
Abstract
The aim of this study was to establish an embryo culture system using collagen gel attached with vascular endothelial growth factor (VEGF) derived from interleukin-1 beta (IL-1β)-treated endometrial tissues from pigs. Endometria were separated from the porcine uterus at the follicular phase of the estrous cycle and were cultured with IL-1β. The collagen gels coincubated with IL-1β-treated endometria (C, without endometrial tissue; CE, with endometrial tissue; and CEI, IL-1β-treated endometrial tissue) were used for embryo culture. We found that, compared with the comparable figures in the control group, prostaglandin synthase-2 (PTGS-2) mRNA was increased in IL-1β-treated endometrial tissue (p < 0.05). The VEGF protein was not observed in collagen gel coincubated without endometrial tissue (C); however, it was detected in collagen gels coincubated with endometrial tissue (CE and CEI). The embryo cleavage rates and blastocyst formation did not differ among the treatment groups. The proportion of blastocysts did not differ among the groups. However, the number of blastocyst cells was significantly (p < 0.05) higher in the CEI group than in the other groups. These results clarify the effects of the intrauterine environment on preimplantation embryos and may be useful in research on the effects of extracellular matrix- and cytokine-treated endometrial tissue on embryo development.
Introduction
I
Generally, a mature oocyte is fertilized with sperm in the ampulla of the oviduct. When porcine embryos are at the 8–16 cell stage, the fertilized egg (embryo) is transported from the ampulla of the oviduct to the uterus. 7 At this time, the corpus luteum is formed in the ovulation location of ovary, and the thickness of the endometrial tissue and the rate of angiogenesis increase compared with during the preovulatory phase, which is defined as luteal phase of estrous cycle.8,9 The endometrial tissue of luteal phase secretes various growth factors, sex hormones, and cytokines important for embryonic development and implantation. 10
Interleukin-1 beta (IL-1β) is a member of the interleukin 1 family of cytokines, which is an important mediator of the inflammatory response, and is involved in cellular activities, such as cell proliferation, differentiation, and apoptosis. 11 The process by which the endometrial tissue secretes IL-1β changes during the estrous cycle, which triggers prostaglandins (PG) synthesis in pigs.12,13 The PG in endometrial tissue play an important role in the regulation of estrous cycle and implantation in animals. 14 The PG biosynthesis occurs from arachidonic acid by prostaglandin synthase-2 (PTGS-2), also known as cyclooxygenase-2 (COX-2). 15 Recent in vivo and in vitro studies have investigated that PTGS-2 expression is associated with increasing of cellular proliferation and angiogenic factor,16,17 practically PTGS-2 mRNA expression was highly upregulated on days 10–14 of estrous cycle in porcine endometrium. 18 Thus, understanding the regulation of endometrial physiology by IL-1β during embryonic development is necessary to understand the mechanism that mediates between the intrauterine environment and the porcine embryo during the luteal phase.
The extracellular matrix (ECM) is essential for maintaining tissue structure, and collagen type I is a major component of endometrial tissue. 19 Cellmatrix Type I-A is used in tissue engineering,20,21 and three-dimensional (3D) culture systems, 22 several studies have established 3D endometrial cancer and fibroblast cell coculture systems based on collagen gel, 23 and identified a relationship among the plasminogen activator system, endometrial cells, and the ECM. 24 However, no study has reported a porcine embryo in vitro culture system for endometrial tissue and endometrial cells based on collagen gel.
Growth factors bind with the ECM, 25 and vascular endothelial growth factor (VEGF) adheres strongly to collagen type I.26,27 VEGF levels in the endometrial tissue increase during the preluteal phase compared with during the late luteal phase,28,29 and this effects the growth of endometrial, endothelial, and epithelial cells, as well as embryo development. 4
We previously used a 3D endometrial, epithelial, and stromal cell coculture system based on collagen gel to investigate embryo development; however, the rates of embryo development and blastocyst formation were low due to differences in the culture conditions required for the epithelial cells and the embryos (e.g., media components). 30 Previously, coincubating collagen gel with endometrial epithelial cells improved porcine embryo development. 31 Therefore, in this study, we investigated the effects of coincubating collagen gel with IL-1β-treated endometrial tissue on embryo development in pigs.
