
Abstract
Women younger than 40 years may face early menopause because of premature ovarian failure (POF). The cause of POF can be idiopathic or iatrogenic, especially the cancer-induced oophorectomy and chemo- or radiation therapy. The current treatments, including hormone replacement therapy (HRT) and cryopreservation techniques, have increased risk of ovarian cancer and may reintroduce malignant cells after autografting. Decellularization technique has been regarded as a novel regenerative medicine strategy for organ replacement, wherein the living cells of an organ are removed, leaving the extracellular matrix (ECM) for cellular seeding. This study aimed to produce a xenogeneic decellularized ovary (D-ovary) scaffold as a platform for ovary regeneration and transplantation. We have developed a novel decellularization protocol for porcine ovary by treatment with physical, chemical, and enzymatic methods. Using hematoxylin and eosin (H&E) staining, DAPI staining, scanning electron microscopy (SEM), and quantitative analysis, this approach proved effective in removing cellular components and preserving ECM. Furthermore, the results of biological safety evaluation demonstrated that the D-ovary tissues were noncytotoxic for rat ovarian cells in vitro and caused only a minimal immunogenic response in vivo. In addition, the D-ovary tissues successfully supported rat granulosa cell penetration ex vivo and showed an improvement in estradiol (E2) hormone secretion.
Introduction
O
For patients wishing to maintain fertility, cryopreservation of ovarian tissues and allogeneic transplantation could be a solution.9–12 Although cryopreservation techniques have achieved success and given birth to more than 36 babies, the risk of reintroducing malignant cells from stromal tissue into transplantation sites cannot be ignored. 13 Moreover, a limited life span of 4–5 years has been reported for retransplanted ovarian tissues. 13 Thus, this research seeks a long-term, hypoallergenic, and safe therapy.
In the past decade, interest has grown considerably in decellularization techniques, wherein the living cells of an organ are removed, leaving the extracellular matrix (ECM) to be used for regenerative medicine. 14 The lack of cells and low immunogenicity of decellularized ECM make it ideal for use in allotransplantation and even xenotransplantation. In 2015, decellularized ECM was successfully harvested from a bovine ovary for the first time. 15 The study provided the initial step of creating an artificial ovary and introduced a novel therapy for ovariectomized mice. However, long-term sodium dodecyl sulfate (SDS) treatment in the study has been reported to have a strong damaging effect on the ultrastructure of native tissue and be cytotoxic.16,17 Therefore, we believed that a novel strategy with shortened SDS treatment and testing of the biocompatibility needed to be discussed. Furthermore, we believed it was necessary to further investigate whether ECM affects cell migration, proliferation, and other biological functions. Therefore, we proposed an alternative approach and evaluated the effects of decellularized tissues on cell bioactivity.
Materials and Methods
Study design
All animal protocols were approved by the Animal Experimental Center of Wenzhou Medical University. Native ovary (N-ovary) tissues were decellularized by sequential treatment with physical, chemical, and enzymatic methods. Figure 1 shows the overview of our study design.
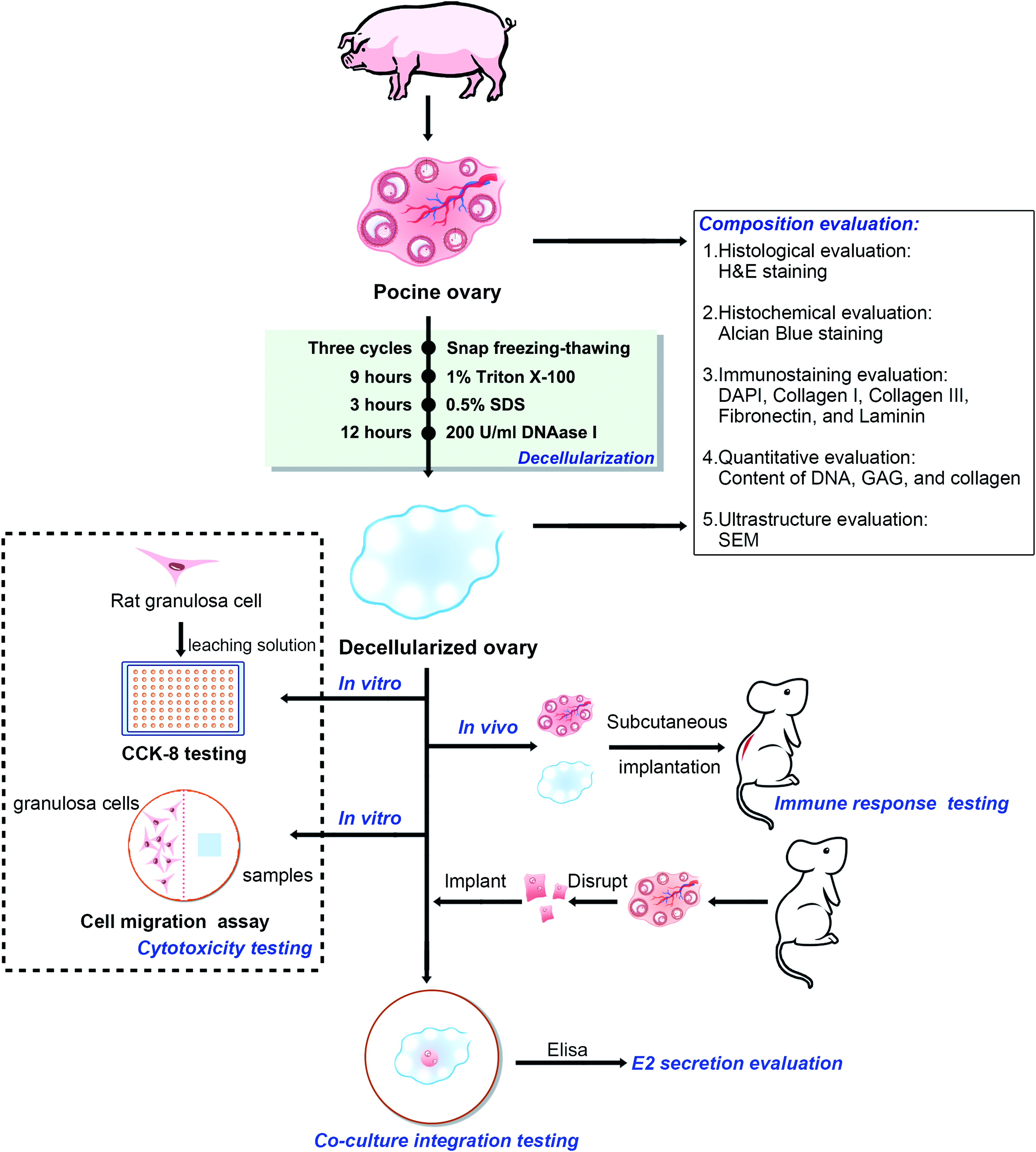
Flow diagram of study design. Color images available online at
Ovary tissue harvest
Ovary samples for decellularization were collected from female Bama miniature pigs weighing 10–15 kg, at a local slaughterhouse. After isolation of the ovary from connective tissue and fat, the ovary was washed thrice with phosphate-buffered saline (PBS) solution and preserved at −80°C for further decellularization.
