
Abstract
Hydrogels can facilitate nucleus pulposus (NP) regeneration, either for clinical application or research into mechanisms of regeneration. However, many different hydrogels and culture conditions for human degenerated NP have been employed, making literature data difficult to compare. Therefore, we compared six different hydrogels of natural polymers and investigated the role of serum in the medium and of osmolarity during expansion or redifferentiation in an attempt to provide comparators for future studies. Human NP cells of Thompson grade III discs were cultured in alginate, agarose, fibrin, type II collagen, gelatin methacryloyl (gelMA), and hyaluronic acid–poly(ethylene glycol) hydrogels. Medium containing fetal bovine serum and a serum-free (SF) medium were compared in agarose, gelMA, and type II collagen hydrogels. Isolation and expansion of NP cells in low compared to high osmolarity medium were performed before culture in agarose and type II collagen hydrogels in media of varying osmolarity. NP cells in agarose produced the highest amounts of proteoglycans, followed by cells in type II collagen hydrogels. The absence of serum reduced the total amount of proteoglycans produced by the cells, although incorporation efficiency was higher in type II collagen hydrogels in the absence than in the presence of serum. Isolation and expansion of NP cells in high osmolarity medium improved proteoglycan production during culture in hydrogels, but variation in osmolarity during redifferentiation did not have any effect. Agarose hydrogels seem to be the best option for in vitro culture of human NP cells, but for clinical application, type II collagen hydrogels may be better because, as opposed to agarose, it degrades in time. Although culture in SF medium reduces the amount of proteoglycans produced during redifferentiation culture, isolating and expanding the cells in high osmolarity medium can largely compensate for this loss.
Introduction
C
As yet, there is no consensus on the most suitable hydrogel for in vitro mechanistic studies on NP regeneration, or for regeneration in vivo. 6 Commonly, studies on hydrogel-based regeneration are performed with one particular hydrogel. However, this study setup does not enable assessment to what extent such a novel hydrogel suitable for further use, that is, represents an improvement over other previously studied gels. Various hydrogels are currently being used for NP cell culture and ECM production. Natural polysaccharides, mimicking the glucosamine-rich part of the NP matrix, and collagen-derived matrices, have been obvious choices. Alginate is a polysaccharide derived from brown algae and can be ionically cross-linked by the addition of divalent ions such as Ca2+, Ba2+, or Mg2+. This gentle cross-linking method makes it a very suitable material for cell culture. It has been shown to support matrix formation by NP cells in vitro.7–9 The seaweed-based polysaccharide agarose is receiving increasing attention in the field of IVD regeneration.10–13 Hyaluronic acid (HA) is a glycosaminoglycan (GAG) and component of the ECM of cartilaginous tissues such as the NP, and thus HA-based hydrogels provide a natural cell environment.14,15 Type II collagen hydrogels and type I collagen-derived scaffolds such as gelatin have also been shown to support the chondrogenic NP phenotype, that is, secretion of collagen type II and aggrecan.16,20 Gelatin can be functionalized with methacryloyl groups (gelatin methacryloyl [gelMA]) that allow subsequent cross-linking, resulting in a more stable and strong hydrogel, while still supporting matrix formation. 21 Although not a native part of the IVD, fibrin also has been used for chondrocyte, 22 annulus fibrosus (AF), 23 and NP cell encapsulation,24–27 due to its extremely good biocompatibility as evidenced by a lack of inflammation, foreign body response, or fibrosis, and thereby widespread clinical applicabilition.28–30
Although hyperosmolality is an important feature of the native NP tissue, most of the above-mentioned hydrogels have no intrinsic swelling pressure, and if they do (e.g., the swelling properties of alginate are known to depend on the type of alginate and cross-linking method,31,32 and those of agarose on concentration 33 ), they are usually used in nonconfined culture conditions. Controversy remains on the requirement of hyperosmolality for regeneration. Although at the mRNA level, hyperosmolality increased aggrecan and type II collagen expression in human NP cells, no effects were shown at the protein level. 19 However, as these data were based on NP cells expanded in monolayer in normosmotic conditions and redifferentiated in high-density culture without hydrogels, possibly the NP cells lost their responsiveness to hyperosmolality or required a natural local milieu. Articular chondrocytes expanded in high osmolality, showed a higher ECM producing capacity, 34 and produced stronger constructs, although in this study, hyperosmolality during redifferentiation even had negative effects. 35 To our knowledge, the role of hyperosmolarity during expansion has not yet been studied for NP cell-mediated regeneration.
To provide the NP research field with a possible comparator for future research on NP regeneration in hydrogels, this study compares the above-mentioned six commonly used natural hydrogels for ECM production by NP cells. These gels were used in the presence of serum or in a more defined, serum-free (SF) differentiation medium. Furthermore, the role of osmolality during expansion and redifferentiation culture in ECM production was investigated in the two best performing hydrogels.
