
Abstract
The human dental follicle (hDF) contains the developing tooth and is involved in regulating tooth maturation and eruption. To investigate the mesenchymal stromal cells of the dental follicle, 2 three-dimensional (3D) culture models were used, based on a dynamic bioreactor: the Rotary Cell Culture System (RCCS™) and the 3D culture of precursor cells isolated from follicular tissue (human dental follicle cells [hDFCs]). The hDFCs were obtained from impacted third molars of 20 patients. Two 3D culture models were tested. In the first model, intact hDF explants were cultured in 3D conditions, preserving the original tissue architecture; they were studied using histomorphological and molecular analyses. The second model involved the 3D culture of hDFCs, which were characterized to evaluate their multipotency in terms of differentiation capability. Of the biomarkers known to characterize hDFCs, hDF precursors were selected for our study. The immunophenotype and in situ immunocytochemistry were evaluated for markers CD44, CD90, CD146, CD105, CD31, CD34, and CD45 Ag. The results show that the conditions provided by the RCCS preserve the original organizational architecture of the cells. The 3D conditions of the model enhanced differentiation in response to adipogenic, osteogenic, and chondrogenic inductive growth media. The immunophenotype and the immunocytochemistry showed generally high expression of CD90, CD44, and CD105, while CD146 expression was more restricted to ∼30% of cells. No expression was observed for CD31, CD34, and CD45 Ags. Two 3D tissue- and cell-based ex vivo models of the hDF supported the long-term maintenance of hDF-specific cell phenotypes and their ability to recapitulate typical cellular differentiation states. As such, these ex vivo models could be used to study the physiopathology of human odontogenesis. In addition, in a therapeutic context, they could be used to examine the role of specific chemical signals (e.g., new therapeutic agents) in the processes of dental tissue repair and regeneration.
Introduction
T
Our study presents two 3D culture models using the fluid-dynamic Rotary Cell Culture System (RCCS™) bioreactor, which seems to be appropriate for the ex vivo/in vitro study of hDF features. The RCCS has been shown to generate optimal hydrodynamic and high mass transfer conditions, thus sustaining the long-term 3D culture of cells and tissue explants.11–17
In the first model, intact hDF explants were cultured in 3D conditions, thus preserving the original tissue organizational architecture. Alongside cell–cell and cell–extracellular matrix (ECM) interactions, this model enabled key components for tooth organogenesis and repair to be preserved.
18
The second model involved the 3D culture of precursor cells isolated from follicular tissue (human dental follicle cells [hDFCs]). While in the tissue culture-based model the native cell heterogeneity and tissue-specific microenvironment were maintained, in this second model hDFCs had to create their own, neo-formed microenvironment. Of the many markers known to characterize hDF stem cells (Supplementary Table S1; Supplementary Data are available online at
In this study we evaluated both 3D models. The first model (where intact hDF explants were cultured in 3D conditions) was characterized using histomorphological and molecular analyses. In the second model, precursor cells isolated from follicular tissue (hDFCs) were characterized in 3D culture to evaluate their multipotency. hDF tissue precursor cells were identified based upon immunophenotyping for known markers of hDF stem cells, including CD44, CD90, CD146, CD105, CD31, CD34, and CD45 Ag.
Materials and Methods
hDF-derived cells
hDFCs were obtained from impacted third molars from 20 patients of both sexes (10 females, 10 males) enrolled in the study (age range: 14–23 years). After signing a statement of informed consent (University of Milan, prot. 22897/2013LB), they underwent the extraction (for orthodontic reasons) of wisdom teeth, in the Department of Biomedical, Surgical and Dental Sciences, University of Milan, Milan, Italy, between September 2015 and September 2016.
Clinical and surgical procedures
All mandibular third molars were radiographically evaluated by orthopantomography. In those cases for which contact between the alveolar inferior canal and the third molar roots was suspected, indicated by the presence of Rood's signs, computed tomography or cone-beam computed tomography was performed. Before surgery, 2 mL mepivacaine 20 mg/mL adrenaline 1:100,000 was infiltrated in the area of the Spix spine or in the vestibular/palatal oral mucosa. For totally or partially impacted molars, a buccal total thickness trapezoidal flap was raised when necessary.