Decellularization process
First, the ovary tissues were treated with freeze–thaw cycles at −80°C and 37°C thrice and sliced into 1.5 mm thick pieces. After freeze–thaw cycles, the samples were washed in deionized water with 1% Triton X-100 (Sigma) for 9 h and 0.5% SDS (Sigma) for 3 h at room temperature with agitation (100 rpm). Then, treated samples were washed with deionized water for 12 h. Finally, 200 U/mL DNase I (Sigma) in PBS was applied to the samples at 37°C for 12 h to further minimize residual DNA. Digested samples were washed again with deionized water for 4 h. Before each testing, the samples were sterilized with 3% peracetic acid.
DNA and collagen content analysis
To investigate the amount of residual nuclear material after the decellularization process, DNA quantification was performed on both decellularized ovary (D-ovary) (n = 7) and N-ovary samples (n = 7) using a genomic DNA extraction kit (TaKaRa) according to the instructions. The extracted DNA materials were analyzed according to the absorbance obtained with a spectrophotometer (Bio-Rad).
The collagen content was evaluated based on the hydroxyproline (HYP) content determined using an HYP assay kit (Keygen), according to the manufacturer's protocol. The D-ovary (n = 7) and N-ovary (n = 7) samples were lyophilized to obtain their dry weight. Then, the amount of HYP was determined from a calibration curve calculated using data from standard HYP samples (Sigma). The ratio of total collagen/HYP content is 7.2:1.
Histological and immunostaining
Histological and immunological staining were performed on D-ovary tissues and native samples to observe the residual components remaining after the process, if any. Samples were fixed in 10% formalin solution in PBS at room temperature, dehydrated in ethanol, embedded in paraffin wax, and sectioned to 5-μm thickness.
For histological analysis, sections were stained with hematoxylin and eosin (H&E) to visualize the cellular components and ECM structure in D-ovary samples. Alcian Blue staining was used to qualitatively evaluate the glycosaminoglycan (GAG).
Immunohistochemical staining was carried out on decellularized and N-ovary samples to observe the presence of collagen types I and III, fibronectin, and laminin. First, the sections were treated with a citrate buffer for antigen retrieval at 95°C for 10 min and cooled at room temperature for 30 min. After blocking endogenous peroxidases and nonspecific binding, sections were incubated with primary antibodies in a washing buffer for 12 h at 4°C. Then, tissue slides were washed thrice with PBS, and an appropriate secondary antibody was added. Finally, the sections were washed in PBS, treated with streptavidin–horseradish peroxidase complex and diaminobenzidine (DAB) solution, and counterstained with hematoxylin. Furthermore, both D-ovary and N-ovary sections were deparaffinized, rehydrated, and stained by DAPI (Sigma) to show the presence of residual nuclei components.
The primary antibodies were against collagen type I (Santa Cruz Biotech), collagen type III (Santa Cruz Biotech), fibronectin (Santa Cruz Biotech), and laminin (Santa Cruz Biotech) and the secondary antibody used was goat anti-mouse IgG1 (Abcam, Cambridge, MA) at a dilution of 1:100. Images of the sections were acquired by light microscopy, using a Nikon ECLIPSE 80i microscope (Nikon).
Scanning electron microscopy
To observe damage to the microarchitecture of the tissue scaffold after the decellularization process, scanning electron microscopy (SEM) was performed for D-ovary and N-ovary tissues. After pretreatment, the samples were coated with gold–palladium and observed by SEM (EM-3200; KYKY). The detailed description of pretreatment process can be obtained in our previous studies. 18
Isolation of rat granulosa cells
Immature female Sprague-Dawley rats (21–24 days old) were injected with 40 IU pregnant mare serum gonadotropin (PMSG) (ProSpec) 48 h before isolation. Granulosa cells were harvested from ovaries by puncturing follicles with a 25-gauge hypodermic needle under a dissecting microscope and purification with a discontinuous Percoll (Sigma) gradient as previously described. 19
Cytotoxicity test of D-ovary tissue
The Cell Counting Kit-8 (CCK-8) test and cell migration assay (conducted by scratch wound assay) were performed to evaluate cytotoxicity and cellular bioactivity of the D-ovary tissues. In the CCK-8 test, D-ovary samples (10 × 10 × 1.5 mm) were incubated in standard Dulbecco's modified Eagle's medium (DMEM)/F12 medium for 48 h in a humidified 37°C/5% CO2 incubator, and the medium was collected as the leaching solution (one sample per milliliter of medium). Subsequently, granulosa cells were seeded in 96-well cell culture plates with DMEM/F12 at a concentration of 5 × 103 cells/well for 24 h. The medium was replaced by the leaching solution (25%, 50%, and 100%) and the standard medium (control group). After 1, 3, and 5 days of culture, CCK-8 solution was added to each well, and optical density (OD450) was measured with a spectrophotometer (Bio-Rad) to determine the metabolic activity level of the cells.
In the cell migration assay, granulosa cells were seeded into 12-well culture plates and incubated in 15% fetal bovine serum (FBS) DMEM/F12 for 24 h. After preculture, a mechanical wound was created to remove cells in the right-side part of each well and the culture medium was replaced by serum-free DMEM/F12. Subsequently, D-ovary samples (6 × 6 × 1.5 mm) were placed into the right-side parts of experimental groups and cocultured with granulosa cells. Following coculture, cell cultures were fixed in 10% formalin solution for 30 min at room temperature, washed in PBS, and stained with 1% crystal violet for 30 min. Cell migration was observed and captured under a Nikon Ts100 microscope (Nikon).