Materials and Methods
Culture
Cell isolation and expansion
Human IVDs were collected during autopsy (Local Medical Ethical Committee [METC] number 12–364). NP was conservatively separated from AF tissue by visual inspection. NP cells from 10 different donors (aged 47–69, mean age 63; Thompson grade III) were isolated in Dulbecco's Modified Eagle's Medium (DMEM) (31885; Gibco, Life Technologies, Carlsbad, CA) with 14 U/mL pronase (Roche, Basel, Switzerland) +110 U/mL DNAse (Sigma, St. Louis, MO) for 1 h followed by incubation with 13.7 U/mL collagenase (Roche) +110 U/mL DNAse overnight. Cells were passed through a 70 μm cell strainer, washed twice with medium, and collected by centrifugation. Cells were expanded in 2D culture using DMEM +10% heat-inactivated fetal bovine serum (FBS; HyClone, Life Technologies) +2% antibiotics mixture (final concentration 200 U/mL penicillin, 200 U/mL streptomycin, 50 mg/L amphotericin-B, and 50 mg/L gentamicin [Lonza, Basel, Switzerland]) and 10 ng/mL recombinant human basic fibroblast growth factor (R&D Systems, Minneapolis, MA). After the first week, antimicrobial agents were reduced to 100 U/mL penicillin +100 μg/mL streptomycin (pen/strep; Gibco). Expansion typically took 3–6 weeks and to minimize dedifferentiation, all cells were used at passage 2. Cell numbers and viability were determined manually with a Bürker-Türk hemocytometer and Trypan blue (Sigma). Cell viability was always at least 98%.
Hydrogel preparation
Hydrogels of six different polymers were used in NP cell culture in concentrations of polymer as suggested by the manufacturer or previously used in literature 6 (Table 1). Alginate (alginic acid sodium salt, medium viscosity; Sigma) 3% (w/v) solutions were mixed with the NP cell suspension (passage 2) to a final concentration of 2% (w/v) alginate and were added dropwise into a sterile 102 mM CaCl2 solution in which the beads were allowed to cross-link for 10 min. Alginate hydrogels required weekly recross-linking (i.e., incubation with CaCl2) for stability since otherwise, they would become softer due to ion leaching. Agarose (low electroendosmosis; Sigma) hydrogels were prepared by autoclaving a 3% (w/v) solution and maintaining it at 45°C until use. The temperature was lowered to 39°C before mixing in the NP cells (final concentration agarose 2% [w/v]), after which gelation was allowed to occur at room temperature (RT). GelMA was prepared as described before by reacting gelatin from porcine skin type A (Sigma) with methacrylic anhydride for 1 h at 60°C, followed by dialysis and freeze-drying. 36 Irgacure 2959 (0.05%; BASF, Ludwigshafen, Germany) was added to a 15% (w/v) gelMA solution and mixed 2:1 with the NP cell suspension to a final hydrogel concentration of 10%. GelMA hydrogels were prepared by injection into a Teflon mold 37 and UV cross-linking for 15 min at 365 nm at an intensity of 2.6 mW/cm2. Fibrin (commercially available as Tissucol; Baxter, Deerfield, IL) hydrogel was prepared according to the manufacturer's instructions. In short, fibrin was mixed with cells and an equal part of thrombin solution was added. Cross-linking occurred at 37°C for 15 min. HA/poly(ethylene glycol) (PEG) (HA/PEG commercially available as HyStem®; Esibio, Alameda, CA) solution was mixed with cross-link solution and left to cross-link at 37°C for 30 min. Type II collagen hydrogels (commercially available as the 3D collagen culture kit; EMD Millipore, Billerica, MA) were prepared by neutralizing the pH of the acidic 6 mM type II collagen solution in acetate buffer. Cells were mixed into this solution and left to cross-link at 37°C for 1 h. All hydrogels contained 2 × 106 cells/mL and were ∼50 μL each. To provide an indication of biomechanical properties of these gels, in an unrelated set of experiments with the same hydrogels, the elastic modulus was measured. For alginate, agarose, GelMA, fibrin, HA/PEG, and collagen II these were shown to be 4 ± 1, 29 ± 12, 11 ± 1.5, 4, 4 ± 2, 4 ± 1, and 2 ± 1 kPa, respectively (article submitted).
HA, hyaluronic acid; PEG, poly(ethylene glycol); gelMA, gelatin methacryloyl.
Hydrogel culture with NP cells
NP cells from seven donors were cultured in hydrogels in a medium with FBS (DMEM +10% FBS +0.2 nM ascorbic acid-2-phosphate [ASAP; Sigma] + 1% pen/strep) or in an SF differentiation medium (DMEM +2% insulin–transferrin–selenium-X [Gibco] + 0.2 nM ASAP +2% human serum albumin [Sanquin, Amsterdam, The Netherlands] + 1% pen/strep) for 28 days. To assess the effect of osmolality during expansion, NP cells of three donors were expanded in a medium of 340 mOsm/kg or in a medium with osmolality adjusted to 400 mOsm/kg by the addition of NaCl. After expansion for two passages, cells were incorporated in agarose, gelMA, or type II collagen hydrogels and cultured in the SF differentiation medium adjusted to 400, 450, or 500 mOsm/kg with NaCl (2, 3.7, or 5.5 mg/mL) and confirmed with a freezing point micro-osmometer (Advanced Instruments, Inc., Norwood, MA). In all experiments, empty hydrogels without cells were cultured and taken along as controls in all analyses.