Ostectomy and tooth sectioning phases were performed using diamond or Allport burs on a low-speed handpiece (30,000 rotations/min), in all cases irrigated with sterile physiological solution. The gingival tissues were closed with silk suture. After surgical curettage, hDFCs were placed in DMEM (Life Technologies, Italy), washed in DMEM supplemented with an antibiotic/antimycotic mix (Life Technologies), and either maintained in culture in the RCCS bioreactor (Synthecon) or used for cell isolation. For all patients, independent of the anesthetic applied, antibiotic and anti-inflammatory medication was prescribed when necessary (usually amoxicillin 2 g or clarithromycin 500 mg, twice daily for 5 days, plus ibuprofen 600 mg, twice daily for 3 days), together with 0.2% chlorhexidine rinses twice daily for 7 days.
3D culture of hDF tissue
Intact hDF, or portions thereof obtained by dividing larger portions into smaller fragments, was cultured. Tissue samples were maintained in 3D culture in the RCCS bioreactor (Fig. 1) in the presence of DMEM supplemented with 10% fetal bovine serum (FBS) and antibiotic/antimycotic mix (complete medium). Medium was refreshed twice weekly.
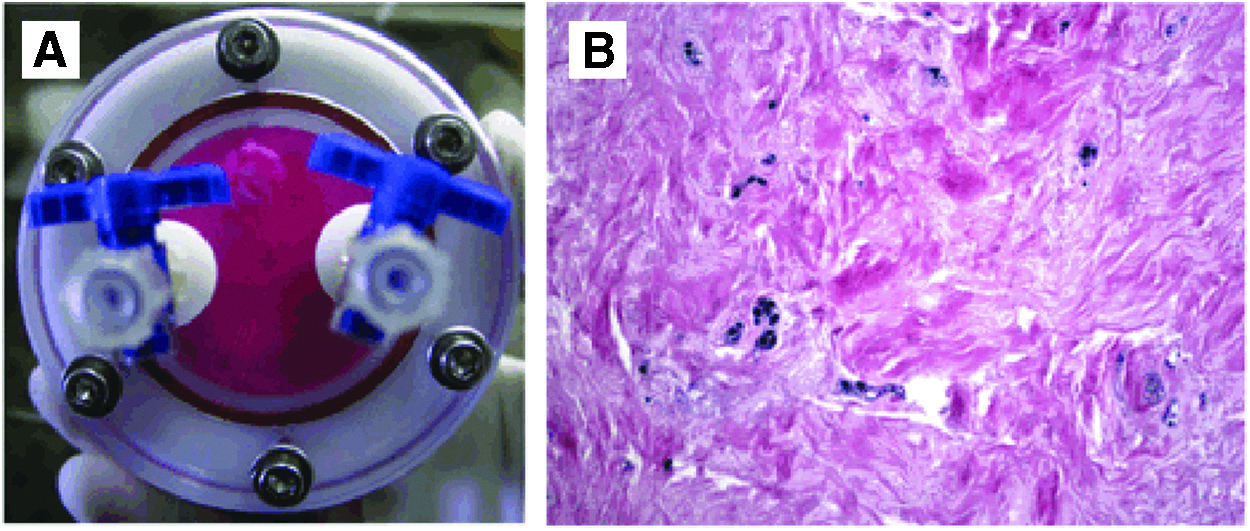
Dynamic 3D culture of follicle explants in the RCCS™ bioreactor.
Isolation of hDF mesenchymal stromal cells
Briefly, under sterile conditions the sample was washed in 1× PBS supplemented with penicillin–streptomycin solution (1 × ; Euroclone, UK). The sample was minced with surgical scissors and then transferred in 15 mL conical tube containing a solution of type I collagenase (0.1% in 1× PBS +0.2% bovine serum albumin, reagents supplied by Life Technologies) with a volume ratio of 3:1 (solution/tissue). The sample was kept at 37°C under stirring condition for 3 h and then centrifuged (300 g, 10 min). The pellet was washed in 1× PBS+0.2% BSA and, after an additional spinning, resuspended in DMEM high glucose medium supplemented with 10% FBS, 2 mM L-glutamine, and penicillin–streptomycin solution (Euroclone). The isolated cells were counted and their viability assessed by Trypan blue (Sigma Aldrich) exclusion. Then cells were seeded in 25 cm2 flask (Euroclone) and maintained at 37°C, 5% CO2. Cell proliferation is daily monitored by inverted microscope (Leitz DIAVERT®), and just plastic-adherent cells were grown for further studies.
