
Abstract
Chondrogenic differentiation of adipose-derived stem cells (ASC) is challenging but highly promising for cartilage repair. Large donor variability of chondrogenic differentiation potential raises the risk for transplantation of cells with reduced efficacy and a low chondrogenic potential. Therefore, quick potency assays are required to control the potency of the isolated cells before cell transplantation. Current in vitro methods to analyze the differentiation capacity are time-consuming, and thus, a novel enhancer and tissue-specific promoter combination was used for the detection of chondrogenic differentiation of ASC in a novel quick potency bioassay. Human primary ASC were cotransfected with the Metridia luciferase-based collagen type II reporter gene pCMVE_ACDCII-MetLuc together with a Renilla control plasmid and analyzed for their chondrogenic potential. On day 3 after chondrogenic induction, the luciferase activity was induced in all tested donors under three-dimensional culture conditions and, in a second approach, also under two-dimensional (2D) culture conditions. With our newly developed quick potency bioassay, we can determine chondrogenic potential already after 3 days of chondrogenic induction and under 2D culture conditions. This will enhance the efficiency of testing cell functionality, which should allow in the future to predict the suitability of cells derived from individual patients for cell therapies in a very short time and at low costs.
Introduction
C
Luciferase-based bioassays represent a useful alternative to conventional analytical methods due to their simple and extremely sensitive properties. The assay used in this work is based on measuring bioluminescence derived from the reaction of Metridia luciferase under control of a tissue-specific collagen type II promoter coupled to a signal-amplified viral enhancer cytomegalovirus (CMV). 9 With our luciferase reporter system, we have established a proof of principle concept for the prediction of the chondrogenic differentiation potential within 3 days.
Materials and Methods
ASC isolation and cultivation
The collection of human tissue was approved by the local ethics board with patient's consent. Subcutaneous human adipose tissue was obtained during routine outpatient liposuction procedures under local tumescent anesthesia. Isolation of ASC was performed according to Wolbank et al. 10 After isolation, cells were cultured in the endothelial growth medium (EGM-2; Lonza) at 37°C, 5% CO2, and 95% air humidity to a subconfluent state before passaging. ASC were seeded at a density of 2 × 103/cm2, and media were changed every 3–4 days. ASC from passage 1 were used for flow cytometry, histological analysis, and reporter experiments. For qRT-PCR experiments, cells were expanded up to passage 2–4.
ASC immunophenotype
ASC from passage 1 were characterized by flow cytometry analysis using the following antibodies: CD73-PE (BD), CD90-PE (BD), CD105-FITC (Abcam), CD14-FITC (Immunotools), CD34-PE (Immunotools), CD45-FITC (BD), HLA-ABC-PE (BD), and HLA-DR-FITC (BD). For staining, 2 × 105 cells in 50 μL phosphate-buffered saline (PBS) with 1% fetal calf serum were incubated with 5 μL primary labeled antibodies at room temperature for 15 min in the dark. Cells were washed with 1.5 mL Cell Wash™ (BD) and centrifuged for 5 min at 400 g. The supernatant was discarded and the cell pellet resuspended in 300 μL 1× Cell Fix™ (diluted 1:10 with aqua dest; BD) and analyzed on a FACSCanto (BD).
Chondrogenic three-dimensional pellet culture
For chondrogenic differentiation and three-dimensional (3D) micromass pellet cultures, 3 × 105 ASC were centrifuged in chondrogenic differentiation media (hMSC Chondro BulletKit [Lonza] containing 10 ng/mL BMP-6 [R&D Systems] and 10 ng/mL TGF-β3 [Lonza]) in 1.5-mL polypropylene screw cap microtubes (Sarstedt). The tubes were placed in an incubator at 37°C, 5% CO2, and 95% humidity with slightly open cap for gas exchange. After 2 days, the pellets were transferred to 96-well U-bottom plates. Media were changed every 2–3 days.
Histological analysis
The 3D micromass pellet cultures incubated over 35 days in chondrogenic differentiation media were measured once a week for their cross-section area. Micromass pellets were fixed in 4% phosphate-buffered formalin overnight for histological analysis. The next day, the pellets were washed in 1× PBS and dehydrated in increasing concentrations of alcohol. After rinsing the pellets in xylol and infiltration with paraffin, deparaffinized sections were stained with Alcian blue for 30 min and counterstained for 2 min with Mayers hematoxylin. For immunohistochemical staining, sections were treated with pepsin for 10 min at 37°C (AP-9007 RTU; Thermo Scientific). Endogenous peroxidase was quenched with freshly prepared 3% H2O2 for 10 min at room temperature, followed by normal horse serum 2.5% (Vector RTU) to block unspecific binding. Sections were incubated 1 h with monoclonal anti-collagen type II (MS-306 P0; Thermo Scientific) at 1:100. After washing with TBS, sections were incubated with the secondary antibody (anti-mouse DAKO EnVision+ System HRP-labeled Polymer; Dako) for 30 min and rinsed in TBS again. Bindings were visualized using Nova Red (SK4800; Vector Labs) for 6 min. Counterstaining was performed with Mayers hematoxylin for 2 min. For quantification of the collagen type II immunohistolgical staining intensity, ImageJ 1.47v (National Institutes of Health) was used.