Materials and Methods
Isolation and culture of endometrium
All procedures involving the use of animals were approved by the Kangwon National University Institutional Animal Care and Use Committee (KIACUC-09-0139). The estrous cycle was induced and synchronized in 12 crossbred gilts (Yorkshire × Landrace; weight, 140–160 kg) using a hormone treatment. 32 The uteri were obtained from follicular-phase gilts (n = 6; age, 7.2 ± 0.6 months) at a local slaughterhouse (Pocheon Farm, Pocheon, Korea) and transported to the laboratory within 2 h at 4°C. The uteri were in the follicular phase according to ovarian morphology. The uteri were washed in Hank's Balanced Salt Solution, and the endometrial integument was removed from the uterine horn. Each endometrial tissue (0.30 ± 0.01 g) was preincubated in Dulbecco's modified Eagle's medium-Ham's F12 (D/F12; Sigma, St. Louis, MO) medium without fetal bovine serum (FBS) in 5% CO2 at 38.5°C for 24 h (Fig. 1A). The endometria were transferred to a culture medium (D/F12) without FBS containing 10 ng/mL IL-1β 33 in 5% CO2 at 38.5°C for 24 h (Fig. 1B). IL-1β-treated endometrial tissues were stored at −80°C until experiment.
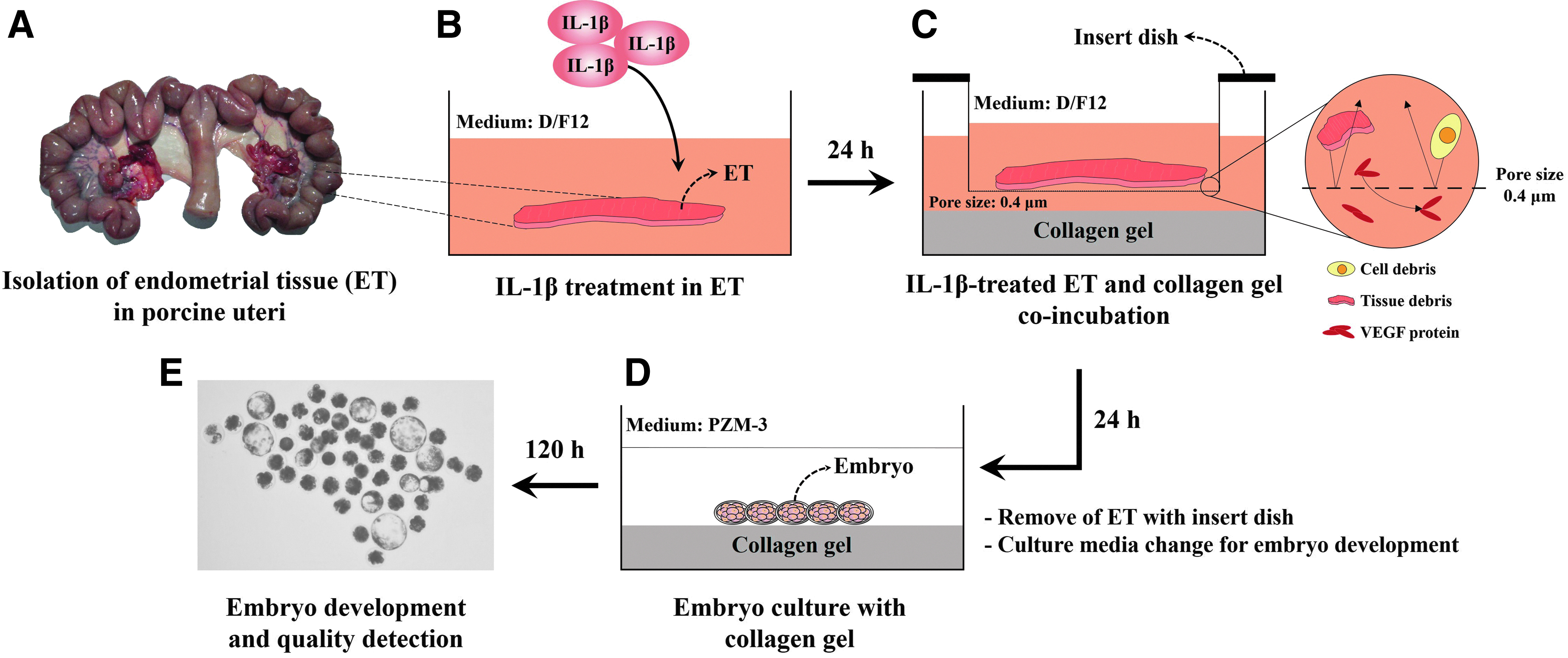
Experimental designs, follicular phase uterine used to isolate endometrial tissues
Preparation of collagen gels and coincubation with endometrial tissue
The collagen gel was made from a type-1 collagen matrix (Cellmatrix Type I-A; Nitta Gelatin, Inc., Osaka, Japan). A 160 μL aliquot of the Cellmatrix Type-I-A liquid was mixed with 40 μL buffer (0.05 N NaOH, 2.2% NaHCO3, and 200 mM HEPES) in a 24-well culture dish (SPL, Seoul, Korea) and was incubated for gelation in 5% CO2 at 38°C for 20 min. Then, the