Mesenchymal stromal cell phenotypic characterization
The primary culture thus obtained was characterized by several parameters: proliferation rate, clonogenicity (colony forming unit-fibroblast [CFU-F]), and expression of the typical mesenchymal stromal cell (MSC) markers.
Cell proliferation
At early passages (from II to V) cells were grown, and MSCs of hDF doubling time (DT) were calculated as follows: DT = t × ln2/ln (Nc/Np), were Np is the number of plated cells, Nc is the number of counted cells, and t is the time in culture expressed in hours.
CFU-F assay
hDFCs (passage I–III) were plated at low density by 1:2 serial dilution (starting from 500 cells/3,5 cm2) in control medium supplemented with 20% FBS. After 6 days medium was changed and at day 14 cells were washed, fixed with 100% methanol, and stained with 2 mg/mL Crystal Violet (Fluka). The clonogenic potential (%) was established by scoring individual colonies (consisting of at least 20 cells) with respect to the number of seeded cells.
Isolation and culture of hDF precursor cells (hDFCs)
hDFCs were obtained either after enzymatic release from intact hDF or after spontaneous outgrowth from hDF tissue. For enzymatic isolation, minced hDF tissues were digested for 2 h in 0.1 U/mL collagenase type I-1 U/mL dispase (Roche Mannheim, Italy) solution, at 37°C. Cells were seeded at 2 × 104 cells/cm2 onto plastic dishes in complete medium. The following day, the culture medium was replaced with fresh medium. Alternatively, minced hDFCs were placed in culture on plastic dishes in complete medium, and adherent cells were left to outgrow for 10 days. hDFCs isolated by means of either technique were amplified and used for experimental purposes up to the 18th passage.
Immunoselection for CD90+, CD146+, and CD14− hDFCs
hDFCs were positively selected for expression of CD90 or CD146 Ags, using the MACS® Separation Kit with the specific CD90 or CD146 microbeads (Miltenyi Biotec Ltd., Italy) and following the manufacturer's instructions. The HepG2 cell line (ISZLER Cell Repository, Italy) was used as a negative control to assess the specificity of cell binding to the different Abs-labeled microbeads (less than 0.2% of HepG2 cells were retained in the columns). Moreover, the low number of hDFCs lost during the entire selection procedure (only 9.0% ± 1.8% of cells) confirmed the reliability of the procedure. hDFCs expressing both CD90 and CD146 Ags were further separated using the MACS Separation Kit with the specific CD14 microbeads. Cells not retained by the specific anti-CD14 Ab were collected and counted to evaluate the percentage of CD14-negative versus positive cells.
CFU-F assay
The CFU-F assay was performed by plating hDFCs at 0, 1, 10, 100, 500, or 1000 cells/35 mm-dish in complete medium. After 14 days of culture, cells were stained with May-Grunwald & Giemsa stains.
Proliferation assays
Cells were plated at a density of 3 × 104 cells/cm2 in complete medium and counted daily with a hemocytometer. Cell viability was assessed by the dye-exclusion assay (Trypan blue 0.4%, 1:1). Growth curves were produced in triplicate.