Quantitative RT-PCR
Samples for qRT-PCR were taken after 3 days and at the end of differentiation cultures (day 35). Three to five micromass pellets per donor were pooled in 1 mL TRI Reagent (Sigma) and incubated for 20 min at room temperature. Extraction was facilitated by repeated pipetting of the pellets. Total RNA isolation was performed according to the TRI Reagent protocol (Sigma), and RNA content and RNA integrity of the samples were assessed by an Agilent 2100 Bioanalyzer, using the RNA 6000 Nano Chips Kit (no. 5065–4476; Agilent Technologies). Isolated RNA was transcribed to cDNA according to the High Capacity cDNA Archive Kit protocol (Applied Biosystems). Quantification of specific cDNAs was conducted in triplicate using a LightCycler W 480 (Roche) and TaqMan gene expression assay (Applied Biosystems) for Col2A1 (collagen type II, Hs01064869_m1) and Sox9 (Hs00165814_m1). The PCR was programmed as follows: initial denaturation at 95°C for 10 min, followed by 95°C for 10 s, 60°C for 45 s, cycled 50 times. Cooling to 40°C was held for 30 s. Slope speed was 20°C/s. Standard curves were prepared for quantification and expression values were normalized to the housekeeping gene, hypoxanthine–guanine phosphoribosyltransferase (HPRT). The efficiency-corrected quantification was performed automatically, using LightCycler 480 Relative Quantification Software (Roche).
Reporter plasmid
All cloning procedures were carried out using a standard Taq hot-start polymerase (Peqlab Biotechnologie GmbH) and the Escherichia coli cloning strain TOP10. The human collagen II alpha 1 promoter (Col2A1 promoter and 5′ mRNA UTR from −478 to +179 from the transcriptional start site, see NC_000012.12) was cloned from human genomic DNA isolated from whole blood using the cloning primers hColIIPs (5′-CTGTGGGCTCCTCCCTGTCCCCACTCC-3′) and hColIIPas (NotI) (5′-GAGGCGGCCGCGCTCACCGCGGGGCCTGGCTGAGC-3′) (Microsynth AG) and the product was ligated into the vector pCR2.1 by TA cloning for sequencing (data not shown) and subsequent cloning. The promoter was transferred to the in-house designed plasmid pCMVE-EYFPHis containing the CMV enhancer (from −524 to −120 from the transcriptional start site, according to 9 ) and an EYFPHis reporter gene by PCR amplification using the primers hC2PHIII_f. (5′-GAG AAG CTT CTG TGG GCT CCT CCC TGT CC-3′) and hC2PHIII_r (5′-GAG AAG CTT GCT CAC CGC GGG GCC TGG-3′) (Microsynth AG), transfer of the PCR product to pCR2.1 and subsequent cloning by HindIII (Fermentas/FisherScientific) digestion of insert and destination vector and ligation into pCMVE_ACDCII using standard procedures. The EYFPHis reporter gene was exchanged by a Metridia-secreted luciferase by cloning the Metridia luciferase open reading frame 3′ of the CMV enhancer and the collagen II promoter in pCMVE_ACDCII using AgeI (Fermentas/Fisher Scientific) and NotI (Fermentas/Fisher Scientific) restriction sites. All vectors and intermediate products were verified by control digests (data not shown) and the Metridia luciferase containing end product pCMVE_ACDCII-MetLuc (Col2A, Fig. 1) was verified by sequencing (data not shown).