Immune response to D-ovary in vivo
For the biocompatibility test, 10 male rats underwent subcutaneous implantation. The D-ovary samples (n = 10) and N-ovary (n = 10) samples (6 × 6 × 1.5 mm) were surgically implanted into the backs of rats (each rat with one each of the decellularized and native samples). First, each rat was anesthetized by an injection of 10% chloral hydrate (5 mL/kg) intravenously. After shaving the operating sites, the skin was cleaned with 10% povidone iodine. Small incisions (7 mm) were made on each side of the dorsal midline of rats and the samples were implanted. After closing the incisions with synthetic absorbable sutures, investigators observed the wounds daily. The implanted samples were harvested during week 2 and 4 (five rats at each time). Histological, immunohistochemical, and quantitative evaluation were performed to show the recruitment of host cells and immune cells in the ovary implants. Immunohistochemical staining was performed using antibodies against CD68 (Novus Biologicals, Littleton, CO), CD86 (Novus Biologicals), and CD3 (Novus Biologicals). Quantification of immune cell density was achieved using NIH Image J software.
Evaluation of the integration of ovarian cells with D-ovary tissue ex vivo and their secretion of E2 hormone
Under laparotomy, oophorectomy was performed on female Sprague-Dawley rats, each of which was given an intraperitoneal injection of 40 IU PMSG (ProSpec) 48 h before surgery. Following removal, ovaries were immediately immersed in DMEM/F12, while connective tissue and fat were eliminated under a dissecting microscope (Nikon). Next, ovaries were washed in PBS thrice and mildly mechanically disrupted into small fragments (2 × 2 × 2 mm). The fragments were immersed in dispersal solution at 37°C for 10 min (0.25% trypsin and Na-EDTA in DMEM/F12). A small hole was drilled into the decellularized sample and the ovarian fragment was implanted in this hole, in addition to being cultured alone as a control environment.
To evaluate the integration of ovarian cells into the D-ovary samples, the complex of ovarian tissue and decellularized samples was cocultured in DMEM/F12 containing 15% FBS and incubated at 37°C in an atmosphere of 5% CO2. Two days before harvest, the medium was switched to serum-free DMEM/F12 and treated with the indicated hormones/compounds in preparation for measuring E2 synthesis as previously described. 20 After 5, 7, and 9 days of coculture, the samples were analyzed by immunohistochemical staining. To determine whether ovarian cells adhered, migrated, and proliferated into D-ovary samples, we used a stain for FSHR, a granulosa cells marker. In addition, the E2 concentrations in the culture supernatant were assayed with an enzyme immunoassay test kit (Boyun Biotech) according to the manufacturer's protocol.
