crystals, often used as a measure of viable cells. Briefly, the medium was aspirated from the culture and MTT reagent (M5655; Sigma) was added to a final concentration of 0.5 mg/mL in culture medium. The samples were incubated for 3 h at 37°C in the dark. After the incubation, the solution was aspirated, and the constructs were lysed using DMSO (D8418; Sigma). The absorbance was read at 550 nm using the microplate reader, BioTek ELX808.
Study of gene expression by qRT-PCR
Real Time RT-PCR (qRT-PCR) reactions were performed with LightCycler® 480 Real-Time Polymerase Chain Reaction System (Roche), using the iQ™ SYBR® Green Supermix (170-8882; Bio-Rad) as a fluorescent reporter. Briefly, the samples were lysed (two 3D cultures for sample), and RNA was extracted with the peqGOLD Total RNA Kit (12-6634-02; PEQLAB), followed by cDNA synthesis using the QuantiTect Reverse Transcription Kit (205311; Qiagen) according to the manufacturer's protocol. Real-time RT-PCR was run with the following parameters: one cycle of 10 min at 95°C, 50 cycles consisting of 15 s at 94°C, 15 s for primer annealing (Tm dependent on primer pair, see Table 2), and 15 s at 72°C. Finally, melting curve analyses were performed (from 58°C to 95°C) to test the specificity of PCR products, together with agarose electrophoresis. Relative gene fold variations were all determined by the comparative CT method (2−ΔΔCt) and expression of the target genes was normalized to the housekeeping gene (ribosomal protein 22L, RPL22).
Early and definitive cardiac markers were analyzed.
Western blotting for protein analysis
Protein extracts were obtained using RIPA buffer (R0278; Sigma) containing the proteinase inhibitor cocktail (11836153001; Roche). Two 3D cultures constituted each sample for western blotting and they were lysed with the help of pestles to crush the samples. All samples were centrifuged and the protein content was quantified from the supernatant using the Micro BCA™ Protein Assay Kit (23225; Pierce Thermo Scientific). Equal amounts of total protein (5 μg) were denatured with sodium dodecyl sulfate (SDS)(L5750-500G; Sigma) and β-mercaptoethanol (M3148; Sigma) at 95°C during 10 min. Subsequently, the proteins were separated by electrophoresis on 10% SDS–PAGE gels at 150 V during 90 min. Proteins were blotted onto PVDF membrane (LC2005; Invitrogen) during 2 h at 40 V. The membranes were blocked with 5% nonfat milk buffer during 2 h at room temperature and incubated with rabbit polyclonal anti-GATA4 1 μg/mL (sc-9053; Santa Cruz) or rabbit polyclonal anti-GJA1 1 μg/mL (sc-9059; Santa Cruz) at room temperature during 1 h. The membranes were washed with PBST (PBS supplemented with Tween-20 [P-1379; Sigma]) and incubated with peroxidase-conjugated rabbit IgG goat polyclonal secondary antibody 1 μg/mL (ab97051; Abcam). The protein bands were detected after incubation with SuperSignal® West Pico Chemiluminescent Substrate (34080; Thermo Scientific) using ImageQuant LAS 4000 mini equipment. Goat polyclonal anti-Actin antibody (sc-1615; Santa Cruz) was used to normalize.
Dynamic mechanical analysis
Viscoelastic properties of 3D constructs were analyzed by dynamic mechanical analysis (DMA) after 2 and 4 weeks using control and cardio medium. A compression assay with DMA Multifrequency–Strain mode and a frequency sweep test was applied to each sample with a DMA Q800 (TA Instruments). The construct was deformed (oscillated) at constant amplitude (strain) over a constant frequency, and the mechanical properties were measured. The conditions of the assay were: Amplitude = 1, Preload force = 0.03 N, and Frequency = 1 Hz. The selected frequency is the standard working frequency used in this type of experiments and the selected amplitude is within a range of amplitude values where the samples remained constant. Results were obtained with TA Instrument Explorer software and analyzed with TA Universal analysis software. Values of storage modulus (G′), loss modulus (G″), complex modulus (G*), and tan delta were obtained and represented in separated graphics. G′ is the measure of the sample's elastic behavior, G″ measures the viscous response of the material, G* is the sum of both components and, finally, the ratio of the loss to the storage is the tan delta and it is a measure of the energy dissipation of the material.