collagen gels were coincubated with IL-1β-treated endometrial tissue in an insert dish (pore size 0.4 μm; SPL) to avoid contamination from tissue and cell debris in D/F12 without FBS in 5% CO2 at 38°C for 24 h (Fig. 1C). The endometrial tissue, insert dishes, and D/F12 medium were removed from the 24-well culture dishes, and only collagen gel (C), C with IL-1β-untreated endometrial tissue (CE), and C with IL-1β-treated endometrial tissue (CEI) were washed three times in an embryo culture medium (porcine zygote medium-3; PZM-3). A 650 μL aliquot of PZM-3 was preincubated in 5% CO2 for 3 h at 38°C before embryo culture (Fig. 1D).
Quantitative reverse transcription polymerase chain reaction
The total mRNA of IL-1β-treated endometrial tissues was extracted using TRIzol (TaKaRa Bio, Shiga, Japan). mRNA concentration was measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher, Wilmington, DE). Total mRNA was mixed with the Reverse Transcription Premix Kit (Intron Biotechnology, Gyeonggi-do, Korea) for cDNA synthesis. Reverse transcription was performed at 45°C for 60 min and at 95°C for 5 min. The PTGS-2 primer was used to conduct polymerase chain reaction (PCR), and β-actin was used as an internal standard. Subsequent PCR analysis was performed according to the primer conditions (Table 1). Then, the products were analyzed with 2% agarose gel electrophoresis stained with 0.5 μg/mL ethidium bromide, visualized with UV light, and photographed. mRNA expression was analyzed with ImageJ software (NCBI, NIH), and the relative amount of mRNA was calculated by normalization with a housekeeping gene. Luteal phase porcine endometrial tissue was used as the positive control.
PTGS, prostaglandin synthase.