Statistical analyses
One-way analysis of variance and Student's t-test, or Mann–Whitney U test was used to compare the differences in the content of DNA, collagen, E2, OD value, and immune results among different treatment groups. All p-values were two sided, and a p-value of <0.05 was considered statistically significant. Analyses were performed in SPSS version 20.0 software (IBM, Armonk, NY).
Results
Decellularization eliminates cellular components from the ovary scaffold
The first goal of a decellularization process is to effectively remove cellular components from the scaffold. After being processed with detergent solution, the color of the ovary samples turned from red to white, revealing a change in the cellular components (Fig. 2A, B). To investigate residual nuclear material, H&E staining was performed. As shown in Figure 2D and F, D-ovary tissues showed an absence of basophilic staining and a large area of eosinophilic staining both in cortex and medulla tissues. In contrast, both the basophilic and eosinophilic staining were performed in N-ovary tissues (Fig. 2C, E). The H&E results suggested that there were no visible bands of DNA after decellularization. To further assess the elimination of nuclear materials from ovary samples, we stained the samples with DAPI, the results of which were consistent with the results of H&E staining (Fig. 2G–J). Then, we quantitatively analyzed the DNA content of decellularized tissues. The results suggested a significant decrease in DNA content from D-ovary samples compared with that from native ones (701.62 ± 469.28 ng/mg vs. 20.92 ± 4.56 ng/mg dry weight, p < 0.05) (Fig. 2K).
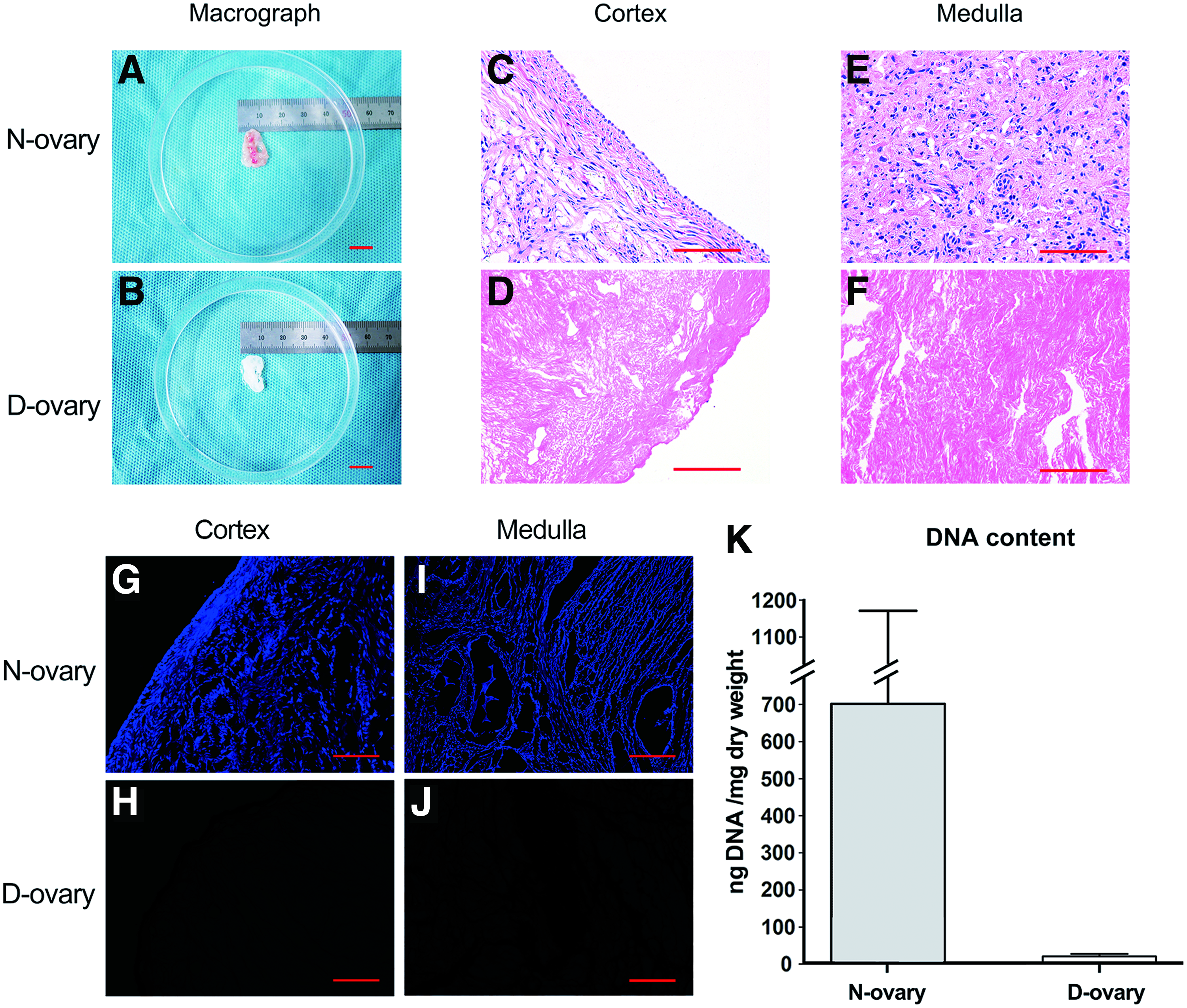
Micrograph
Decellularization preserves the ECM components and microarchitecture of D-ovary tissue
Next, we investigated the composition and microarchitecture of decellularized ECM. Alcian Blue staining was performed to visualize GAG distribution in the ECM. Figure 3A and B show that both cortex and medulla layers in D-ovary samples were rich in GAGs. However, our quantitative analysis revealed a reduction in the GAG content of D-ovary compared to N-ovary samples (85.1 ± 12.5 μg/mg vs. 44.4 ± 17.8 μg/mg dry weight, p < 0.05) (Fig. 3A, B).
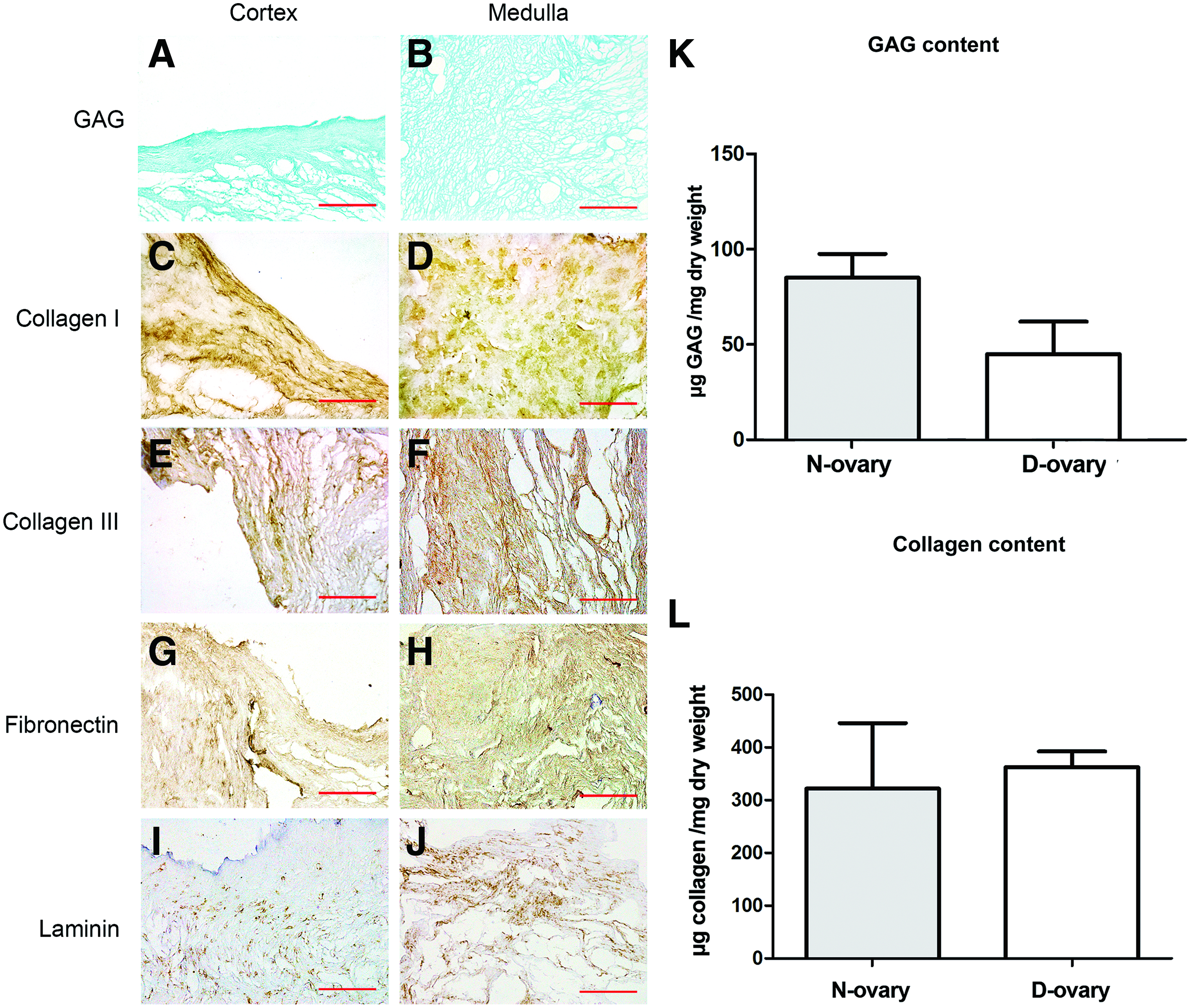
Composition of decellularized ECM in D-ovary samples using Alcian Blue and immunohistochemical staining
Immunohistochemical staining showed that collagen type I, collagen type III, fibronectin, and laminin were all identifiable (Supplementary Fig. S1; Supplementary Data are available online at
Next, we investigated the total collagen content based on HYP (Fig. 3L). There was no significant difference in the collagen content (321.8 ± 124.1 vs. 362.8 ± 29.7 μg/mg dry weight, p > 0.05), confirming the results of the immunohistochemical staining.