Statistics
GraphPad Prism (version 5.00 Windows; GraphPad Software, San Diego, CA,
Results
SubATDPCs behavior in 2D cultures
SubATDPCs cultivated in classical cell culture flasks with control medium presented an elongated, individual morphology, but the addition of cardiac induction medium promoted star-shaped morphology and cluster formation as it can be observed in Figure 1a. Additionally, the growth pattern showed that control medium allowed slow but constant proliferation over time, whereas cardiac induction medium stopped the proliferation presumably due to a differentiation process (Fig. 1b). Therefore, the morphology and the growth rate of subATDPCs growing on flat surfaces highly depend on the culture condition.
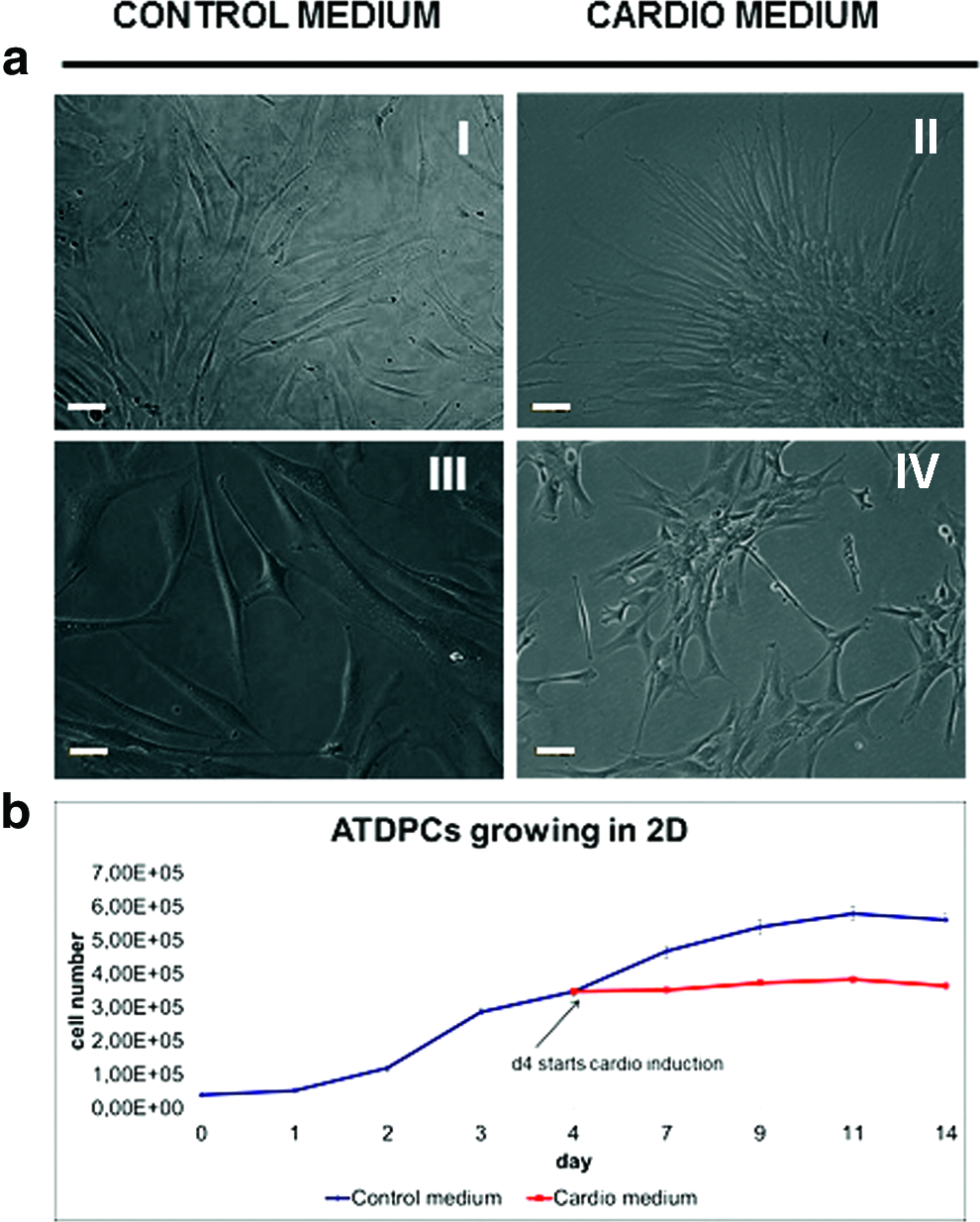
Subcutaneous adipose tissue-derived progenitor cells (subATDPCs) growing on flat surfaces.