Induction of cell differentiation
For osteogenic, chondrogenic, and adipogenic differentiation assays, hDFCs were seeded at a density of 2 × 103, 2 × 104, or 3 × 104 cells/cm2, in alpha-MEM (Lonza, Italy), HAMF12, and DMEM, respectively. All culture media were supplemented with FBS and antibiotic/antimycotic mix. Twenty four hours after seeding, cells were exposed to osteogenic medium (alpha-MEM, including 15% FBS, 50 mg/mL L-ascorbate-2-phosphate, 0.01 μM dexamethasone, and 10 mM glycerol 2-phosphate) for 14 days. Alternatively, cells were cultured in adipogenic medium (HG-DMEM, with 10% FBS, 10 μM dexamethasone, 10 μg/mL insulin, 0.5 mM 3-isobutyl-1-methylxanthine, and 200 mM indomethacin) for 3 days, after which the adipogenic medium was replaced with HG-DMEM, supplemented with 10% FBS and 10 μg/mL insulin for 11 days. Control hDFCs were stored in complete DMEM with neither osteogenic nor adipogenic supplements. For chondrogenic induction, the cells were cultured for 21 days in RCCS 12 in the HARV™ culture chamber, in the presence of HAM:DMEM (v:v) supplemented with 10% FBS, 0.1 μM dexamethasone, 5 μg/mL L-ascorbate-2-phosphate, 100 μg/mL sodium pyruvate, 10 mg/mL ITS, and 10 ng/mL transforming growth factor-β1. Culture media was changed twice weekly.
Histochemical study of hDF tissues
hDF samples from freshly harvested tissue or from 3D-cultured explants were fixed in 10% buffered formalin, embedded in paraffin, and then processed for histology and immunohistochemistry procedures. 5 μ hDF samples from freshly harvested eosin & hematoxylin solution (E&H) were processed for cell/tissue morphological analysis.
The response of hDFCs to the differentiating media was assessed by Alizarin red/Von Kossa, Alcian blue, or oil red staining, specific to identify osteogenic, chondrogenic, and adipogenic lineages, respectively. For isolated cells cultured in the RCCS, samples were embedded as described elsewhere. 19
Immunohistochemical characterization of hDF explants
After deparaffinization and alcoholic rehydratation, 5 μ of deparaffinized was exposed to 3% hydrogen peroxidase to quench endogenous peroxidase. Bond Polymer Refine Detection (Leica Biosystems, UK) was used on the Bond-Max automated immunostainer (Leica Microsystems, UK). The following primary antibodies were used to characterize hDF components: anti-CD31 (1:50; Novocastra Laboratories, UK), anti-CD34 (1:200; Novocastra Laboratories), anti-CD44 (1:50; Novocastra Laboratories), anti-CD45 (1:70; Novocastra Laboratories), anti-CD90 (1:100; Abcam, UK), anti-CD146 (1:25; Novocastra Laboratories), anti-CD105 (1:50, Monosan; Sanbio B.V., the Netherlands), anti-smooth muscle actin (SMA) (1:300, Thermo Scientific; Thermo Fisher Scientific, UK), and anti-pancytokeratin (1:300; DakoCytomation, Denmark). Staining was performed with 3,3′-Diaminobenzidine tetrahydrochloride and hematoxylin counterstaining to visualize the cell nuclei. Double immunostaining was performed with anti-CD90 and anti-CD146 Abs, using Bond Polymer Refine Red Detection system for the former and Bond Polymer Refine Detection system for the latter.
Results
3D culture of hDF tissue explants
The innovative 3D culture method described in this study, based on the use of the dynamic RCCS (Fig. 1), was applied successfully to hDF tissue explants, as demonstrated by characterization of the cultured samples vs. freshly isolated counterparts (Fig. 2). In both cultured and freshly explanted tissues, E&H staining evidenced the original nervous and vascular structures and the connective tissue. In situ immunostaining of the 3D-cultured explants confirmed maintenance of smooth muscle cells at the perivascular location (anti-SMA Ab) and close to the mature vascular structures (anti-CD31 Ab). The presence of neovascularization processes (anti-CD34 Ab) demonstrated cell health. However, compared to their freshly isolated counterparts, the 3D-cultured hDF tissue showed more CD31- and CD34-positive elements. Keratinized cells were also evidenced in both freshly harvested and 3D-cultured samples (anti-pancytokeratin Ab), probably in areas where the normal resorption of Hertwig's epithelial root sheath/epithelial rests of Malassez had not fully occurred.