To qualitatively evaluate any destructive effects on the scaffold from the decellularization process, SEM was used to observe the microarchitecture of D-ovary samples. Figure 4A shows that ovarian cells were attached to the ovary (label “a”), while no visible cells can be found on the surface of D-ovary tissues (Fig. 4B), providing evidence supporting the effective removal of cellular components. The structure of the follicular cavity can be observed in Figure 4C and D (labels “c” and “b”) in native and processed samples, showing an empty space where it once contained follicles, granulosa cells, and theca cells. After the decellularization process, these cavities were still preserved; however, the pore walls presented a higher degree of porosity because of the lack of intact cells (labeled as “d” in Fig. 4D). Figure 4E and F showed a well-connected and oriented collagen network in both and there was no deformation or fiber breakage resulting from the decellularization process. These findings were consistent with the result from H&E staining and quantitative analysis, which showed that the collagen has not undergone obvious damage and significant loss.
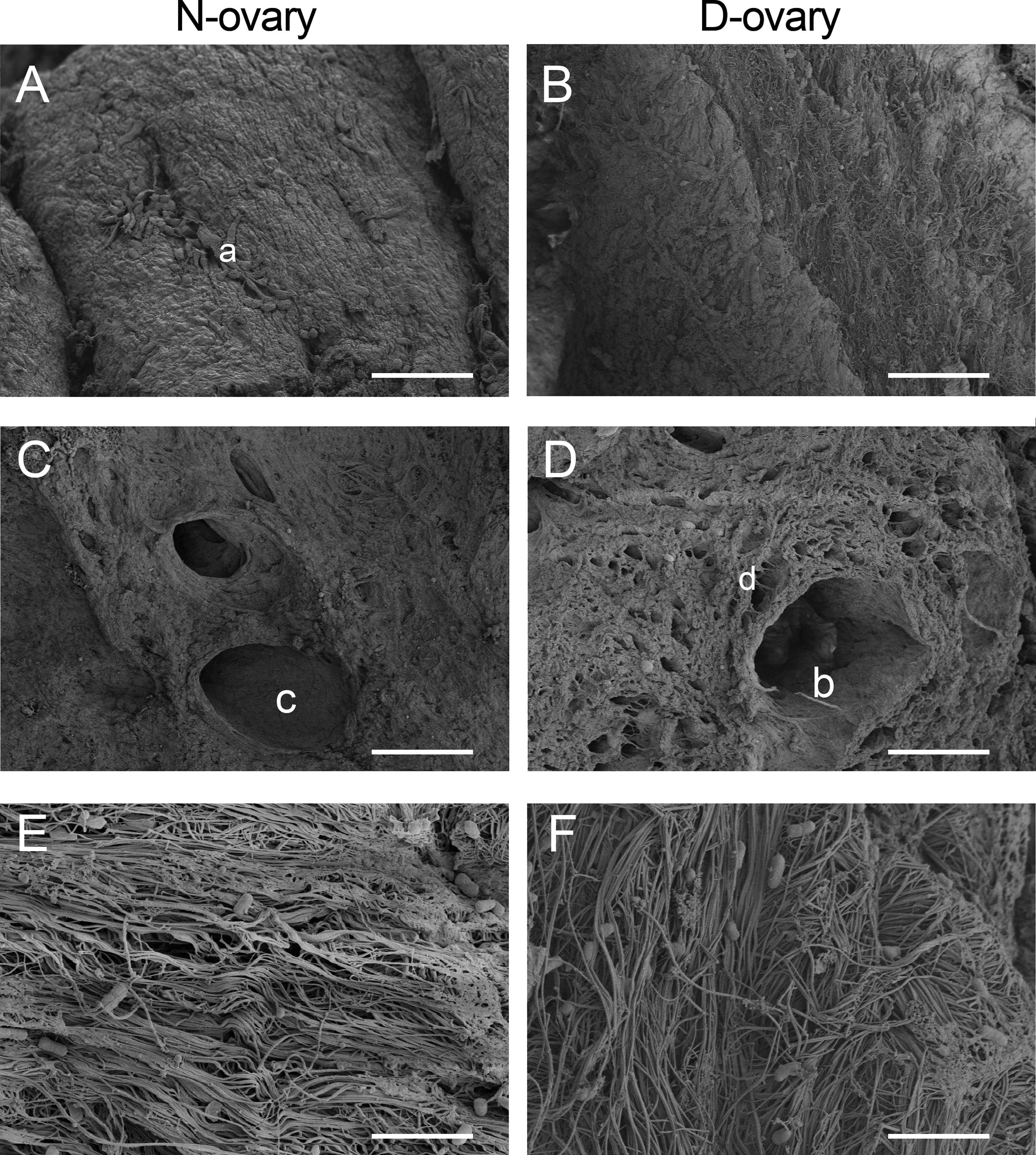
Microarchitecture of decellularized scaffold presented by scanning electron microscopy images. “a”: Ovarian cells in native scaffold; “b” and “c”: follicular cavity was observed both in native and decellularized scaffold; “d”: pore walls of D-ovary became porous because of the lack of intact cells.
D-ovary scaffolds show no cytotoxicity for ovarian cells in vitro
The cytotoxicity of residual chemicals in D-ovary tissues after the decellularization process may affect cell viability and migration. Therefore, the CCK-8 test and cell migration assay were performed in vitro. In CCK-8 test, the OD value measured cell proliferation and survival. After granulosa cells were treated with the leaching solution, the granulosa cells presented no significant difference in OD value, compared with cells treated with the standard medium (Fig. 5G). In the cell migration assay, granulosa cells were cocultured with D-ovary tissues in 12-well culture plates for 5, 7, and 9 days (Fig. 5D–F). Compared with the control group (Fig. 5A–C), the experimental groups showed more ovarian cells migrating across the red lines, indicating that D-ovary ECM improved cell migration.