Detection of VEGF protein in collagen gels
The collagen gel coincubated with endometrial tissue samples were lyophilized and resuspended in mammalian protein extraction buffer (M-PER®; Thermo Scientific, Waltham, MA). Then, the samples were rotated for less than 1 h and centrifuged at 12,000 g for 10 min at 4°C. The supernatant was transferred to a new microcentrifuge tube, and the protein concentration was measured by the Bradford method. The proteins (70 μg/20 μL) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis at 130 V for 70 min, transferred to a polyvinylidene difluoride membrane for 60 min at 90 V, and incubated in blocking solution (5% skim milk in Tris-buffered saline/0.5% Tween-20; TBS-T) for 1 h at room temperature. The membranes were incubated with rabbit anti-VEGF (1:200, sc-25784; Santa Cruz Biotechnology, Santa Cruz, CA) and rabbit anti-β-actin IgG (1:1000, NB600-503; Novus Biological, Littleton, CO) overnight at 4°C. The membranes were then washed three times with TBS-T and incubated for 1 h at room temperature. The membranes were incubated with goat anti-rabbit IgG horseradish peroxidase (1:1000, sc-2030; Santa Cruz Biotechnology) at room temperature for 2 h, washed three times with TBS-T, and visualized using the West Save Enhanced Chemiluminescence Kit (AbFrontier, Austin, TX). Protein expression was measured using the EZ-Capture II system (ATTO, Tokyo, Japan), and protein band intensity was calculated using ImageJ software (NCBI).
In vitro fertilization
Ovaries were collected from gilts at a local slaughterhouse and transported to the laboratory within 2 h. Cumulus–oocyte complexes (COCs) were aspirated from antral follicles (3–6 mm) with an 18-gauge needle, and intact COCs were selected using a mouth pipette under a microscope. The COCs were transferred to an in vitro maturation medium consisting of modified TCM-199 with 10% (v/v) porcine follicular fluid (pFF), 5 μg/mL luteinizing hormone, 0.5 μg/mL follicle-stimulating hormone, 0.01 μg/mL epidermal growth factor (EGF), and 10 IU human chorionic gonadotropin and cultured for 22 h. Subsequently, the COCs were cultured in a medium with 10% pFF and 0.01 μg/mL EGF for 22 h. All cultures were performed at 38.5°C in 5% CO2.
The COCs were transferred to an oocyte maturation medium containing 0.1% hyaluronidase to remove the cumulus cells. The fertilization medium was based on modified Tris-buffered medium (mTBM) with caffeine. The mature oocytes were washed three times in mTBM and transferred to a 25 μL droplet of mTBM covered with paraffin oil (15 oocytes/droplet). Sperm were collected from five terminal Duroc cross (Gumbo, Wonju, Korea) boars (28.6 ± 5.3 months), washed in modified Modena B semen extender,
34
and resuspended to 6.0 × 105 sperm/mL in mTBM; 50 μL aliquots were added to a fertilization drop. The sperm–oocyte drops were incubated for 6 h at 38.5°C under 5% CO2. After fertilization, the extra sperm and cumulus cells were removed from the eggs by repetitive pipetting. The eggs were washed three times in PZM-3 and incubated at 38.5°C and 5% CO2. After 48 h, the embryos were transferred to the collagen gels (C, CE, and CEI) in 24-well culture dishes and cultured for 120 h in PZM-3. Then, embryo cleavage and blastocyst formation were detected under a microscope (BX50; Olympus, Tokyo, Japan) at 38.5°C (Fig. 1E) and early, expanded, hatching, and hatched blastocysts were classified according to morphology (Supplementary Fig. S1; Supplementary Data are available online at
Nucleus staining of blastocysts
Blastocysts after experiment at 120 h were fixed for 15 min in 4% paraformaldehyde in PBS-polyvinyl alcohol (PVA) at room temperature. The blastocysts were transferred to 5 μg/mL of Hoechst 33342 in PBS-PVA and incubated in a dark room for 30 min. Then, the embryos were placed on slide glasses mounted with slightly compressed coverslips in 247.64 μM 1,4-diazabicyclo[2.2.2]octane (Sigma) in glycerol (Sigma). The blastocysts were observed using a fluorescence microscope (Olympus). The total number of nuclei was recorded for each blastocyst.
Statistical analyses
Data were analyzed using SAS ver. 9.4 software (SAS Institute, Cary, NC). Data are presented as mean ± standard error. Treatment groups were compared for differences using the least significant difference test. All data were analyzed using one-way analysis of variance. A p < 0.05 was considered significant.