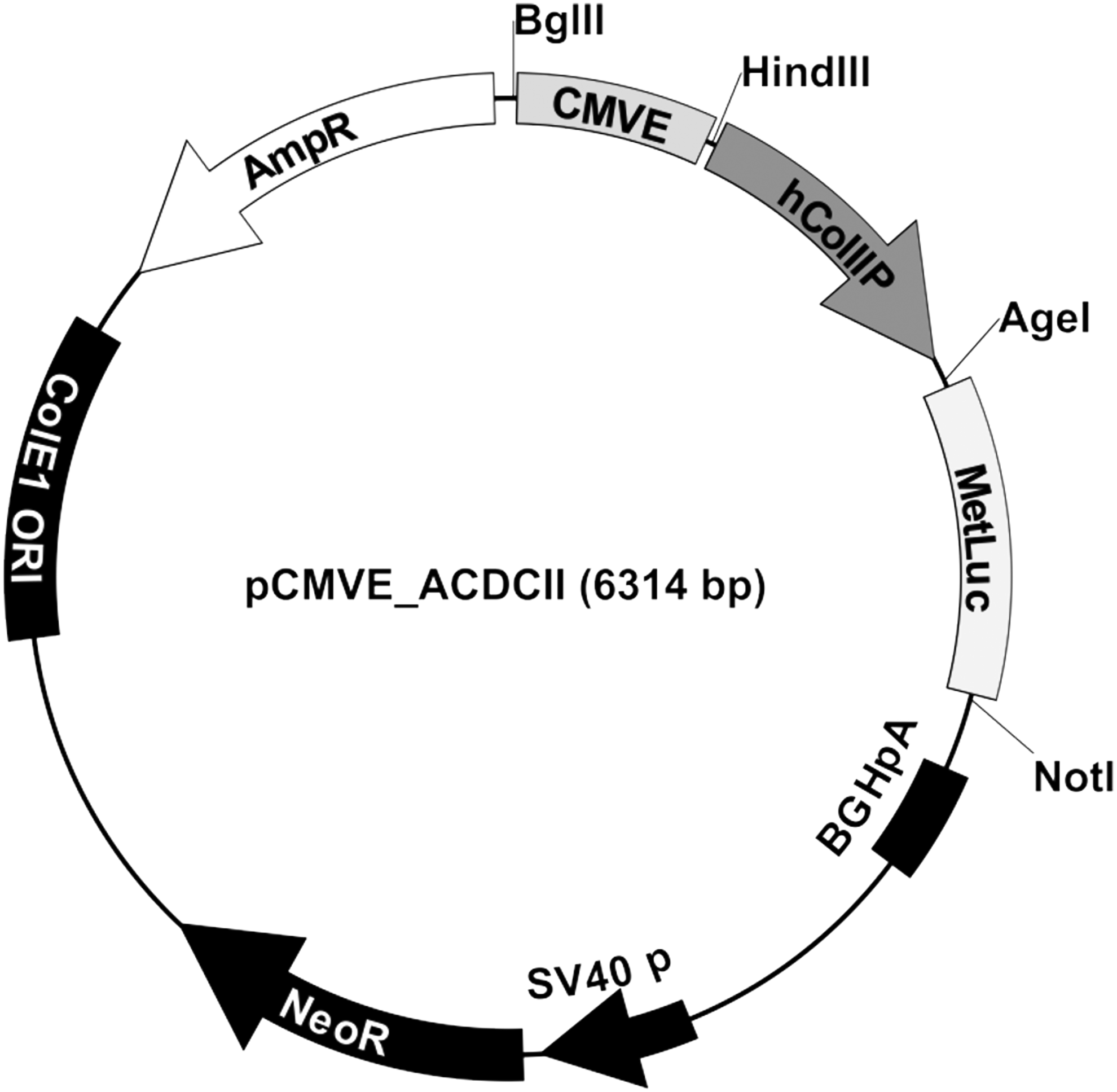
Metridia luciferase-based reporter vector pCMVE_ACDCII-MetLuc. This construct utilizes an artificial, chimeric cis-acting regulatory sequence through combination of a pan-active cytomegalovirus enhancer cloned directly upstream of the human COL2A promoter, which drives the expression of the reporter gene.
Luciferase assay
Pellets were cotransfected with Metridia luciferase-based reporter genes pCMVE_ACDCII-MetLuc together with the Renilla control plasmid using X-tremeGENE HP DNA Transfection Reagent (Roche) and 0.4 μg DNA in a 3:1 reagent/DNA ratio after 2 days of pellet formation. For chondrogenic two-dimensional (2D) luciferase assay, 3 × 104 ASC per well were seeded in a white 96-well plate (Fisher Scientific) in EGM-2 media (9.4 × 104/cm2). Sixteen hours later, media were replaced by differentiation media. ASC were cotransfected with Metridia luciferase-based reporter vector pCMVE_ACDCII-MetLuc together with Renilla control plasmid using the same concentrations like for 3D pellet transfection. The Metridia luciferase activity was measured in the supernatant samples (50 μL) with the Ready-To-Glow™ Secreted Luciferase Reporter System (Clontech) according to the manufacturer's protocol on day 1, 2, and 3. For the evaluation of this bioassay and as internal control to normalize the transfection, we cotransfected the cells with a Renilla reporter gene under control of a ubiquitary promoter. Before measurement of Renilla luciferase activity, ASC were lysed with 1× Renilla Luciferase Assay Lysis Buffer (Promega) for 15 min and pellets with 5× Renilla Luciferase Assay Lysis Buffer for 15 min at 1400 rpm. Then, the Renilla luciferase activity was measured using the Renilla Luciferase Assay System (Promega) according to the manufacturer's instructions. Luciferase activities were recorded with an Infinite® M200 Multimode Microplate Reader (Tecan). Data were normalized for transfection efficiency to Renilla luciferase activity. Pellet cultures and adherent cells were transfected using the green fluorescent protein (GFP) to evaluate transfection efficiency. GFP-positive pellets and cells were microscopically analyzed (Axiovert 200; Zeiss).
Statistical analysis
Data are presented as mean ± standard deviation and statistical analysis was performed using PRISM6 (GraphPad). p-Values of <0.05 were considered to be significant acquired by parametric two-way analysis of variance-Bonferroni post hoc or parametric two-tailed t-test assuming normal distribution according to the Kolmogorov–Smirnov normality test.