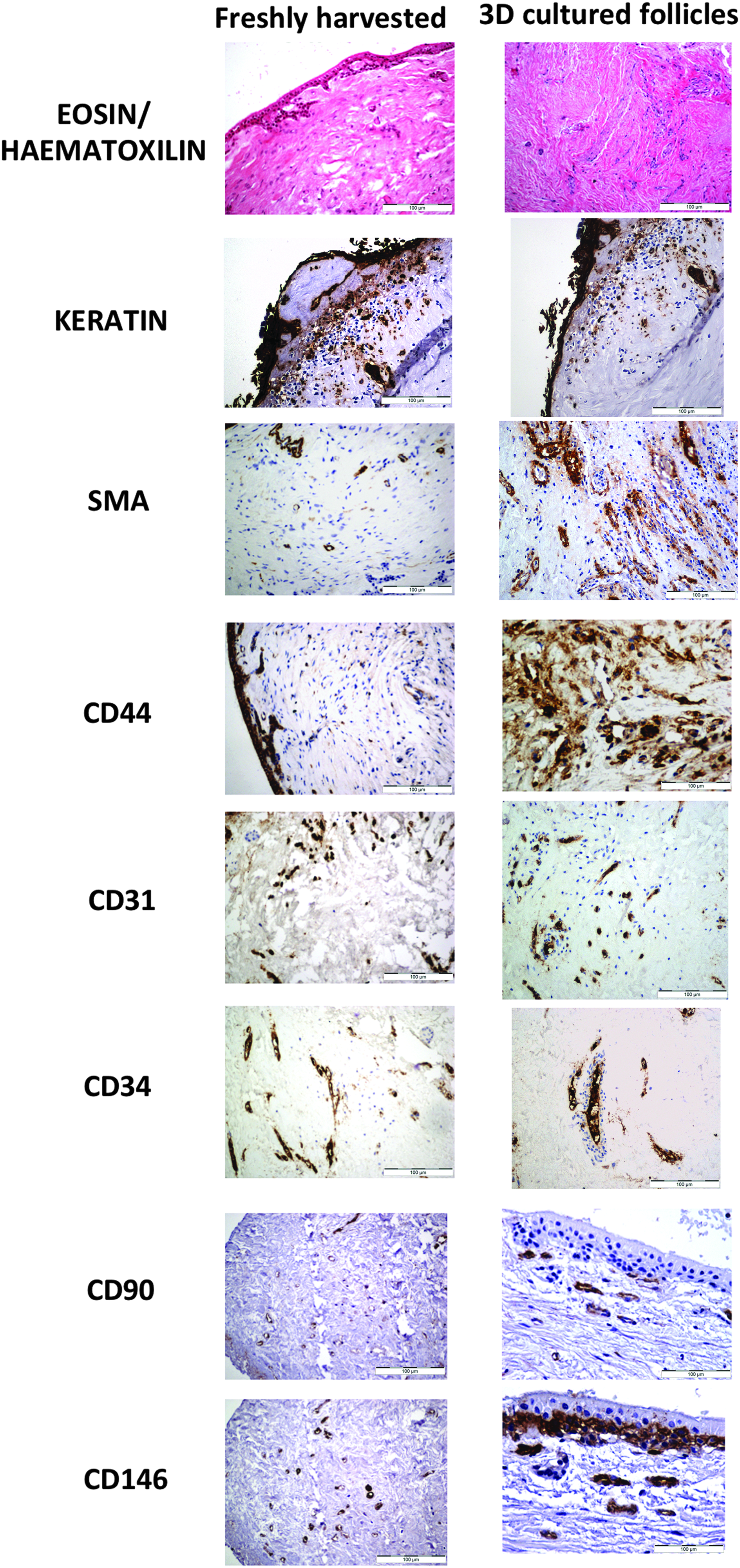
In situ immunomorphological characterization of freshly harvested and 3D-cultured hDF. Samples from freshly isolated and 3D-cultured hDF were stained with E&H and subjected to in situ immunodetection of the indicated markers for different cell types (initial magnification: 200 × ). The 3D tissue culture was maintained for 10 days in fluid-dynamic conditions (RCCS bioreactor). Pancytokeratin, SMA, CD44, CD31, CD34, CD90, and CD146 were used as markers to characterize epithelial, muscular, mesenchymal, and endothelial tissues and potential precursor cells. SMA, smooth muscle actin.