Results
Expression of PTGS-2 mRNA in IL-1β-treated endometrial tissue
Changes in PTGS-2 mRNA in the IL-1β-treated follicular-phase porcine endometrial tissue are shown in Figure 2. PTGS-2 mRNA expression increased significantly during the luteal phase (positive control) compared with during the follicular phase (p < 0.05). PTGS-2 mRNA expression also increased in the 10 ng/mL IL-1β-treated endometrial tissue compared with the untreated endometrial tissue (UT). In particular, PTGS-2 mRNA expression was significantly (p < 0.05) higher in the 10 ng/mL IL-1β-treated endometrial tissue than in the UT group.
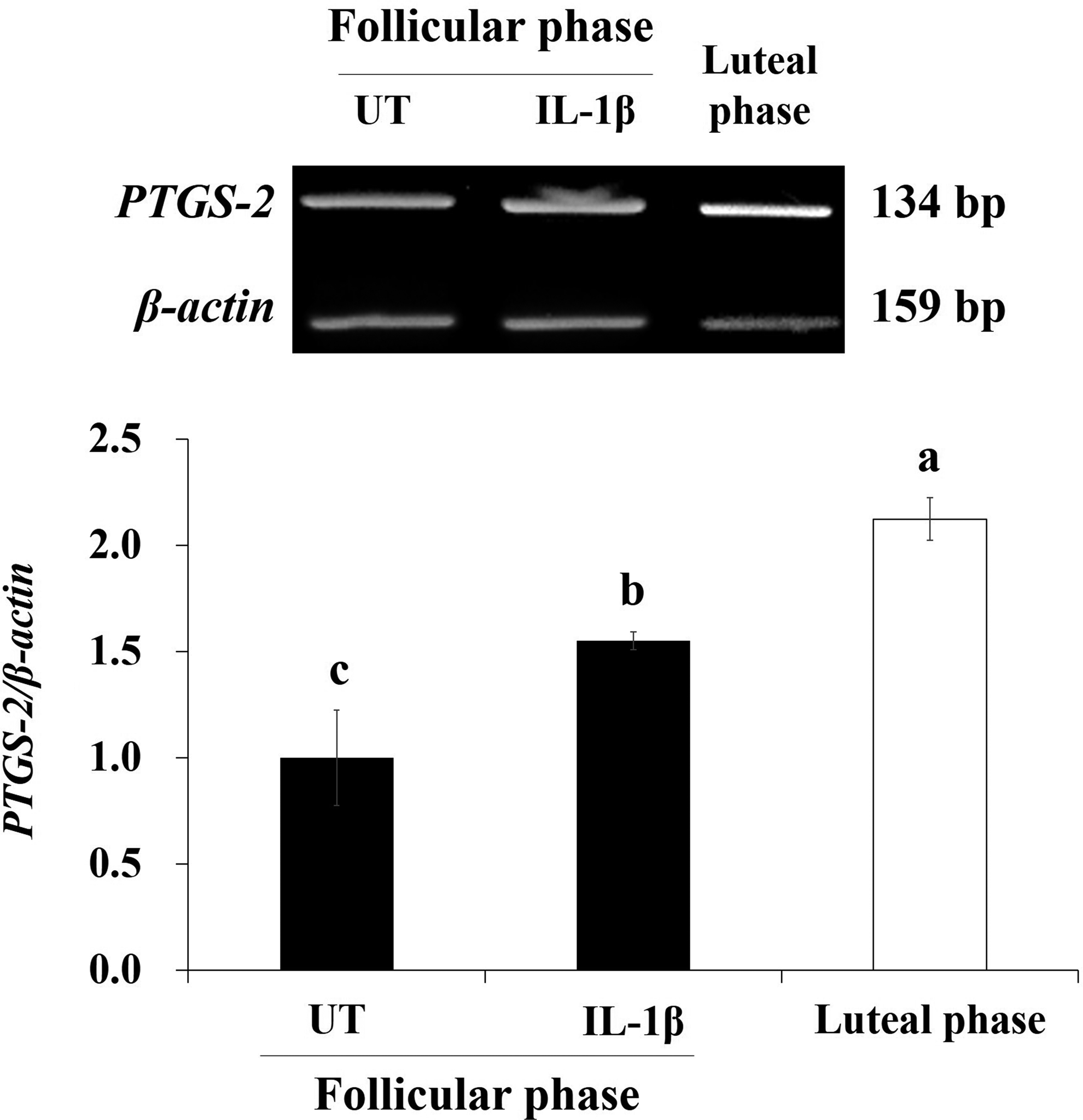
Effects of 10 ng/mL IL-1β on prostaglandin synthase-2 (PTGS-2) mRNA in follicular-phase porcine endometrial tissue. The untreated (UT) group involved untreated follicular-phase endometrial tissue. Porcine endometrial tissue in the luteal phase was used as a positive control. Bars represent mean ± standard error. a–cWithin the histogram, similarly shaded bars with different letters are significantly different (p < 0.05).