SubATDPCs behavior in 3D cultures
The best conditions in terms of cell density and peptide concentration for subATDPCs cultivated in 3D SAP RAD16-I nanofiber environment were assessed. Regarding cell seeding density, values between 2 × 106 and 4 × 106 cells/mL were tested to analyze the best condition to allow cell movement and cell–matrix interaction. The results showed that the use of high cell concentration (4 × 106 cells/mL) leads to amorphous cluster formation and large number of dead cells (data not shown). In terms of peptide concentration, different RAD16-I peptide values were tested using a cell density of 2 × 106 cells/mL. Figure 2 shows cell viability and construct diameter over time depending on initial peptide concentration. Additionally, the effect of cardiac induction medium in the construct morphology and cell viability was analyzed.
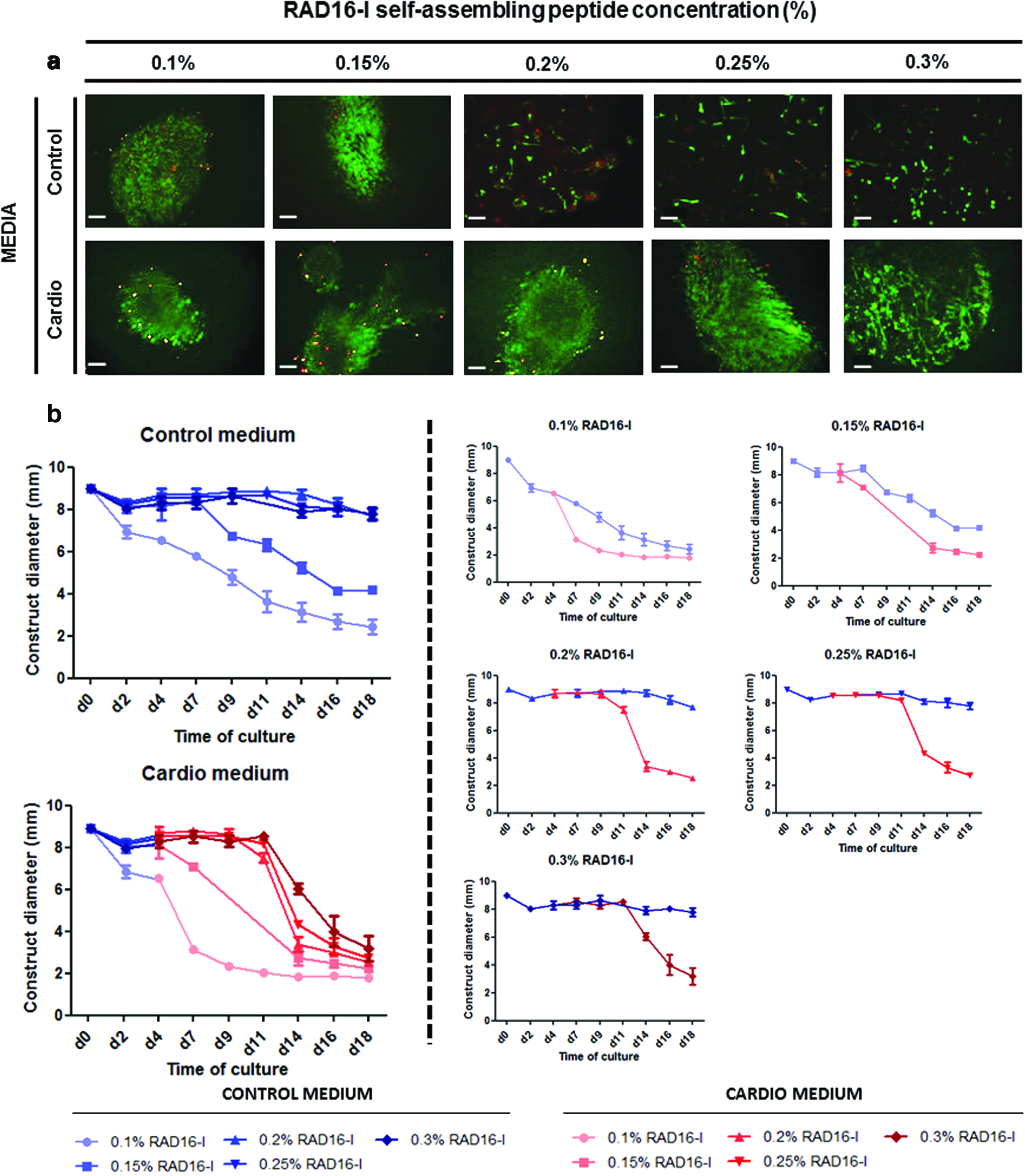
SubATDPCs growing in different RAD16-I self-assembling peptide (SAP) concentration.
In all tested conditions high viability was observed; however, high peptide concentrations led to a lower reduction in construct diameter. Once embedded, cells are able to elongate processes by networking one to each other and establishing connections with the matrix. This latter exerts forces that could end in a construct diameter reduction. 31 Interestingly, it was observed that at high peptide concentration (from 0.2% to 0.3%) subATDPCs cultured in control medium were not able to elongate properly and as a consequence, the construct did not condense. On the other hand, the addition of cardiac induction medium allowed the cells to elongate cellular processes to increase cell–cell contact and establish cell network, which leads into a construct condensation. After 18 days almost all constructs grown in cardiac induction medium, indistinctly of peptide concentration, presented a diameter between 2 and 4 mm. In view of all these results, it was concluded that