Histology and immunohistochemistry
Live/dead staining (Life Technologies) of two hydrogels of each type was performed after 7 days of culture. Hydrogels were washed in phosphate-buffered saline (PBS) and incubated in PBS containing 2 μM calcein AM and 4 μM ethidium homodimer-1 for 20 min at 37°C. The staining solution was replaced by PBS and the hydrogels were examined with a fluorescent BX51 microscope, DP 70 camera, and cell^F software (Olympus, Tokyo, Japan). After 28 days of culture, hydrogels were fixed overnight in 4% buffered formalin, dehydrated, and embedded in paraffin. Five micrometer sections were mounted on Superfrost™ Ultra Plus microscope slides (Thermo Scientific). Before staining, sections were rehydrated through a graded alcohol series to demi water. For Safranin-O/Fast Green staining, sections were incubated with Mayer's hematoxylin (Merck, Kenilworth, NJ) for 5 min, washed in tap water for 10 min, and stained with 0.4% (w/v) Fast Green (Merck) for 4 min. After washing in 1% (v/v) acetic acid, 5 min staining by 0.125% (w/v) Safranin-O (Merck) was followed by dehydration and mounting. For aggrecan immunohistochemistry, antigen retrieval was performed by incubating in citrate buffer (10 mM sodium citrate, pH 6.0) at 96°C for 20 min and cooling to RT for an additional 20 min. For type II collagen, antigen retrieval consisted of incubation with pronase (1 mg/mL in PBS; Roche) followed by hyaluronidase (10 mg/mL in PBS; Sigma) (both at 37°C for 30 min). Immunostaining protocols were the same after the antigen retrieval. Sections were blocked with PBS +5% bovine serum albumin (PBS–BSA) for 30 min at RT and incubated overnight at 4°C with the primary antibody diluted in PBS–BSA (aggrecan [1:150; 6-B-4 [Novus Biologicals, Littleton, CO]) or type II collagen (0.4 μg/mL, DSHB II-II6B3 [DSHB, Iowa City, IA]). The EnVision kit (α-mouse; Dako, Glostrup, Denmark) was used for visualization according to the manufacturer's protocol. Microscopy pictures were taken with a BX43 upright microscope and CellSens software (Olympus). Exposure was adjusted in Photoshop (Adobe, San Jose, CA) for descriptive analysis of the images.
Biochemistry
After 28 days of culture, 3–5 hydrogels per donor and hydrogel type were incubated overnight at 60°C in a papain digestion buffer with 250 μg/mL papain (Sigma) and 1.57 mg/mL
Statistics
All statistical analyses were carried out with SPSS 21 (IBM, North Castle, NY). Outliers determined with the 5*σ rule were removed (a value is considered an outlier when it is outside the mean plus or minus five times the standard deviation range). For all data, Kruskal–Wallis tests were performed, with post hoc multiple Mann-Whitney tests with a Benjamini-Hochberg correction. All graphs display data as mean ± standard deviation.
Results
Comparison of different hydrogels for NP cell culture
Hydrogel handling
Cross-linking of type II collagen hydrogels resulted in shrinking of the hydrogels with fluid expelled on top of the hydrogels. The type II collagen hydrogels were also very soft and difficult to handle, as were the HA/PEG hydrogels. During culture, degradation was seen of type II collagen and HA/PEG hydrogels, and most notably for fibrin. The fibrin hydrogels had completely disappeared after 2–3 weeks of culture, ending up with a cell pellet in the presence of FBS. Alginate, agarose, and gelMA hydrogels were very stable. Collagen gels already showed degradation within the first week of culture. After 28 days, this resulted in clearly smaller constructs in the differentiation medium and even in hydrogel-free pellets upon culture in the presence of FBS (Supplementary Fig. S1; Supplementary Data are available online at
Histology and immunohistochemistry
All cell-free hydrogels were negative for background immunohistochemical staining (data not shown). Cell-free alginate and HA/PEG hydrogels stained positive for Safranin-O, as both have a net negative charge. Fast Green stains collagenous fibers and thus also stained cell-free gelMA and type II collagen hydrogels green. Figure 1 displays live/dead staining performed after 7 days of culture, and histology and immunohistochemistry after 28 days. In all hydrogels, primarily live cells were present. Microscopic tissue-like clusters were formed by several cells in lacuna-like cavities inside alginate and HA/PEG hydrogels. These clusters were positive for Safranin-O, suggesting the presence of proteoglycans, although the background staining found cannot fully confirm this. However, immunohistochemistry indicated the presence of aggrecan and type II collagen. Also, around individual cells in alginate, positive aggrecan immunostaining was visible. Diffuse pericellular Safranin-O staining was present in the agarose hydrogels, indicating GAG and hence proteoglycan production, and this matrix was also immunopositive for aggrecan and type II collagen. Very little matrix seemed to be deposited in gelMA hydrogels, as no staining was seen for any type of (immuno)staining, except for Fast Green. Since the fibrin had completely disappeared, histology resembled cells grown in pellets. Safranin-O staining indicated that proteoglycans were present, but very limited aggrecan staining was visible. Type II collagen was present throughout the matrix. In type II collagen hydrogels, Safranin-O staining was absent and aggrecan and type II collagen staining was sparse.