Detection of endogenous VEGF protein in coincubated collagen gels with endometrial tissue
VEGF protein expression levels in IL-1β-treated porcine endometrial tissue coincubated with collagen gel are shown in Figure 3. VEGF was not detected in the C treatment; however, VEGF protein intensity significantly increased in the CEI treatment groups compared with the CE group (Fig. 3A, p < 0.05). β-actin was not detected in all the treatment groups (Fig. 3B).

Vascular endothelial growth factor
Effect of collagen gels on embryo development and blastocyst formation rates in pigs
The effects of collagen gels with coincubating IL-1β-treated porcine endometrial tissue on the rates of embryo development and blastocyst formation are summarized in Table 2. Egg cleavage rates at 48 h after IVF did not differ among the treatment groups (data not shown). Day-2 embryos were cultured with collagen gel (C, CE, and CEI), but cleavage rates and blastocyst formation rate did not significantly differ among the groups (Table 2).
C, collagen gel; CE, coincubated C with endometrial tissue; CEI, coincubated C with 10 ng/mL interleukin-1 beta-treated endometrial tissue, n = 5; IVF, in vitro fertilization.
Effect of collagen gels on proportion and cell number of porcine blastocysts
The influence of collagen gels with coincubating IL-1β-treated porcine endometrial tissue on proportion and cell number in porcine blastocysts are shown in Figure 4. More blastocysts hatched in the CEI treatment groups than in the other treatment groups; however, the rates of early, expanded, hatching, and hatched blastocysts did not significantly differ among the groups (Fig. 4A). Blastocyst cell number was significantly (p < 0.05) higher in the CEI group than in the other groups (Fig. 4B).
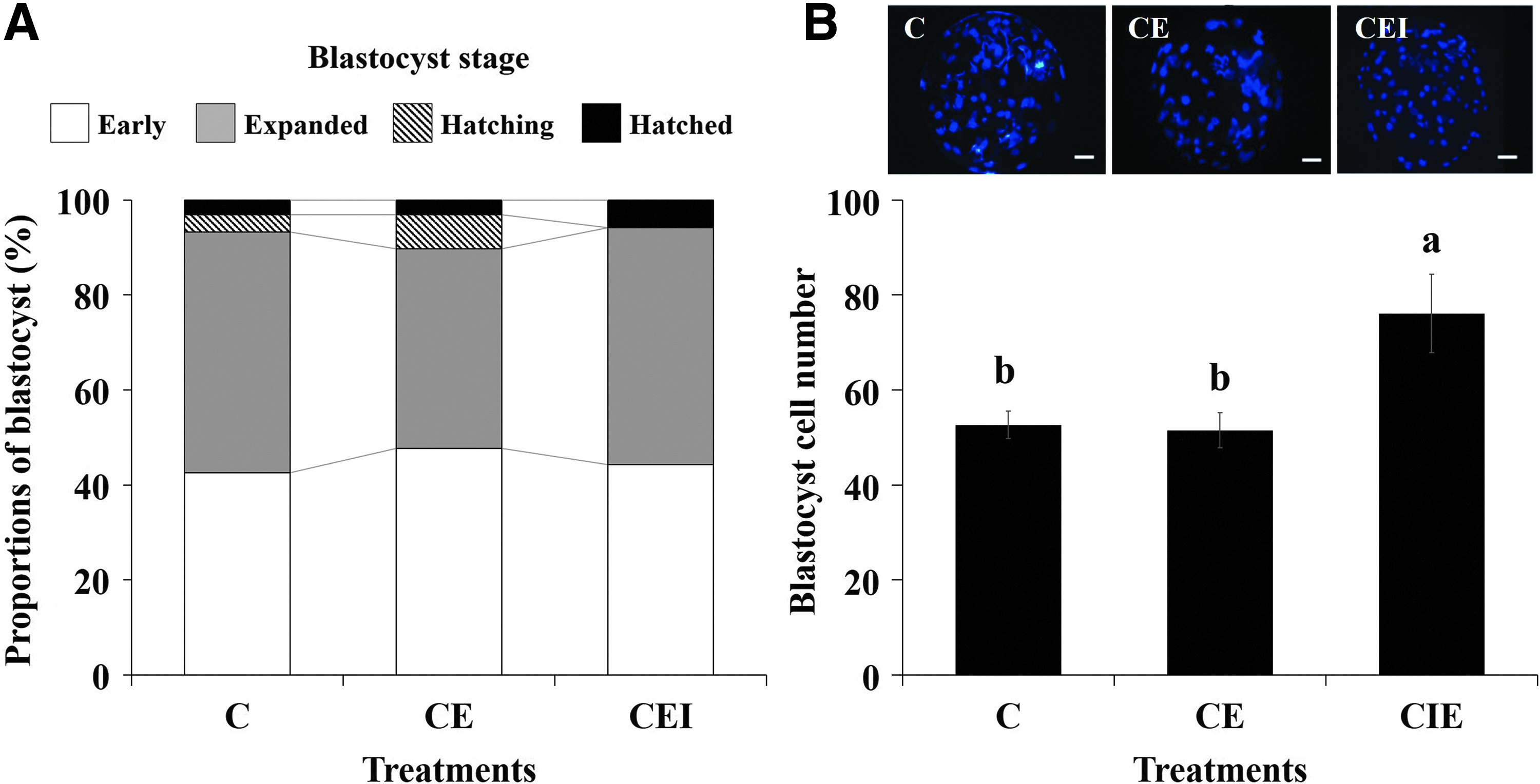