Well-organized odontogenic epithelium was observed at the periphery of some hDF explants. In the 3D-cultured hDFCs, E&H staining evidenced rare, small round cells at the perivascular location. Immunocharacterization of these cells showed that they were negative for all markers used (SMA, CD31, CD34) except for CD44. Thus three different populations of cells were identified by expression of stem cell markers CD90 and CD146: cells expressing only CD90, those expressing only CD146, and those expressing both Ags. In nervous structures, for example, cells expressed either CD90 or CD146 markers. In vascular structures, CD90-positive staining was generally more marked than CD146. In the stratified epithelium, the basal stratum of cells only expressed CD146 Ag. Cells expressing both markers were found to be mostly located in the vascular/perivascular areas, although rare cells were also seen in the connective tissue.
3D culture of hDFCs
The initial cell population isolated from hDFCs was tested for the expression of stemness markers (CD44, CD90, CD146, CD105, CD31, CD34, and CD45), and in situ immunocytochemistry demonstrated expression of all markers except CD31, CD34, and CD45 Ags (Fig. 3A). CD90, CD44, and CD105 were broadly expressed, while CD146 expression was more restricted (about 30% of all cells). After authentication of these cells as potential hDF precursors, isolated cells were further subjected to immunoselection by sequential elution, first on MS columns loaded with anti-CD90-labeled microbeads and then on MS columns loaded with anti-CD146-labeled microbeads.

Characterization of the whole cell population isolated from intact hDF.
Figure 3B shows that about 33% of cells in the initial highly heterogeneous population (P2) expressed CD90 (CD90+ cells) and, of these CD90+ cells, about 37% also expressed CD146 (CD90+/CD146+ cells). These double-CD-positive cells accounted for almost 12% of cells in the initial population. Moreover, sequential elution, performed first on MS