the most appropriate conditions to analyze subATDPCs behavior using RAD16-I peptide to recreate a 3D model in vitro were, RAD16-I 0.15% at cell density of 2 × 106 cells/mL. In Figure 3 an overview of subATDPCs behavior in these conditions is presented. In these specific conditions, the morphogenic process of construct diameter reduction was faster in the case of constructs grown in cardiac induction medium, but constructs grown in control medium also displayed this condensation (Fig. 3a). As it can be observed in Figure 3b, cells appeared round shaped before encapsulation. During the first 4 days in which cells remained in control medium, they started to elongate. Importantly, cells remained alive in both media conditions after 20 days of culture and, interestingly, cardiac induction medium induced some kind of cell alignment (Fig. 3c).

SubATDPCs growing at a cell density of 2000 cells/μL in RAD16-I 0.15%.
Matrix modifications
The behavior of subATDPCs cultured in RAD16-I SAP containing heparin molecules and the motif RGD for cell integrin receptor recognition was analyzed. High cell viability was observed in all tested matrix combinations after 11 days of culture (Live/Dead assay, Fig. 4a). Moreover, DAPI and phalloidin fluorescent images in Figure 4a showed that scaffolds composed of RAD16-I/RADRGD motifs presented a higher cell density as compared to RAD16-I peptide alone. Therefore, we suggest that RGD motif was indeed enhancing elongation and, as a consequence, maybe cell–cell contact and cell migration. Regarding the presence of heparin in the scaffold, it was shown to enhance cell elongation and, interestingly, in combination with RGD motif and by visual inspection, cells seemed to form clusters and looser networks. Additionally, it looks like constructs that presented RGD motifs condensed more than the constructs without it. Moreover, no differences were appreciated macroscopically with the addition of heparin (Fig. 4b).