Results
Expression of specific ASC surface markers
ASC displayed strong expression of MSC marker CD73 (99.68% ± 0.35%), CD90 (98.70% ± 1.46%), and CD105 (97.95% ± 1.62%) and almost no expression of the lymphatic marker CD14 (1.64% ± 0.87%), the endothelial marker CD34 (3.90% ± 7.69%), and the hematopoietic marker CD45 (1.23% ± 0.52%) (Supplementary Table S1; Supplementary Data are available online at
Definition of donor potential by qRT-PCR, histology, and pellet diameter area
Micromass pellets from nine different donors were analyzed after 35 days of chondrogenic differentiation for their capability to express collagen type II at the mRNA and protein level by means of qRT-PCR and immunohistological staining. Furthermore, sulfated glycosaminoglycans (sGAGs) were stained with Alcian blue to analyze the potential of individual donors to synthesize proteoglycans. Donors 3, 4, 5, 8, and 9 showed Col2A1 expression levels in the range of 70.8–354.8 (relative mRNA expression normalized to HPRT), whereas donors 1, 2, 6, and 7 demonstrated significantly lower values in the range of 0.0066–3.4 (Fig. 2a, b). Immunohistological stainings showed that donors with low Col2A1 mRNA expression have also low potential to produce collagen type II and sGAGs on protein level, while donors with high Col2A1 expression on mRNA level showed intense staining for Alcian blue and collagen type II (Fig. 3a). Based on the results of qRT-PCR and histological evaluations, we defined donors with high and low differentiation potential, as “good donors” and “bad donors.” The pellet diameter area of each good and bad donor was evaluated over time in culture (day 7, 14, 21, 28, and 35) (Fig. 3b). These results corroborate the immunohistological stainings for Alcian blue and collagen type II. Donor 2 is in the range of the good donors regarding pellet diameter area but is still negative for Alcian blue and collagen type II staining. According to the analyses, the threshold for Col2A1 expression on mRNA level was set to 10 and together with the histological and immunohistological data, we identified five good donors (3, 4, 5, 8, 9) and four bad donors (1, 2, 6, 7).
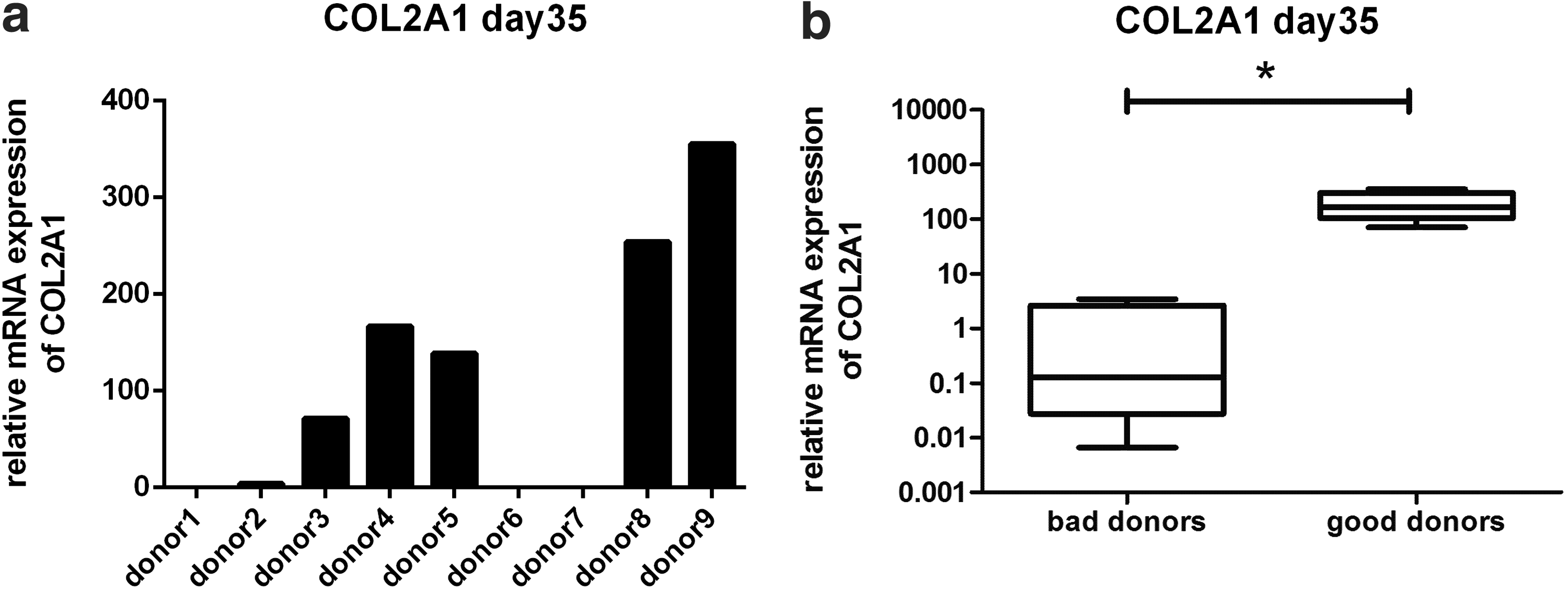
COL2A1 mRNA expression. On day 35 of micromass pellet cultures, three to five pellets were pooled from each donor. COL2A1 mRNA expression demonstrates variations between the nine tested donors

Alcian blue and collagen type II staining
Activation of Col2A luciferase reporter under 3D culture conditions
To visualize transfection efficiency of 3D micromass pellets, a GFP plasmid was used (Supplementary Fig. S1a). The weak fluorescence of untransfected pellets is probably due to the known autofluorescence of cartilage-like matrix. 12 For the analysis of Col2A reporter activation, micromass pellets of the nine different donors were transfected with a plasmid containing Metridia luciferase under control of the Col2A promoter together with a plasmid containing Renilla luciferase under control of a ubiquitary promoter. After 3 days of differentiation, the activation of the Col2A luciferase reporter was analyzed and normalized to Renilla luciferase. While there were remarkable differences in the activation of the Col2A reporter (Fig. 4a), all donors showed similar activation of Renilla luciferase (Fig. 4b), indicating similar transfection efficiency and promoter activation. These differences in the Col2A reporter activation also occurred after normalization to Renilla activation (Fig. 4c).