columns loaded with CD146-labeled microbeads and then on MS columns loaded with CD90-labeled microbeads, indicated that about 15% of cells from the initial population co-expressed both CD146 and CD90 markers. Elution of these CD146+/CD90+ cells on MS columns loaded with anti-CD14 labeled microbeads demonstrated that these cells did not express CD14 Ag (number of positive cells <0.1%).
Proliferation and capacity to form colony
The different hDFC populations were evaluated for their capacity to form colonies when seeded at low density. Figure 4 provides an overview of cell behavior after seeding at densities of 1, 10, 100, 500, or 1000 cells/35-mm dish. Interestingly, CD146+/CD90+ cells formed more and bigger colonies than did CD90+/CD146+ cells.
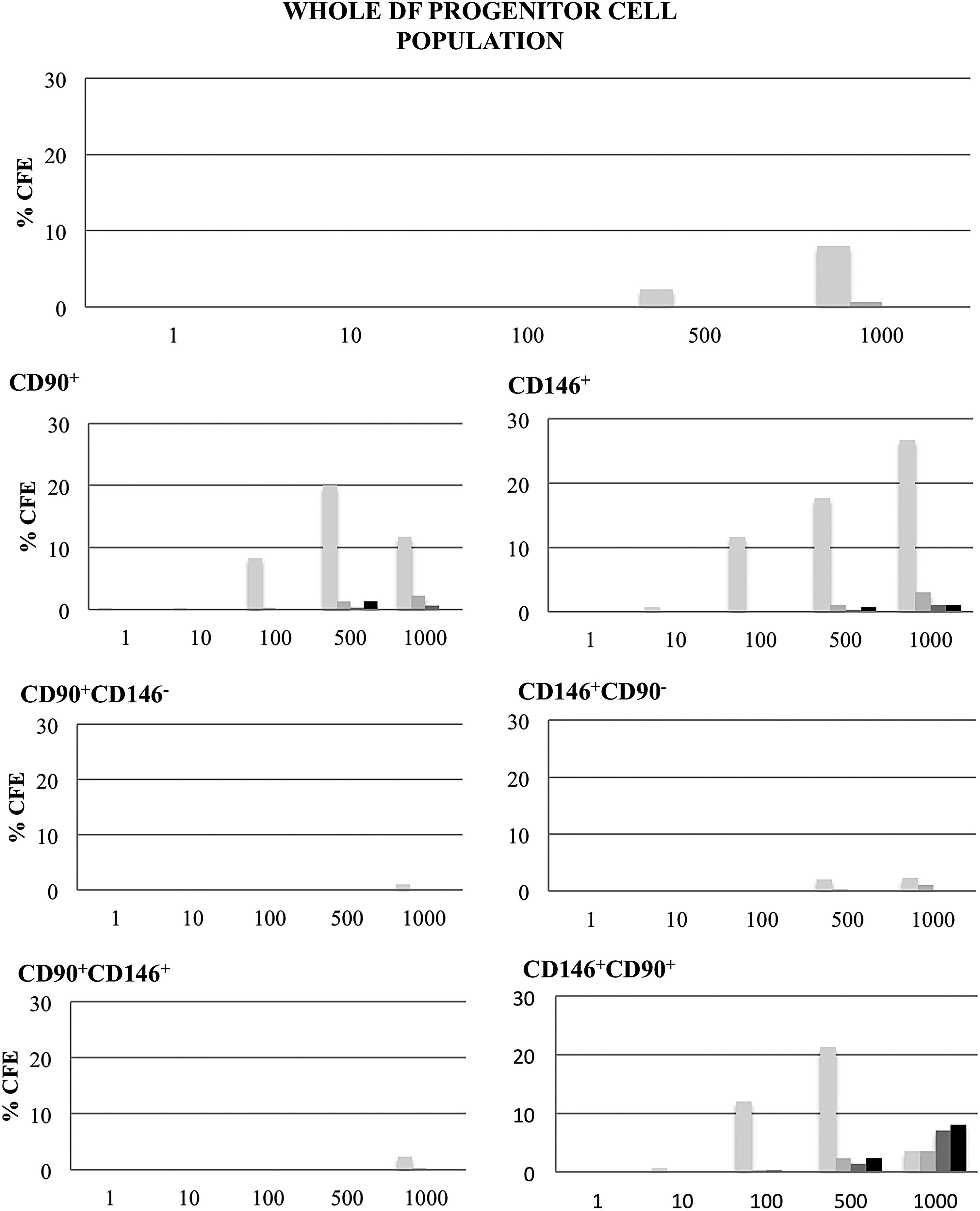
Self-renewal capacity of hDF-derived cell populations. The CFU-F assay was used to evaluate the capacity of the different hDF-derived cell populations to give rise to colonies at different seeding densities (from 1 to 1 × 103 cells/35-mm Petri dish). CFU-F, colony forming unit-fibroblast.
The quantitative determination of CFU-F data, calculated in terms of colony number per seeding density (percentage of colony forming efficiency [% CFE]) and colony size (average number of cells/colony), is shown in Figure 5. While the initial population of isolated cells presented a limited capacity to form colonies (low number of colonies, even at high seeding densities), the CD90+ or CD146+ selected cells had a greater capacity to renew themselves. CD90+/CD146+ cells were less capable of forming colonies than CD146+/CD90+ cells, the latter having the greatest proliferative capacity of all cell populations. The difference in cell growth capacity between the different cell populations (CFU-F assay) was confirmed by analysis of cell growth rates: CD146+/CD90+ cells grew faster than did CD90+/CD146+ cells (doubling the population in about 23 and 26 h, respectively) and reached the highest cell density at confluence.