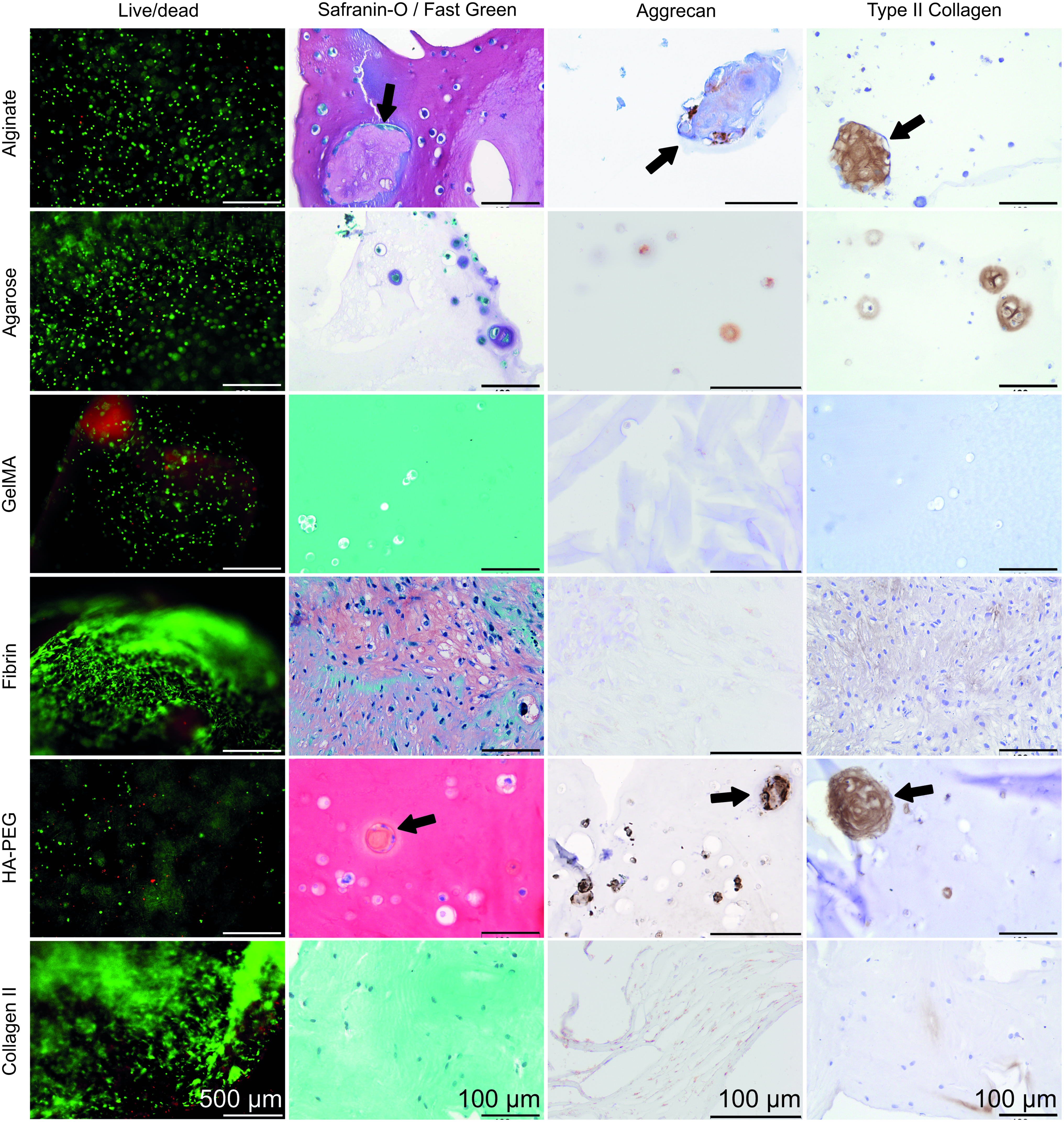
Live/dead staining after 7 days of culture shows that viable cells outnumber dead cells in all hydrogel types. Histology for Safranin-O (red staining) and Fast Green (green staining), and immunohistochemistry for aggrecan and type II collagen (brown staining) after 28 days show that matrix was deposited in majority of the hydrogels. Matrix accumulated directly around the cells in most hydrogels, but in fibrin hydrogels, matrix was formed more dispersedly since hardly any hydrogel was left at the end of culture. In alginate and HA/PEG hydrogels, microscopic tissue-like structures were formed (see arrows), positive for Safranin-O, aggrecan, and type II collagen. Scale bar represents 500 μm in live/dead images and 100 μm in histology images. HA, hyaluronic acid; PEG, poly(ethylene glycol). Color images available online at
GAG and DNA content
After correction for the signal of cell-free hydrogels, cells in agarose and type II collagen hydrogels were shown to produce the highest amount of GAGs (p ≤ 0.014, Fig. 2), whereas fibrin and HA/PEG hydrogels contained more GAGs than alginate hydrogels (p ≤ 0.016). In fibrin and type II collagen hydrogels, DNA content was higher than in alginate, agarose, and gelMA hydrogels (p < 0.007), and DNA content in HA/PEG was higher than in gelMA (p = 0.01). When GAG content was normalized to DNA, this was found to be highest for NP cells cultured in agarose hydrogels (p < 0.001 compared to alginate, fibrin, and HA/PEG, p = 0.01), although there was no significant difference with gelMA. More GAG/DNA was produced in type II collagen than in alginate or fibrin hydrogels (p > 0.001 and p = 0.008, respectively), and GAG/DNA in HA/PEG was higher than in alginate (p = 0.02).