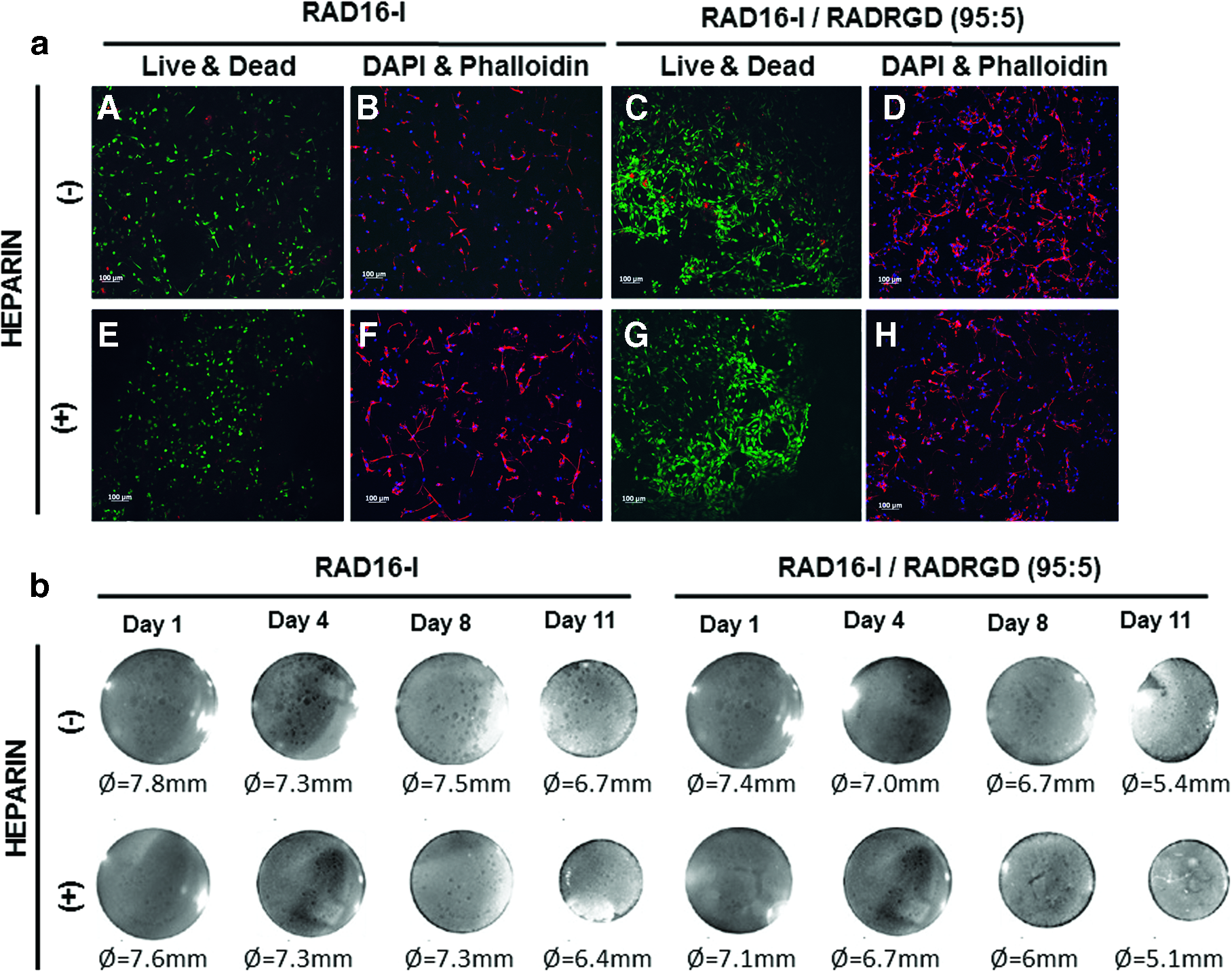
SubATDPCs encapsulated in RAD16-I nanofiber peptide and RAD16-I with modifications.