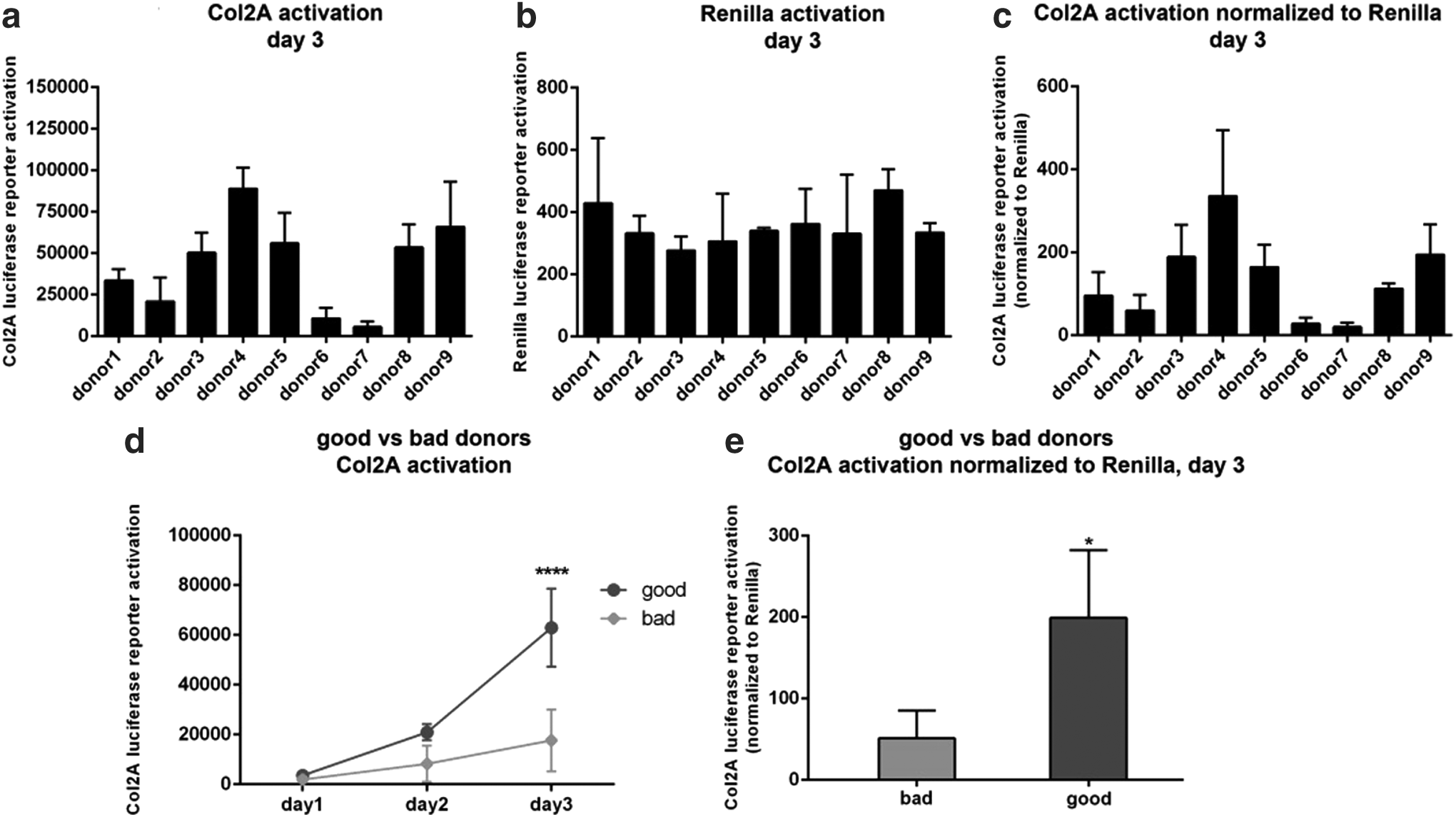
Activation of Col2A luciferase reporter under 3D culture conditions. Micromass pellets were cotransfected with Metridia luciferase-based reporter gene pCMVE_ACDCII-MetLuc together with Renilla control plasmid. Pellets were incubated in chondrogenic differentiation media with TGF-β3 and BMP-6. Col2A Metridia luciferase activity was measured in the supernatant on day 1, 2, and 3. For measurement of Renilla luciferase activity, pellets were lysed on day 3. Activation of Metridia and Renilla luciferase reporter and normalization of Metridia luciferase to Renilla luciferase on day 3 for each single donor
The data were compared to the results of qRT-PCR and histological stainings. Good donors exhibited relative light units (RLU) in the range of 5.0 × 104–8.9 × 104 of Col2A activation corresponding to relative values of 112–335 after normalization to Renilla, while bad donors were in the range of 5.5 × 103–3.4 × 104 and 20.1–95.7 after normalization (Fig. 4a, c). Sequential analysis of Col2A reporter activation over the first 3 days of induction showed an increase for both groups, but significantly higher Col2A activation for good donors on day 3 (Fig. 4d). Even after normalization of Col2A activation to Renilla activation, the induction of Col2A reporter was significantly stronger in good donors (199 ± 83) compared to bad donors (51 ± 34) (Fig. 4e).
Col2A1 and Sox9 mRNA expression under 3D culture conditions
As we have observed a luciferase reporter signal only after 3 days of differentiation, we performed qRT-PCR for Col2A1 and Sox9. After 3 days, Col2A1 expression was not detectable for any donor except minimal levels for donor 2 (Col2A1 mRNA expression of 0.0154) (data not shown). Early marker Sox9 expression after 3 days of differentiation showed also a very low but detectable signal in all donors (Supplementary Fig. S2a). However, no difference between bad and good donors could be discriminated (Supplementary Fig. 2b).
Activation of Col2A luciferase reporter under 2D culture conditions