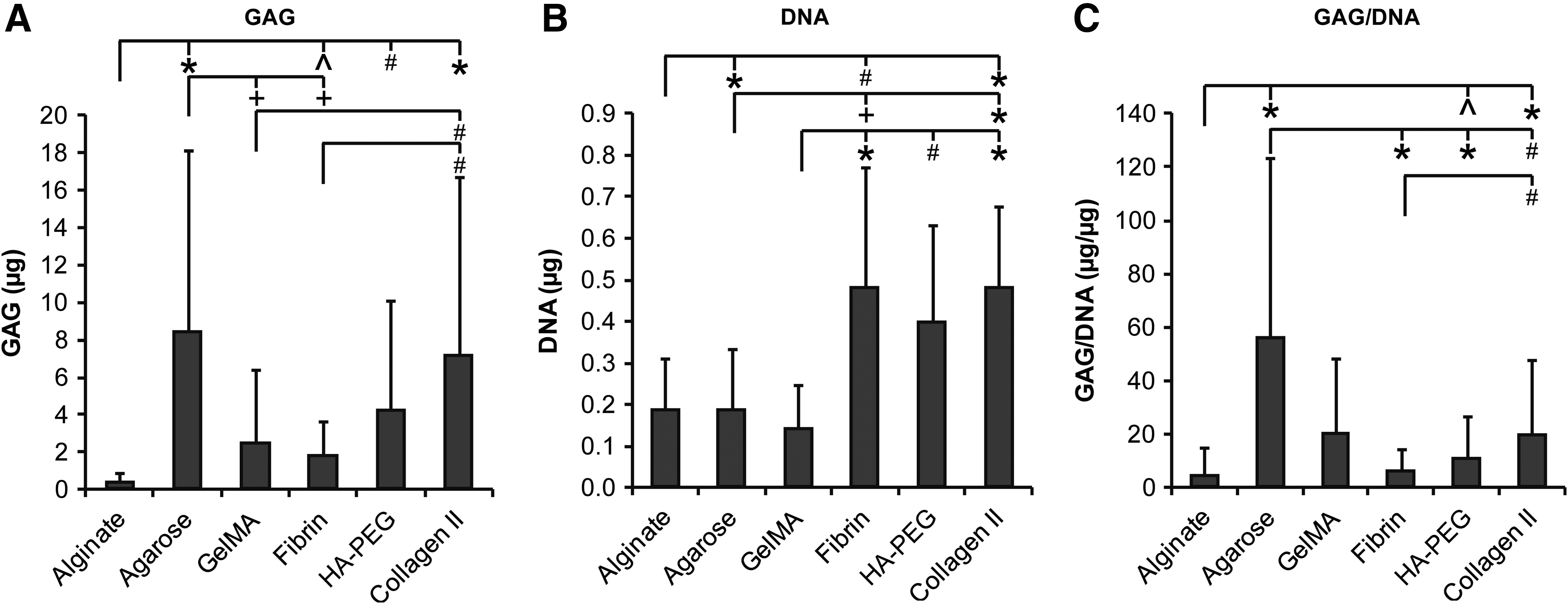
NP cells of four different donors were cultured in six different types of hydrogels for 28 days (n = 3 per condition). All measurements were corrected with the background values of empty hydrogels.
Effect of serum in culture
Although serum is a common addition to cell culture, clinical application precludes the use of animal-derived components such as serum. Therefore, the response of cells to the best-performing polysaccharide hydrogel (agarose) and the collagen-based hydrogels (gelMA and type II collagen) was also tested in the absence of serum.