On the other hand, during the first days of culture, modified peptides showed higher cell growth than cells grown in unmodified peptide. However, after 4 days there were no significant differences among them (Fig. 5 A, B). Interestingly, the cells that grew in bare RAD16-I SAP in cardiac induction medium started to decrease their absorbance after 11 days of culture (Fig. 5a), probably as a consequence of condensation. Careful analysis of the constructs after 20 days of culture showed that cells migrate outward the construct (Supplementary Data Fig. S1). This effect was also observed in other conditions, where 3D cultures were considerably condensed. The morphology of cytoskeleton and nuclei (Fig. 6) showed elongation and high interconnection between cells after 6 days of culture. Additionally, cells grown in RAD16-I with heparin after 13 days of culture appeared highly organized, whereas samples that did not contain these polysaccharide molecules showed a disordered pattern (Fig. 6). Viability assays confirmed that cells remained alive after 13 and 21 days of culture, respectively.

MTT assay of SubATDPCs 3D cultures. SubATDPCs viability was also assessed quantitatively with MTT assay.

Effect of heparin in 3D cultures of subATDPCs. D&P staining after 6 days of culture. Nuclei stained in blue and actin filaments in pink. L&D staining after 13 and 21 days of culture. Live cells stained in green and dead cells in red. Scale bars 100 μm and close ups 20 μm. Color images available online at
Gene and protein expression of subATDPCs encapsulated in RAD16-I peptide with and without the presence of RGD motif and heparin molecules were analyzed. TBX5 and MEF2C early cardiac markers, and ACTN1 and GJA1 definitive cardiac markers were quantified at gene expression level (Fig. 7a) and GATA4 (also early cardiac marker) and GJA1 at protein expression level (Fig. 7b). As it can be observed in Figure 7a, an upregulation of TBX5, MEF2C, and ACTN1 genes was detected in subATDPCs cultured in 3D environment in control medium as compared to 2D cultures. Moreover, the combination of 3D environment and cardiac induction implied a significant upregulation of TBX5, MEF2C, and GJA1, but ACTN1 was much more expressed in 2D cultures with cardiogenic medium than in the 3D environment. The obtained results did not present significant differences in terms of gene expression for constructs modified with RGD motif or heparin either in control or cardiac induction media. At protein level (Fig. 7b) western blot results revealed that GATA4 transcription factor and GJA1 were up-regulated in cardiac induction medium in contrast to control medium. Although, subATDPCs growing on flat surfaces with cardiac induction medium showed expression of GATA4 and GJA1, the environment provided by 3D systems up-regulated the expression of both proteins. Neither the presence of RGD motif or heparin polysaccharide increased GATA4 or GJA1 expression in comparison with RAD16-I peptide (Fig. 7).

Early and definitive cardiac marker expressions by subATDPCs growing in RAD16-I 0.15% with and without modification depending on culture media.
Another important aspect to analyze was the mechanical properties of the 3D constructs cultured with control and cardiac induction media at different time points (Fig. 8a). Regarding the elastic component, constructs cultured with control medium presented lower G′ values than constructs cultured with cardiac induction medium, except in the case of constructs without heparin at 4 weeks of culture that presented a storage modulus similar to those cultured with cardiac induction medium. The loss (G") and complex (G*) modulus presented a similar tendency than the storage modulus. Moreover, all samples presented G" values lower than G′ values, which meant that constructs are more elastic than viscous. Finally, tan delta, which gives an idea of full viscoelastic properties of the constructs, indicated that samples cultured with cardiac induction medium and the ones cultured with control medium and without heparin at 4 weeks of culture have the same viscoelastic nature and dissipate the energy equally. However, samples grown in control medium (except those without heparin at 4 weeks of culture) presented higher tan delta values, indicating a behavior more viscous than elastic.

Dynamic mechanical analysis and study of the effect of nanofiber RAD16-I concentration at protein level.