To simplify the assay, we changed from the standard chondrogenic 3D culture conditions to 2D culture conditions, which reduces time and required cell numbers. Transfection efficiency was analyzed by using a GFP plasmid (Supplementary Fig. S1b) corresponding to 45.3% ± 11.8% of transfected cells as determined by nucleocounting. ASC were seeded and induced in 2D culture and cotransfected with Col2A reporter plasmid together with Renilla luciferase control plasmid. There was a clear difference in the activation of Col2A reporter between good (6.2 × 105–1.1 × 106 RLU) and bad donors (2.4 × 105–7.0 × 105 RLU) found on day 3 of culture (Fig. 5a), while Renilla activation was similar in all tested donors (Fig. 5b). The differences in Col2A reporter activation were still present after normalization of Metridia to Renilla activation (19.9–31.3 vs. 7.3–18.3) (Fig. 5c), which demonstrates that the potential for chondrogenic differentiation is reflected in Col2A promoter activity at early time points even under 2D conditions.

Activation of Col2A luciferase reporter under two-dimensional culture conditions. Adipose-derived stem cells were cotransfected with Metridia luciferase-based reporter gene pCMVE_ACDCII-MetLuc together with Renilla control plasmid and incubated for 3 days with chondrogenic differentiation media with TGF-β3 and BMP-6. On day 1, 2, and 3, Col2A Metridia luciferase activity was measured in the supernatant. On day 3, cells were lysed for measurement of Renilla luciferase activity. Activation of Metridia and Renilla luciferase reporter and normalization of Metridia luciferase to Renilla luciferase on day 3 for each single donor
Sequential analysis of Col2A reporter activation over the first 3 days showed an increase for both groups, but again a significantly higher Col2A activation for good versus bad donors on day 3 (Fig. 5d). Moreover, activation of Col2A reporter was still higher in good donors (25.6 ± 4.3) compared to bad donors (14.2 ± 4.8) after normalization to Renilla activation (Fig. 5e).
Relationship of Col2A luciferase reporter activation to Col2A1 mRNA expression, collagen type II immunostaining, or pellet diameter area
For a more clear relationship of the 3D micromass pellet data, our day 3 quick potency assay was plotted against qRT-PCR, immunostainings, and pellet diameter area of day 35 3D cultures (Fig. 6).