Effect of coincubating collagen gel with endometrial tissue on proportion of blastocysts
Discussion
Coculture systems have been used to study interactions among different types of cell.35,36 Some studies have investigated the culture conditions for various types of cell, such as embryo–endometrial cells, 31 and embryo–luteal cells. 37 Some studies concluded that the use of a common medium for culturing embryo, endometrial, and luteal cells is limiting.31,37 To solve this problem, endometrial tissues were coincubated with collagen gel because collagen gel (type I) provides endometrial structure, binds VEGF from the endometrial tissue, 26 and does not have a limiting effect in the embryo culture.
PGs are produced by the endometrial tissue during luteal phase of estrous cycle and early pregnancy in pigs. 38 PTGS is synthesized from PGs, which play roles in luteolysis, implantation, and recognition in the female reproductive tract. 39 PTGS-2 increases in the endometrial tissue from the follicular to the luteal phases of the estrous cycle,12,13 and IL-1β is also increased in the endometrial tissue during the days 10–14 of the estrous cycle (luteal phase). 40 In addition, IL-1β increased PTGS-2 mRNA of porcine endometrium in a dose-dependent manner. 33 Eventually, embryo locates on endometrial tissue of the uterus when PTGS-2 level is increased,7,18 and at this time, the endometrial tissue secretes various angiogenic factors for successful embryonic development.41,42 In this study, PTGS-2 expression was higher during the luteal phase compared with during the follicular phase, and was increased by IL-1β in the follicular-phase endometrial tissue. Thus, follicular-phase endometrial tissue developed a physiological state similar to that of luteal-phase endometrial tissue in response to IL-1β.