We also examined the effect of the material stiffness at the time of the encapsulation (initial conditions). SubATDPCs were cultured in RAD16-I and RAD16-I/RADRGD at 0.15% (120 Pa) and 0.2% (220 Pa), respectively. As it has been stated in Figure 2, subATDPCs cannot elongate and the aggregate do not condense at high peptide concentration in control media. For this reason, this experiment was performed culturing the cells in a cardiogenic medium. As it can be seen in Figure 8b, either initial stiffness conditions or matrix modification presented a strong effect by downregulating the protein expression (p < 0.0001). This effect could be observed clearly at 2 weeks of culture with less significant changes at 4 weeks. GATA4 and GJA1 proteins were significantly downregulated (p < 0.0001) when the initial stiffness of the matrix was 220 Pa as compared with an initial stiffness of 120 Pa. The addition of RGD motifs in the 3D structure lead to a significant reduction of both protein expressions (p < 0.0001) as compared with RAD16-I control matrix.
Discussion
Heart is a complex tissue whose behavior is affected by multiple factors. The understanding of the individual effect of each factor can be of great interest for the development of further therapeutic treatments, but the study of all these factors and their combination in vivo proved extremely difficult. 6 The deconstruction of heart tissue using in vitro models can help to elucidate the effect of these factors on healthy or pathologic tissue.32–34 Interestingly, by combining different materials and different cell types, it is possible to analyze different parameters. Each of these combinations leads to really distinct systems that allow analyzing a wide range of specific parameters.35–40
In this work, a specific system of subATDPCs embedded in a 3D matrix of RAD16-I SAP with and without integrin receptor binding (RGD) and heparin molecules were analyzed. This 3D model has been widely used with different cell lines, but slight changes such as peptide concentration and cell seeding density must be assessed for each cell type.13,14,30,31,41 In this study, particular attention was focused on cellular connectivity. This concept is really important in cardiac tissue since, in vivo, the synchronous action of CMs lead to the obtention of a functional tissue. 42 Therefore, all the mentioned parameters were switched to obtain a good milieu where cells could interact forming an intricate network mimicking as optimally as possible the native tissue.
Here, we demonstrated that the 3D environment provided by RAD16-I SAP is able to maintain the cardiac commitment of subATDPCs, without significant changes in gene or protein expression. However, chemical induction, which is used to promote cells to undergo cardiac lineage, induce the upregulation of cardiac markers such as TBX5, MEF2C, ACTN1, and GJA1. Moreover, the viscoelastic behavior of the entire constructs at different time points could provide us an idea about the evolution of the scaffold in terms of matrix remodeling. The full mechanical response of constructs cultured with cardiogenic medium was quite different from those cultured with control medium at 2 weeks of culture. The tendency changes at 4 weeks of culture, where constructs without heparin cultured with control medium have a similar behavior than constructs cultured with cardiogenic medium. Depending on the conditions provided during culture time, cells could behave differentially in terms of cell–cell and cell–matrix interactions driving internal forces to reduce the macroscopic diameter of the constructs. Moreover, ECM components synthesized by the cells and the constant turnover of ECM proteins could also influence the viscoelastic properties of the construct. Interestingly, regardless of the initial components of constructs cultured with cardiogenic medium, the viscoelastic nature is maintained, which could indicate a strong influence of the chemical induction.
These in vitro studies could provide valuable information for further in vivo experiments. Interestingly, a good viability profile of subATDPCs encapsulated inside RAD16-I SAP with and without RGD motif and heparin polysaccharide was observed after 13 and 21 days of culture. These results lead us to hypothesize that subATDPCs would afford in vivo implantation. Furthermore, the cytotoxicity of materials on stem cells was assessed using an MTT assay, observing no undesirable results. Additionally, the cell alignment presented in the new nanofiber is a good starting point to obtain cells with a degree of organization, which suggests their potential similarity to CMs distribution. At this point, considerable research is ongoing to try to functionally differentiate these cells using SAP platforms.
Footnotes
Acknowledgments
The authors wish to thank the patients who made this study possible and the members of the Department of Cardiac Surgery for their collaboration in obtaining human samples. The research leading to these results has received funding from the European Union Seventh Framework Program (FP7/2007-2013) under grant agreement n°229239. The authors thank the SUR of DEC of Catalan Government and the ESF for the predoctoral fellowship 2015FI_B2 00109 to L.R.S. They also thank the Materials Science Laboratory of IQS School of Engineering for the kind help with the DMA analysis. Finally, they thank M.P.A. from the Genomic Unit of the IGTP for her help with the quantitative PCR.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.