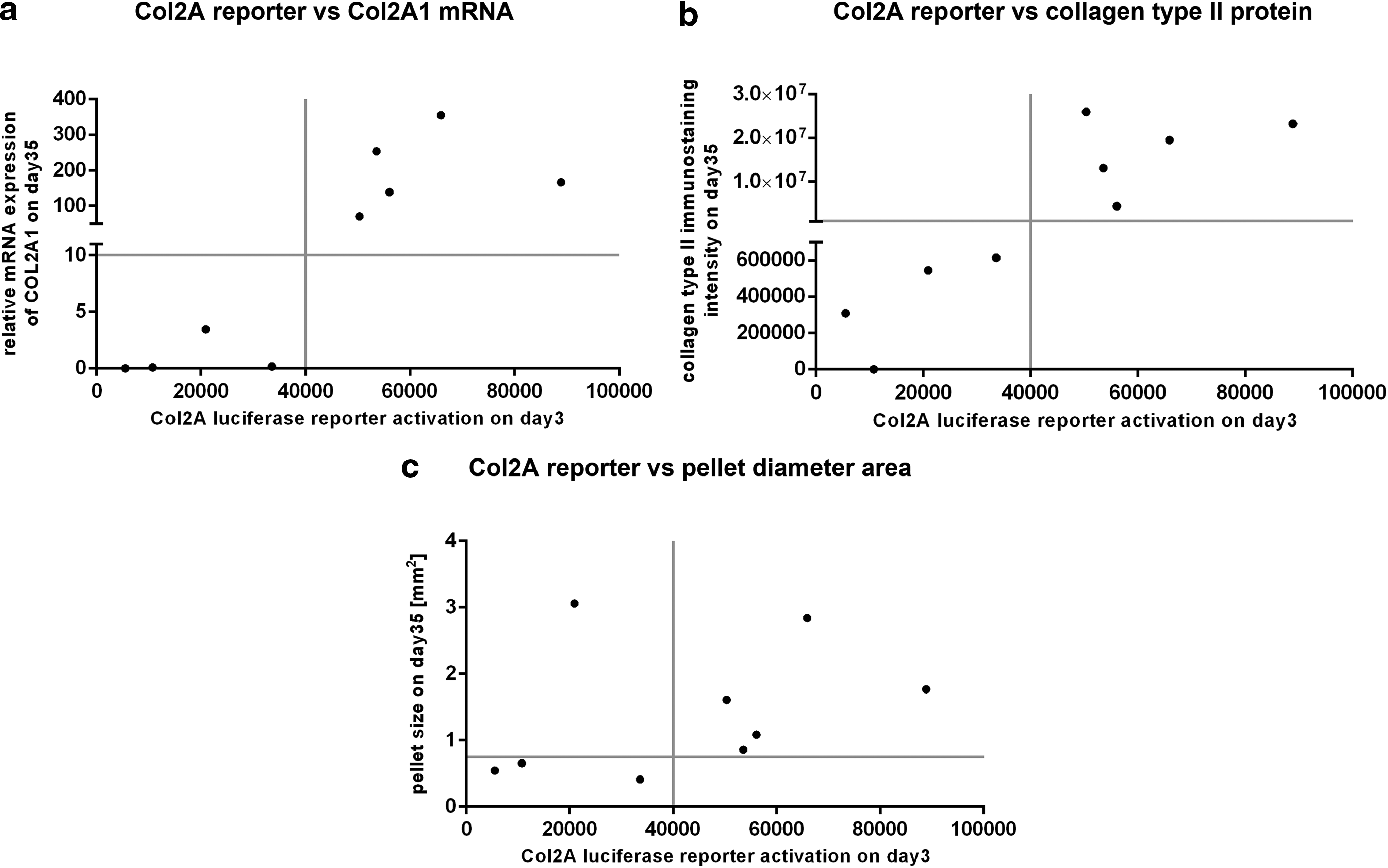
Relationship of Col2A luciferase reporter activation to Col2A1 mRNA expression, collagen type II immunostaining, and pellet diameter area. Data of single donors are displayed with luciferase reporter activation on the x-axis, and mRNA expression
By comparing values of Col2A luciferase reporter activation to Col2A1 mRNA expression (Fig. 6a) as well as to collagen type II immunostaining intensity (Fig. 6b), the determined good donors 3, 4, 5, 8, 9 and bad donors 1, 2, 6, 7 could clearly be identified. Regarding the relationship between Col2A luciferase reporter activation and the pellet diameter area (Fig. 6c), there is no clear cutoff line to identify donor quality, which may, however, partly be due to the low correlation of pellet size and the quality of chondrogenic differentiation.13,14
Discussion
In the current study, we developed a quick potency assay for fast determination of chondrogenic differentiation status of ASC. ASC can be induced for collagen type II expression after extensive passaging, 15 but also directly after isolation from adipose tissue. 16 The expression of collagen type II is mainly regulated through the TGF-β pathway17,18; in a previous work, we could demonstrate that TGF-β and BMP-6 induce collagen type II expression in ASC, in a 3D culture system. 19 Also, other pathways such as MAPK and Wnt/β-catenin signaling have impact on collagen type II expression but have not been investigated in detail as the TGF-β pathway.20–24 To predict chondrogenic potential of ASC from different donors, Kang et al. measured activation of TGF-β receptor expression. The chondrogenic potential was increased with a bicistronic vector system containing TGF-β receptors. 25 Duryagina et al. developed a new reporter system based on the Gaussia luciferase reporter to monitor HSC-supportive proteins in human MSC. 26
The advantages of assessing reporter genes with bioluminescent assays are the high sensitivity and the nondestructive and rapid application compared to protein and RNA analysis. 27 Based on a previous study, where we designed a reporter construct for detection of osteogenic differentiation, 9 we used in this study a novel reporter for chondrogenic differentiation. Cells from four individuals demonstrated a value for Col2A1 relative mRNA expression lower than five, which confirmed the weak Alcian blue and absent immunohistological staining for collagen type II. These donors were regarded as bad donors. The remaining five donors expressed Col2A1 in the range of 70–350 and showed intense Alcian blue and collagen type II expression on protein level. These donors were regarded as good donors. The pellet diameter area of each good and bad donor corroborate these results, except donor 2, which is in the range of the good donors but still negative for Alcian blue and collagen type II staining. This is slightly different to a study of Hennig et al., 7 who investigated the chondrogenic differentiation potential of nine donors treated with TGF-β and BMP-6 resulting in seven of nine donors positive for collagen type II immunohistochemistry and all investigated samples positive for Col2A1 gene expression. They also found that low-potency donors lack the expression of TGF-β receptor I, which could be partly overcome by coinduction with TGF-β and BMP-6. 7 We have identified that 20–50% are low-potency donors (with low or no collagen type II expression); also, in the presence of BMP-6, there is need for further analysis to find out differences between good and bad donors.