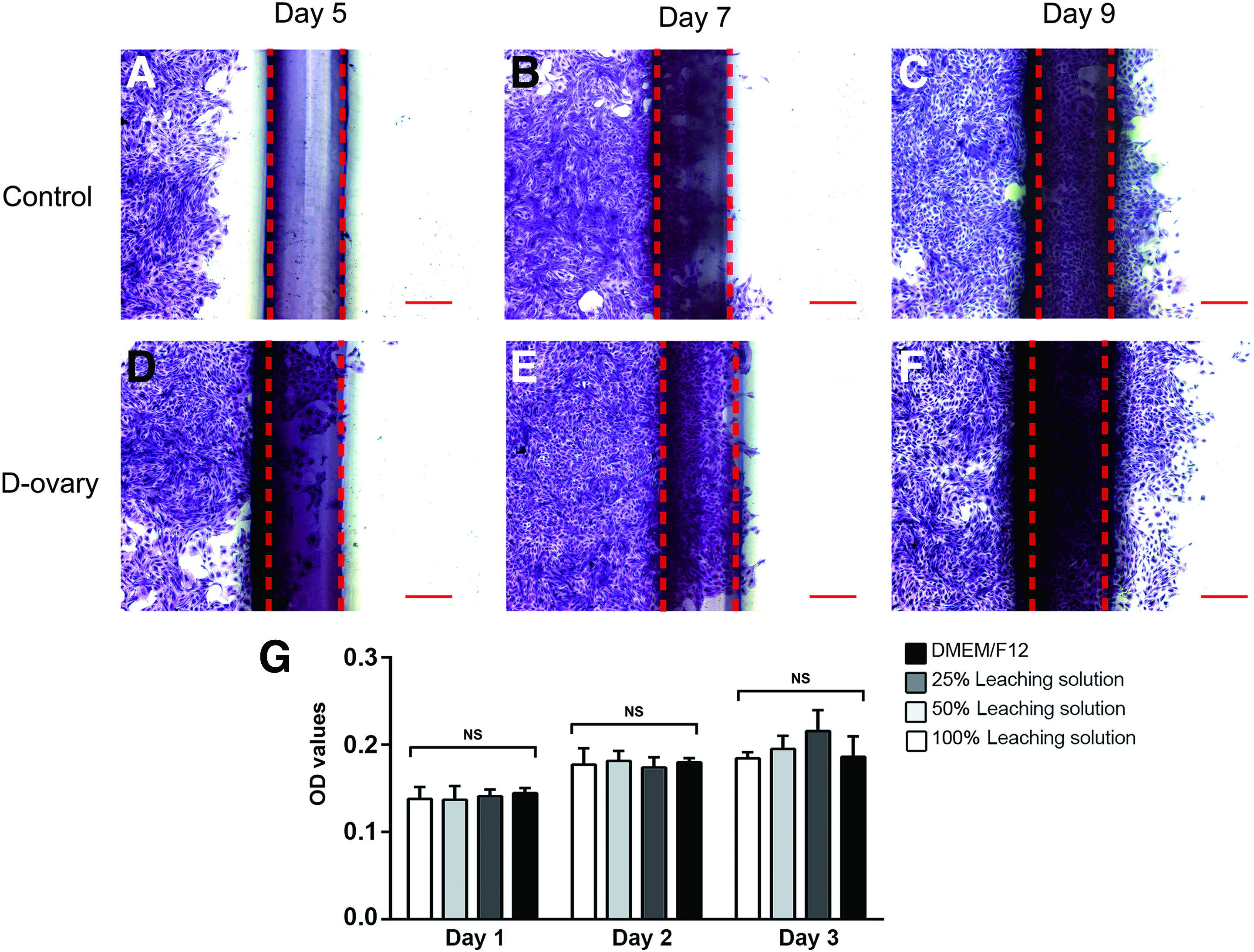
Cytotoxicity of D-ovary samples. The migration and metabolic activities of ovarian cells were measured using the cell migration assay
Decellularization alleviates immunological response in xenotransplantation
After testing cytotoxicity in vitro, the in vivo biocompatibility of D-ovary tissue was evaluated by xenogeneic subcutaneous implantation. The samples were implanted subcutaneously to assess host response after 2 and 4 weeks. All rats survived without complications during the 4 weeks of implantation. H&E staining showed that a large number of host cells were observed in N-ovary samples, while only a few cells were observed in D-ovary tissues (Fig. 6A–D). Furthermore, immunohistochemical staining was performed to identify the immune cells. Figures 6E–P show infiltration of CD68+, CD86+, and CD3+ cells after 2 and 4 weeks of implantation. We observed an exaggerated immune response in native samples after 2 weeks. In contrast, the decellularized samples presented only a mild immune response (43.40 ± 1.21 vs. 82.14 ± 2.79 cells × 10−5 μm3, p < 0.001 for CD68+; 25.29 ± 1.22 vs. 61.62 ± 7.63 cells × 10−5 μm3, p < 0.001 for CD86+; and 12.38 ± 1.77 vs. 49.78 ± 7.15 cells × 10−5 μm3, p < 0.001 for CD3+) (Fig. 7). After 4 weeks, the D-ovary samples still presented a lower density of immune cells compared with the native samples (47.37 ± 1.53 vs. 92.70 ± 1.98 cells × 10−5 μm3, p < 0.001 for CD68+; 30.76 ± 5.41 vs. 80.05 ± 1.50 cells × 10−5 μm3, p < 0.001 for CD86+; and 14.15 ± 0.22 vs. 67.25 ± 12.09 cells × 10−5 μm3, p = 0.001 for CD3+). Moreover, no additional immunological responses by CD86+ and CD3+ cells could be observed in D-ovary samples at the end of 4 weeks compared those after 2 weeks (25.29 ± 1.22 vs. 30.76 ± 5.41 cells × 10−5 μm3, p = 0.086 for CD86+ and 12.38 ± 1.77 vs. 14.15 ± 0.22 cells × 10−5 μm3, p = 0.089 for CD3+). Taken together, the data presented the improved biocompatibility of the D-ovary samples in xenogeneic subcutaneous implantation.