GAG and DNA content
NP cells deposited higher amounts of GAGs in the presence of FBS than cells in the SF differentiation medium in agarose and gelMA hydrogels (p ≤ 0.001, Fig. 3). DNA content was higher for cells cultured in agarose and type II collagen hydrogels with the SF differentiation medium. Higher GAG/DNA was measured with FBS in all three hydrogels (p ≤ 0.001). Total GAG production, that is, GAG content in the cultured hydrogels and GAGs released into the culture medium, was increased in the presence of FBS compared to the SF differentiation medium (p < 0.001 for all three hydrogels tested). Incorporation efficiency, determined as the percentage of total GAG deposited in the hydrogels, was higher in cells cultured in type II collagen hydrogels in the SF differentiation medium compared to the medium containing FBS (p < 0.001). This difference was not evident in agarose or gelMA hydrogels.
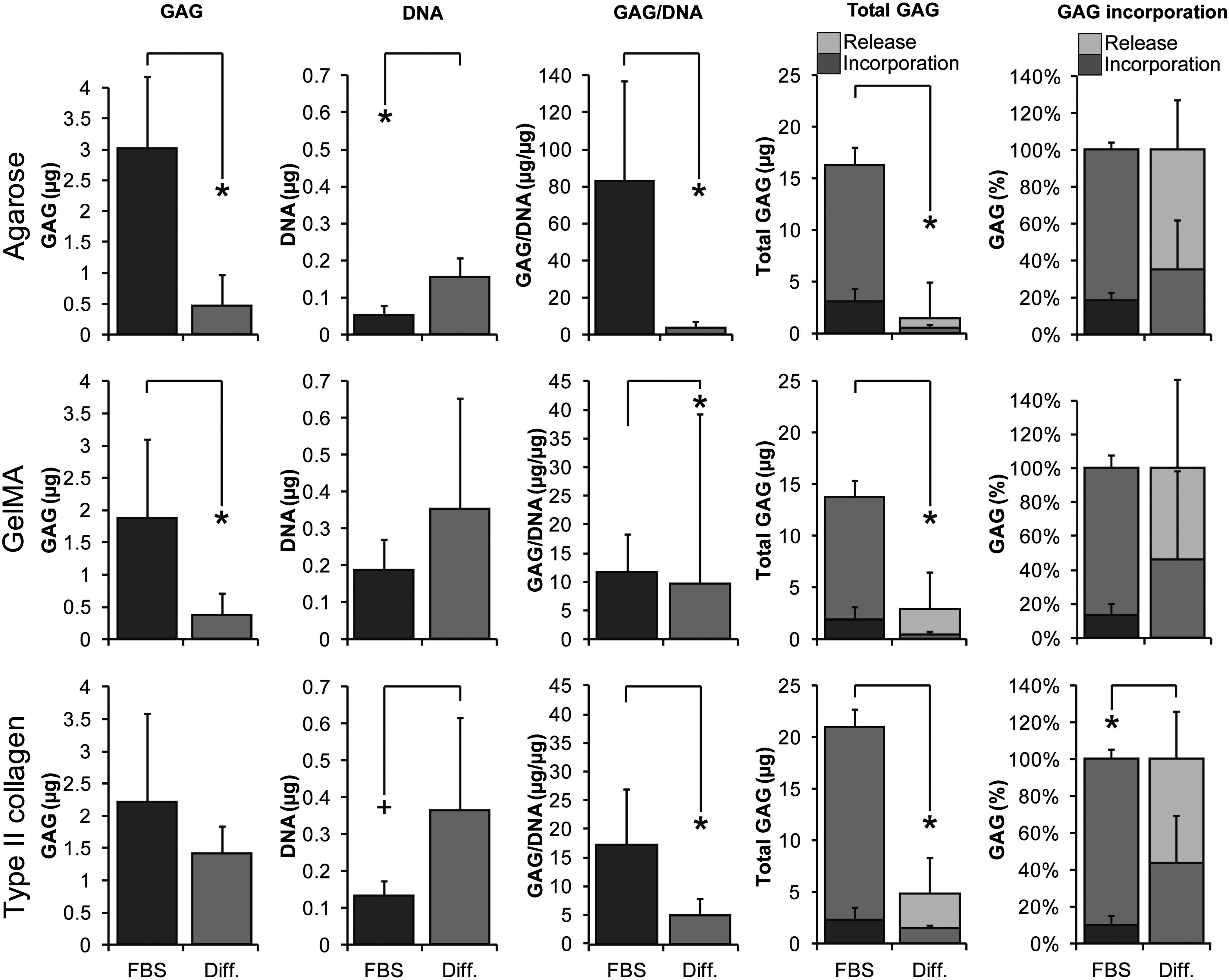
NP cells of three donors were cultured in hydrogels (n = 5 per condition per donor) in medium containing FBS or SF differentiation medium (Diff.). In agarose and gelMA hydrogels, significantly higher GAG, GAG/DNA, and total GAG production was measured compared to SF differentiation medium. DNA content was higher in the absence of FBS for agarose, but no differences in DNA content were detected in gelMA hydrogels. In type II collagen hydrogels, DNA and incorporation efficiency were higher in SF differentiation medium compared to medium with FBS, but GAG/DNA and total GAG production were higher in the presence of FBS. Significant differences are given as follows: *p ≤ 0.001 and +p ≤ 0.005. FBS, fetal bovine serum; SF, serum free.