Our determined good and bad donors were evaluated with our reporter gene assay using the identical differentiation condition and compared to collagen type II expression on mRNA and protein level on day 35. To our knowledge, we were the first who showed transfection of chondrogenic micromass pellet cultures. The results of the quick potency assay measured for Metridia luciferase in the supernatant showed an increase of Col2A promoter activation for all donors from 1 to 3 days with significant higher values of the good donors on day 3. Even the values normalized for Renilla luciferase activity were significantly higher for the good donors. Notably, the Col2A reporter activation normalized against Renilla demonstrated higher values in each of the good donors compared to each single bad donor. These results corroborate the analyses of the histological stainings and qRT-PCR after 35 days with five good donors and four bad donors. At the single-donor level, Col2A luciferase reporter activation measured on day 3 was comparable to the readout from day 35 pellet culture Col2A1 mRNA levels and immunostaining for collagen type II and Alcian blue. We could hence identify the four donors with the weakest potential for chondrogenesis resulting in no deposition of collagen 2 in the pellets after 5 weeks. This result based on Col2A promoter activity could be achieved, although Col2A is a late marker and Col2A1 mRNA levels on day 3 were indeed negative.
mRNA levels of the early marker Sox9 could be detected in all nine donors already on day 3. However, expression levels were extremely low and did not allow identification of the previously determined donors obtained by Col2A1 mRNA level and immunostainings on day 35. Hence, Sox9 expression on day 3 was not predictive for chondrogenic potency at an early time point.
Although 3D micromass pellet culture is the prerequisite for induction of chondrogenic differentiation, we simplified this assay and changed to 2D culture. Surprisingly, also under 2D culture conditions, the Col2A reporter activity increased over 3 days for all donors with a significant difference on day 3. The change from 3D to 2D contributed toward further improvement by saving time (2 days less, since no pellet formation was necessary) and cell number (1/10), while maintaining the functionality of the assay. Moreover, under 2D conditions, no autofluorescence was visible, which interferes with the luciferase reporter signal. Considering absolute Metridia (Figs. 4a and 5a) and Renilla (Figs. 4b and 5b) values, obviously 2D culture conditions give higher signals compared to 3D culture conditions. This might be due to inferior transfection or cell lysis efficiency in 3D micromass pellets compared to monolayer cultures. However, Metridia values normalized against Renilla are higher under 3D (Fig. 4c, e) than under 2D (Fig. 5c, e) culture conditions, which confirms the fact that ASC cultured in micromass pellets are more committed to the chondrogenic lineage compared to 2D culture. The results demonstrate that a determination between good and bad donors is possible under both 3D and 2D conditions. This implies that the functionality and quality of the assay are maintained, combined with the advantages of a quick potency assay.
In this study, we have established a proof of principle concept for the analysis of donor quality in terms of chondrogenic differentiation potential. Based on this study, the identification of donor material with poor chondrogenic differentiation potential should allow excluding these cells at early time points. Before an implementation as a valuable current good manufacturing practice quality control could be established in the future, further careful evaluation of the cutoff point with a high number of donors would be required.
Footnotes
Acknowledgments
We thank Ludwig Aigner from Paracelsus Medical University (PMU) in Salzburg, Austria, for providing the Renilla control plasmid. This work was funded by a grant from the Austrian Research Promotion Agency (FFG) (Bridge1 programme, grant No.846062).
Disclosure Statement
The authors declare that they have no competing interests. The authors alone are responsible for the content and writing of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.