The VEGF family of proteins is involved in angiogenesis and cell proliferation and can bind to ECM components, such as collagen. 43 The maturation ability and in vitro fertility of COCs are promoted during in vitro maturation, 44 and the development of embryos created by IVF and somatic cell nuclear transfer improve by adding 5 ng/μL human recombinant VEGF to the in vitro culture. 4 However, a study reporting the effects of VEGF from porcine endometrial tissue on porcine embryo development has not been reported. In our previous study, we detected a positive effect of coincubated collagen gel with IL-1β-treated endometrial epithelial cells on porcine embryo development and VEGF.
Collagen type I is used to treat wounds, in dentistry, as an experimental material to investigate interactions with the ECM, and to establish 3D culture systems. 23 In addition, collagen type I is a major ECM component in the endometrial tissue that provides support to surrounding cells. 19 In this study, we found that VEGF from porcine endometrial tissue was successfully attached with type I collagen gel; however, no β-actin protein was observed in endometrial tissue coincubated with type I collagen gel. Because VEGF is secretion proteins, 43 and β-actin is cytoskeletal protein of cells, 45 we suggested that VEGF, excluding tissue and cell debris from endometrium, be passed through membrane of insert dish, and only then VEGF protein was attached in collagen gels. In addition, VEGF in collagen gels with coincubating porcine endometrial tissue was detected using western blot, but VEGF protein concentration of collagen gels could not be observed. The VEGF concentration of in vivo endometrial tissue changes during estrous cycle and 5 ng/μL human recombinant VEGF of culture media improve embryo development in pigs. 4 Thus, accurate concentration of VEGF from endometrial tissue in collagen gels may help in vitro embryo culture. Based on these results, we are undertaking additional studies to analyze the VEGF protein concentration of collagen gels with coincubating porcine endometrial tissue.
Preimplantation porcine embryos almost hatch as blastocysts to implantation after fertilization. 46 At this time, IL-1β, PTGS-2, and VEGF are regulated in the porcine intrauterine environment.12,13 High cell number of blastocyst increases hatching efficiency. 47 Generally, the number of cells per in vivo- and in vitro-derived blastocysts on day 7 after fertilization has been reported as average 80 to 140 and 40 to 60 cells in pigs. 46 Additionally, pregnancy and birth rate were increased when in vivo-derived blastocysts are transferred in pigs. 48 Therefore, blastocyst cell number influences pregnancy and birth rate in pigs. Practically, the total cell number of in vitro-derived blastocysts influence reproductive performance in pigs, pregnancy and efficiency of piglet production were improved in higher than lower cell number in in vitro-derived porcine blastocysts.49,50 Thus, the total cell number of blastocyst is very important in embryo development, blastocyst cell number is directly involved in fetus elongation and implantation in pigs, 46 and this being so, blastocyst cell number is used to decide embryo quality. In this study, the VEGF protein from IL-1β-treated endometrial tissue attached with collagen gel increased the number of blastocyst cells during in vitro culture. Especially, collagen gels coincubated with IL-1β-treated endometrial tissue improved the in-vitro-derived blastocyst cell number (76.1 ± 8.3), in which results were higher than normal condition in-vitro-derived blastocyst cell number in other studies.51,52 Thus, our results suggest that VEGF from IL-1β-treated follicular-phase endometrial tissue in collagen gel may have a positive effect on blastocyst quality
In this study, we developed a porcine IVF system using a combination of angiogenic factors from endometrial tissue and the ECM, which were applied to study the influence of the endometrial ECM and growth factors on embryo development. In addition, biomaterials of ECM family could be used to apply in vitro culture system and improve production efficiency in animal embryos. These results may help improve our understanding of the relationship between the intrauterine environment and embryos during the preimplantation phase.
Footnotes
Acknowledgment
This research was supported by the Agricultural Biotechnology Development Program (IPET31060-05), Ministry of Agriculture, Food, and Rural Affairs, of the Republic of Korea.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.