H&E and immunohistochemical staining show the immunological response of the host to native and decellularized ECM implanted subcutaneously after 2 and 4 weeks. Scale bar 100 μm. Color images available online at
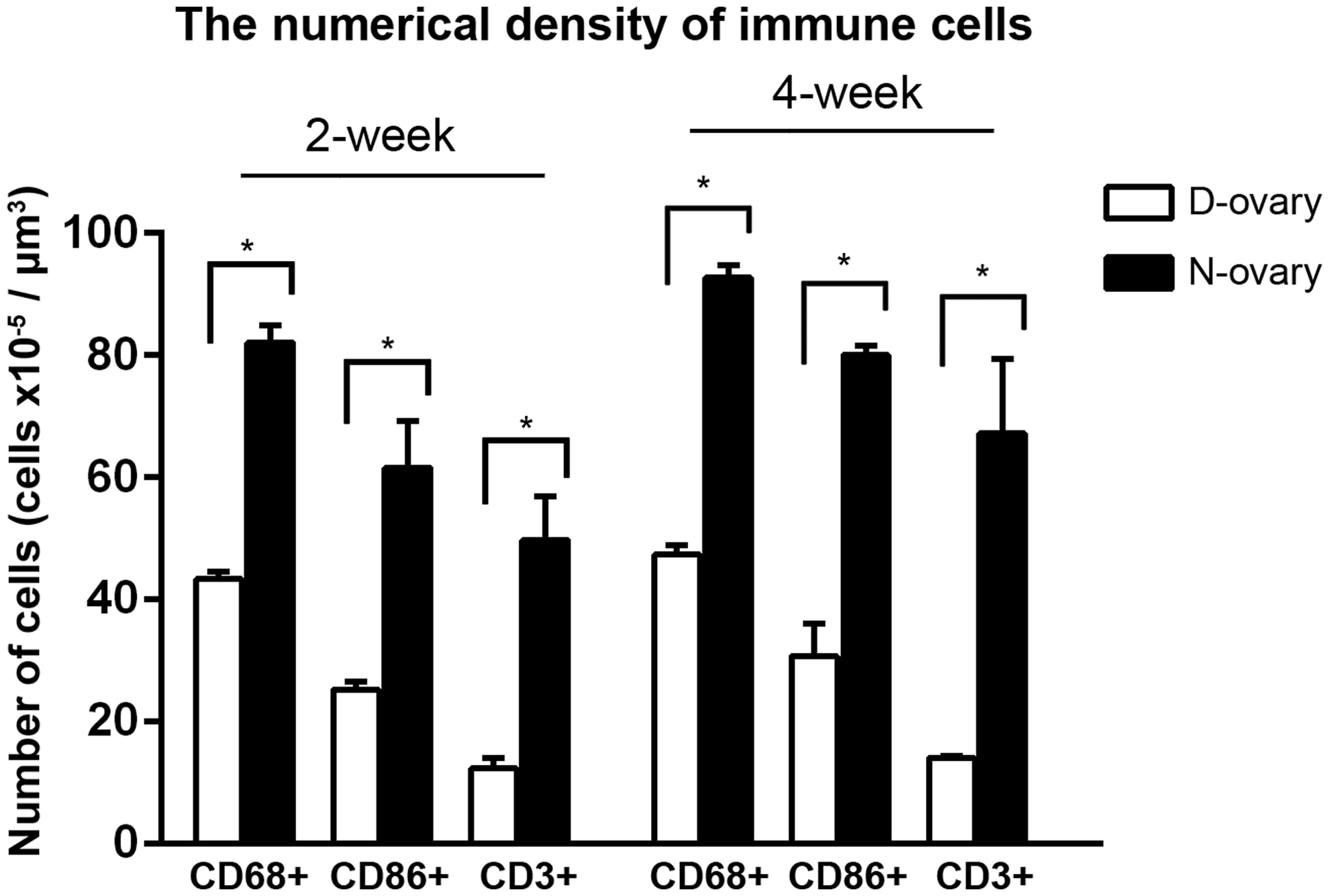
The numerical density of CD68+ (pan-macrophage/monocyte marker), CD86+ (M1 macrophage marker), and CD3+ (T lymphocyte marker) immune cells in D-ovary and N-ovary samples at 2 and 4 weeks. *p < 0.05.
The D-ovary scaffold supports the migration and secretion functions of ovarian cells ex vivo
In an attempt to demonstrate the regenerative potential of the D-ovary scaffold, xenogeneic ovary tissues were implanted into D-ovary samples and cocultured in DMEM/F12. After the culture period, samples were harvested for immunohistochemical staining. Figure 8 shows the granulosa cells stained with the specific marker FSHR. The red dotted lines indicated the border of N-ovary tissues and arrows indicated the direction of cell migration. After 6 days of culturing, the cells started to penetrate into the D-ovary scaffold (Fig. 8E). As shown in Figure 8E, a few granulosa cells were scattered around the red line in the porcine scaffold. At days 8 and 10, there were many more cells observed far from the N-ovary tissues (Fig. 8F, G). Finally, the granulosa cells seemed to aggregate together in the acellular scaffold after 12 days of culture (Fig. 8H).

Integration of granulosa cells into D-ovary scaffold ex vivo
Furthermore, E2 expression levels in culture media were also measured. Two days before the test, the culture medium was replaced with serum-free DMEM/F12, and isolated ovary tissues were utilized as a control group. As shown in Figure 8I, the ovary tissue implanted in decellularized ECM secreted more E2 than the isolated ovary tissue (enzyme immunoassay test). Overall, our study showed that D-ovary tissues could support the adhesion, migration, and proliferation of granulosa cells, and sustained E2 secretion functions.
Discussion
Conditions such as cancer, organ failure, and chemo- and radiation therapy can result in whole-organ resection and call for transplantation. To address these problems, researchers have been developing a decellularization technique that removes cellular components from ECM to build a regenerative platform for organ replacement.21–24 In 2015, the first D-ovary platform was successfully obtained and it partly restored endocrine functions in a mouse model. 15 However, some limitations existed in that method. First, the long SDS process has a strong damaging effect was demonstrated on the ultrastructure of the native tissue.16,17 Therefore, an alternative approach with a reduction in SDS incubation time could improve the preservation of ECM. After decellularization, the authors analyzed the ECM compositions, but did not quantify the GAG content, which is closely associated with bioactive factors in ECM, but is sensitive to decellularization solution. Furthermore, residual chemicals and cellular antigens can cause serious adverse effects in organ recipients. Therefore, the biological compatibility of decellularized tissues must be discussed in detail. In addition, it is also necessary to investigate whether the D-ovary scaffold can improve cell proliferation, migration, and endocrine function. Thus, we redesigned the decellularization protocol and further investigated the biosafety and effects of the decellularized scaffold on cell bioactivity.