The effect of medium osmolality during NP cell expansion and differentiation in hydrogels
As agarose and type II collagen stimulated GAG deposition by NP cells most, both in FBS and serum free medium, these hydrogels were further employed to determine the effect of osmolality during expansion and redifferentiation of NP cells.
Histology
NP cells isolated and expanded in 400 mOsm/kg showed more Safranin-O and type II collagen staining upon redifferentiation in both type II collagen and agarose hydrogels than cells isolated and expanded in 340 mOsm/kg (Fig. 4). This Safranin-O and type II collagen staining pattern remained the same even after tuning osmolality during redifferentiation up to 500 mOsm/kg.

NP cells isolated and expanded in either 340 or 400 mOsm/kg medium were cultured in 340–500 mOsm/kg SF differentiation medium.
GAG and DNA content
Redifferentiation of NP cells that were isolated and expanded in high osmolality medium in agarose hydrogels resulted in higher GAG, DNA, and GAG/DNA compared to hydrogels with cells expanded in normal osmolality (p < 0.001, Fig. 5A). Similar effects of osmolality in GAG and GAG/DNA production were found for cells cultured in type II collagen hydrogels (p < 0.001, Fig. 5B). In both hydrogels, GAG production appeared to be independent of osmolality during redifferentiation.

NP cells from three donors isolated and expanded in either 340–400 mOsm/kg medium were cultured in hydrogels (n = 5 per condition) in SF differentiation medium with an osmolality range of 340–500 mOsm/kg. GAG, DNA, and GAG/DNA content of the agarose hydrogels indicate that increased osmolality during isolation and expansion augmented the regenerative capacity of the NP cells. In type II collagen hydrogels, GAG and GAG/DNA were higher for cells isolated and expanded in 400 mOsm/kg medium compared to 340 mOsm/kg cells. Increasing osmolality during regeneration did not have any effect in either hydrogel. Significant differences are given as follows: *p ≤ 0.001.
Results are also summarized in the Supplementary Table S1.
Discussion
In this study, six commonly used hydrogels, the presence of serum in culture medium, and osmolality of isolation, expansion, and culture medium were compared for redifferentiation and ECM production by NP cells. There was a clear difference in ECM producing capacity of human degenerated NP cells in hydrogels of different natural polymers. More matrix was deposited in agarose and type II collagen hydrogels, and when normalized for DNA content, the capacity of the NP cells to produce ECM was highest in agarose. The presence of FBS clearly affected cell behavior, resulting in augmented deposition of proteoglycans in all three hydrogels. However, in type II collagen hydrogels, incorporation efficiency of GAG was decreased compared to the SF differentiation medium. Regardless of the hydrogel employed, isolation and expansion in high osmolality medium strengthened the differentiation of NP cells, as indicated by the higher GAG and GAG/DNA in SF culture conditions compared to normal osmolality. The anabolic effect of increasing osmolarity during redifferentiation, however, was inconsistent.
The comparison of six commonly used hydrogels in this study points toward two hydrogels that are relatively effective for human NP cell-based ECM deposition. These are agarose and type II collagen hydrogels, as indicated by the production and deposition of proteoglycans, as markers of late-stage NP cell differentiation together with collagen II, and matrix proteins crucial for tissue functionality. Although a large number of studies have been done with biomaterials for NP repair, to the best of our knowledge, only two directly compared the response of NP cells to different hydrogel types. In the first, an immortalized human NP cell line produced more proteoglycans, even when normalized to cell number, in alginate than in atelocollagen hydrogels in the presence of FBS. 38 Porcine NP cells and mesenchymal stem cells (MSCs) separately cultured in SF medium were also shown to deposit more ECM in 1.5% alginate than cells in chitosan hydrogels. 39 However, none of these studies referred to above were based on primary human NP cells isolated from degenerated IVDs, and/or the hydrogels used to compare against were not included in this study. Aside from the culture environment and the type of polymer used, the cross-link density of the hydrogels, the polymer concentrations, and the resulting mesh size and stiffness may affect cell behavior. 6 Research on the effect of these properties of NP cell behavior is limited. However, alginate concentrations varying between 1% and 6% were shown not to affect gene expression levels of caprine NP cells, 40 nor was matrix production by human NP cells affected in 1% versus 2% alginate or 1% versus 2% agarose. 41 Also, bovine NP cells produced similar amounts of GAGs in 3% versus 4% agarose. 42 Addition of HA to a type II collagen hydrogel decreased NP cell proliferation, but there were no differences between different HA concentrations, suggesting other causes than mere polymer concentration. 43 Cross-link density in a similar hydrogel influenced NP cell viability in the long term, but gene expression only in the short term. 44 Articular chondrocytes, however, did respond to increasing agarose concentration from 2% to 3% (w/v) with 30% increased GAG production, 45 and GAG production by equine chondrocytes in gelMA hydrogels decreased by around 40% with increasing polymer concentration from 10% to 20% (w/v), again stressing the difference between NP cells and articular chondrocytes. 47 It remains to be determined how the concentration or cross-link density of type II collagen may further affect human NP cell behavior. A disadvantage of the collagen II gel is its water expelling behavior during cross-linking. This makes it difficult to estimate final polymer density and may result in reduced porosity, altering cell behavior. We have also not further verified whether exposure to different osmolarity affected hydration level, nor assessed hydration after equilibration, after cross-linking of these gels. However, as a collagenous matrix does not have a very high fixed charge density, osmotic pressure of the gel most likely was not changed. Moreover, no effect on viability was noted, nor was regeneration in collagen II gel affected by media of varying osmolarity, suggesting that these differences in hydration, in any case, did not have profound biological effects. The lack of Saf O staining, known to be insensitive to low proteoglycan levels, 46 in contrast to the higher proteoglycan content as indicated by DMMB for sulfated GAGs, may suggest that in this gel, the deposited proteoglycans were distributed more evenly throughout the gel compared to the agarose gels.