According to a previous study, preprocessing with Triton X-100 could destabilize the membranes of tissues, which resulted in better penetration of SDS. 25 In addition, Triton X-100 caused less damage to the scaffold microstructure compared with SDS. 22 Thus, we improved the decellularization strategy by adding Triton X-100 solution and shortening the duration of the SDS incubation. Moreover, we further shortened the chemical processing time and improved the elimination of cellular components by adding DNase digestion and freeze–thaw steps to our process. The physical process of freezing and thawing can induce rapid lysis of cells and promote chemical solution infiltration. The final process of DNase digestion can further reduce residual DNA fragments. Taken together, the improved decellularization process in our study is to sequentially treat ovary samples with freeze–thaw cycles, Triton X-100, SDS solution, and DNase I.
The first goal of decellularization was to successfully remove cellular components from the ECM. Removal of allogeneic or xenogeneic cellular content is necessary because cellular antigens can induce adverse immune responses.26–29 However, because of the dense structure of ECM, it is impossible to completely remove all of the cellular components. Therefore, a DNA content of less than 50 ng/mg dry tissue weight is regarded as an objective criterion for effective elimination of cellular components. Our results meet this criterion, indicating suitability for transplantation. 25
Another challenge for decellularization techniques was to find the balance between effective removal of cellular components and retention of ECM materials. As mentioned above, detergents were destructive to ECM materials. The function of acellular ECM is not only to provide a three-dimensional skeleton for cell growth but also to play an essential role in various biological processes. For example, heparin sulfate of GAGs is the binding site for various growth factors that participate in physiological processes. 30 Moreover, there is evidence that ECM directly affects cellular differentiation, proliferation, and gene expression. 31 Our immunohistochemical examinations showed that ovary-derived ECM materials, including collagens I and III, fibronectin, and laminin, were well preserved both in the cortex and medulla of ovary tissues. However, a quantitative evaluation revealed a reduction of GAG content in D-ovary tissues. These findings indicated that GAGs were more susceptible to the destructive effects of the decellularization processes than other ECM components. Still, considering that abundant GAGs remained after the entire process was completed, we think that the decellularization protocol is acceptable.
Cytotoxicity and biocompatibility are important issues of recellularization techniques. In our protocol, we used chemical solutions to efficiently fabricate porcine decellularized ovaries. However, there is evidence that excessive residual SDS in ECM can be cytotoxic and inhibit cell infiltration.32–34 Thus, it is necessary to ensure that the decellularized scaffold is noncytotoxic after washing. The CCK-8 test measured cell proliferation and survival in D-ovary leaching solution compared with that in the standard medium. The results showed that D-ovary samples did not significantly inhibit the cell metabolic activity. Moreover, the cell migration assay investigated the effect of the D-ovary scaffold on cell migration. There was an enhancing effect of decellularized scaffold on cell migration, where more cells were observed migrating further away from the red lines in the cocultured group.
Despite the residual chemicals, xenogeneic tissues can affect the biocompatibility of decellularized tissues because of the host immune response. After subcutaneous implantation, the CD68 (pan-macrophage/monocyte marker), CD86 (M1 macrophage maker), and CD3 (T lymphocyte maker) cells showed significant infiltration into native tissues, whereas minimal immunological reaction was seen in decellularized tissues. Our findings are in agreement with those of Fishman et al.'s study, which demonstrated an inhibited M1 macrophage response and reduced proliferation of T cells against the decellularized scaffold. Furthermore, the authors even suggested that the decellularized scaffold exerts anti-inflammatory and immunosuppressive effects. 35 This may explain why the D-ovary group showed no increase in infiltration with M1 macrophages and CD3+ T-cells after 4 weeks compared with that after 2 weeks. The underlying mechanism may be partly ascribed to the reduction of MHC I and II molecules after decellularization.36,37
Ideally, the decellularized scaffold would well support cell adherence, penetration, and proliferation, and sustain E2 secretion functions. Building on a previous decellularization study with pancreatic cells, 38 we implanted rat ovary tissues into a porcine acellular scaffold to assess the potential of a recellularized graft. Interestingly, the granulosa cells not only successfully penetrated into the D-ovary scaffold but also presented improved endocrine functions. These findings were consistent with the result from the cell migration assay, which also showed that the decellularized scaffold had beneficial effects on cellular bioactivity. We suspected that the success might be partly associated with the high-level GAG content preserved from the ECM. As aforementioned, the GAG component provides binding sites for many bioactive factors (such as VEGF, bFGF, and PDGF), and Yang et al. proved that GAG was a determinant for the preservation of endogenous and exogenous bioactive factors in their decellularization experiment. 39 The high retention of GAG revealed that abundant bioactive factors may exist in D-ovary tissues. The researchers have suggested that these factors not only have an effect on cell attachment, proliferation, and differentiation but also play a crucial role in ovarian biology.39,40 Therefore, the coculture system provided extra growth factors promoting the cells' bioactivity and secretion functions.
There are, however, some limitations of our study. First, our study analyzed endocrine, rather reproductive, function in decellularized ovarian samples. We consider the production of sex hormone as the initial step in the restoration of fertility. Second, the results showed endocrine function and the potential for regeneration ex vivo; however, continuing studies are needed in vivo to confirm these conclusions. Third, our protocol was based on sliced ovary pieces instead of natural ovary. Future research directions will focus on decellularization and reconstruction of whole organ. Fourth, we used matured ovarian cells to evaluate the possibility of organ regeneration, which may have several limitations in clinical application. Seeding induced pluripotent stem cells (iPSC) into a decellularized platform may solve this problem and it has already been reported in the decellularized kidney, liver, and heart.29,41,42
Conclusion
In summary, we have developed a novel decellularization protocol for xenogeneic ovary regeneration by treatment of the tissues with physical, chemical, and enzymatic decellularization methods. This approach proved effective in removing cellular components and preserving ECM. Our evaluation of biological safety suggested that the scaffold was noncytotoxic for rat ovarian cells and caused only a mild host response in xenogeneic subcutaneous implantation. Furthermore, the D-ovary tissues successfully supported cell penetration and enabled an improvement in E2 hormone secretion.
Footnotes
Acknowledgment
This study was supported by the Zhejiang Provincial Natural Science Foundation of China (Y14H070034).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.