Another major limitation in standardizing NP regeneration research is the variation in composition and biologic effects between batches of FBS, a well-known issue, for example, in chondrogenic differentiation of adipose-derived MSCs 48 that will likely affect NP cells as well. FBS contains many (un)defined growth factors and nutrients that influence matrix production and deposition. 49 In our study, NP cells in hydrogels produced more GAGs in the presence of FBS compared to a more defined and clinically relevant SF differentiation medium, which is in line with a previous report. 50 Moreover, cell-based clinical applications, where cells are being expanded before transplantation, would require the use of chemically defined media because of the risks associated with the use of xenogeneic serum and the possible contaminations it might contain. 48 Serum deprivation may lead to more autophagy and lower ATP levels in murine NP cells in normoxic conditions. 51 It should be noted, however, that the murine NP cells were cultured in SF medium containing only DMEM, while in this study, a medium containing pyruvate, albumin, insulin, transferrin, selenium, and ascorbic acid was used to support NP cell redifferentiation.
Osmolality during isolation and expansion of NP cells was clearly shown to define the ECM producing capacity of redifferentiating human NP cells, independent of the hydrogel employed and osmolality during redifferentiation. The observation that NP cells produce more matrix in differentiation culture after expansion in high osmolality, even when brought back to normal osmolality, suggests that the phenotype of cells is irreversibly altered during isolation and expansion at low osmolality. These findings are especially important when employing NP cells for cell-based therapies, for example, herniated NP cells for transplantation into a degenerated NP. 52 Although this is not a common strategy, we suggest that culture osmolality should be considered for any cell type used for NP regeneration. Interestingly, increasing osmolality during redifferentiation culture of NP cells isolated and expanded in standard osmolality medium, did not affect matrix production. This is in line with previous studies where increasing medium osmolality did not result in increased matrix production in high-density cultures of human NP cells 17 and bovine NP cells in alginate. 53 Similar effects of osmolality during different stages of culture have been observed for chondrocytes. For healthy and osteoarthritic human chondrocytes, isolation and expansion at an osmolality of 380 mOsm/kg improved the cell phenotype compared to using hypo-osmotic medium of 280 mOsm/kg. 34 Bovine chondrocytes expanded at 400 mOsm/kg, cultured in agarose hydrogels, produced constructs with increased mechanical strength compared to cells expanded at 300 mOsm/kg, independent of differentiation osmolality, 35 although GAG content was not different. In contrast, in articular chondrocyte 3D culture, increasing osmolarity during redifferentiation culture actually inhibited proteoglycan production.35,54 This again underlines the notion that, although NP cells are “chondrocyte like,” their distinct expression of different membrane markers, different composition of the native tissue, 55 and different biological responses mean that they should be studied as a unique cell type.
Conclusions
This study shows that the response of degenerated human NP cells to culture in a variety of natural hydrogels is different for each polymer. Overall, agarose and type II collagen hydrogels facilitate NP cells to produce high amounts of GAGs. Nondegrading agarose hydrogels may be suitable for studying NP redifferentiation, and degrading type II collagen hydrogels may be chosen for in vivo application, despite the partial dehydration found. Although redifferentiation of NP cells can be further improved, one of these hydrogels may be used in subsequent studies to compare against other candidate hydrogels, until a better reference is identified. Furthermore, while human NP cells do not thrive in the absence of FBS, their isolation and expansion in high osmotic medium can restore their level of matrix production. Moreover, the effect of expansion in high osmolarity merits further investigation. Possibly epigenetic changes, such as methylation and/or differential expression of non-coding RNA, such as miRNA, play a major role here. From a translational point of view, the responsiveness to growth factors after expansion in high osmolarity should be verified to see whether further enhancement of ECM production can be achieved.
Footnotes
Acknowledgments
The authors would like to thank Peter A. Levett for supplying the gelMA. This work was financially supported by the Dutch Institute for Regenerative Medicine and the Dutch Arthritis Foundation (LLP12 and LLP22). Federal and institutional funds were received in support of this work.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.