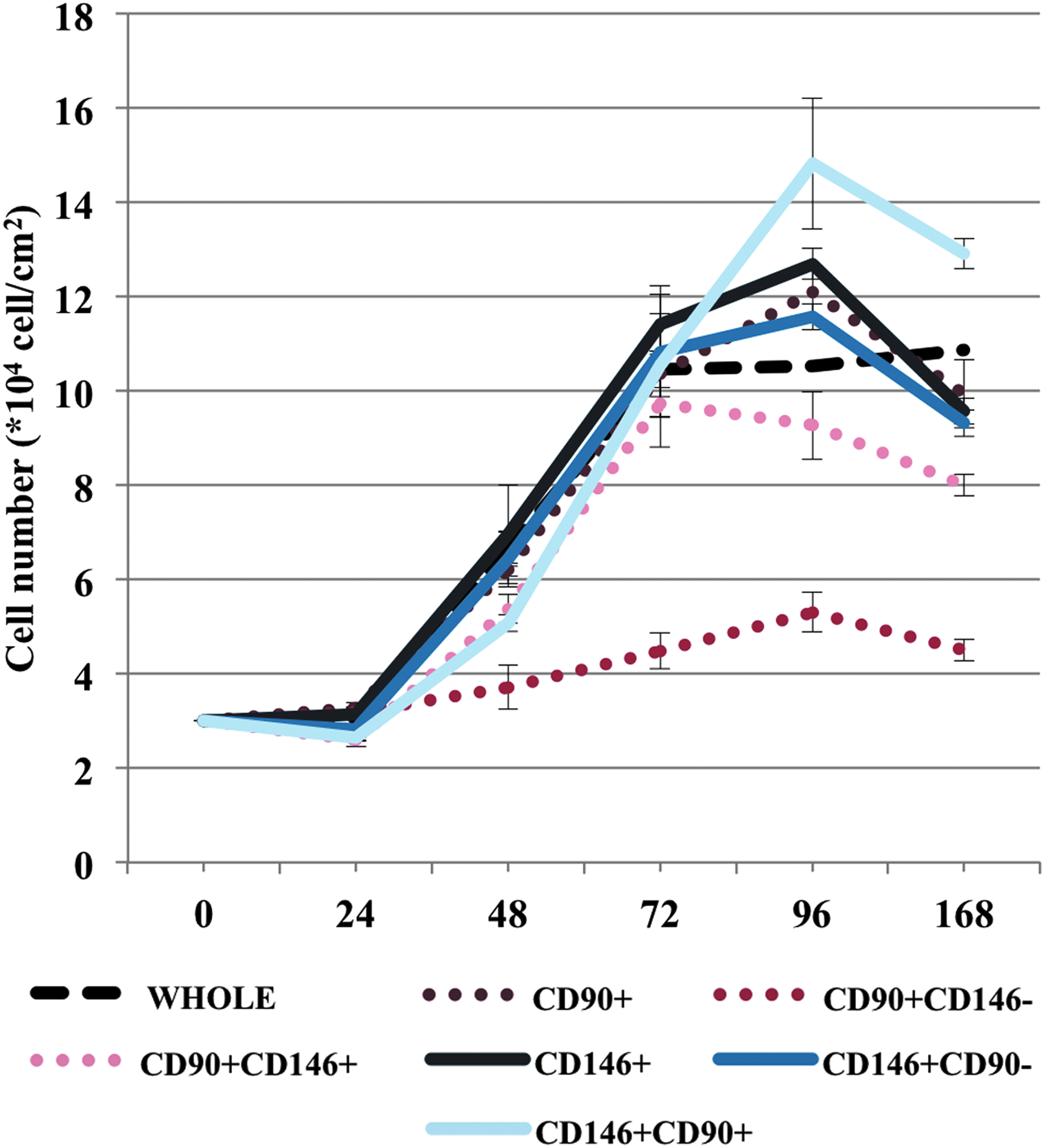
CFU-F assay on different cell populations from hDF. CFU-F colonies were optically counted; for large colonies (>2000 cells/colony), cell number was assessed by analysis of several micrographic images. Cells were plated at a density of 3 × 104 cells/cm2 in complete medium and counted daily with a hemocytometer. Cell viability was assessed by the dye-exclusion assay (Trypan blue 0.4%, 1:1). Growth curves were produced in triplicate. The % CFE expresses the ratio between the number of colonies formed and the number of plated cells × 100. % CFE, percentage of colony forming efficiency.
Capacity to differentiate
The multipotency of hDFCs was then evaluated by assessing their capacity to differentiate into osteogenic or adipogenic precursors after 2 weeks of 2D culture in dishes. All cell populations were able to differentiate into osteoblast-like cells (Von Kossa and Alizarin red staining), although the double-CD positive cells were slightly more able to accumulate mineral matrix, compared to the initial population (Fig. 6). In the adipogenic medium, the cells changed morphology and weakly accumulated lipid droplets (oil red staining).
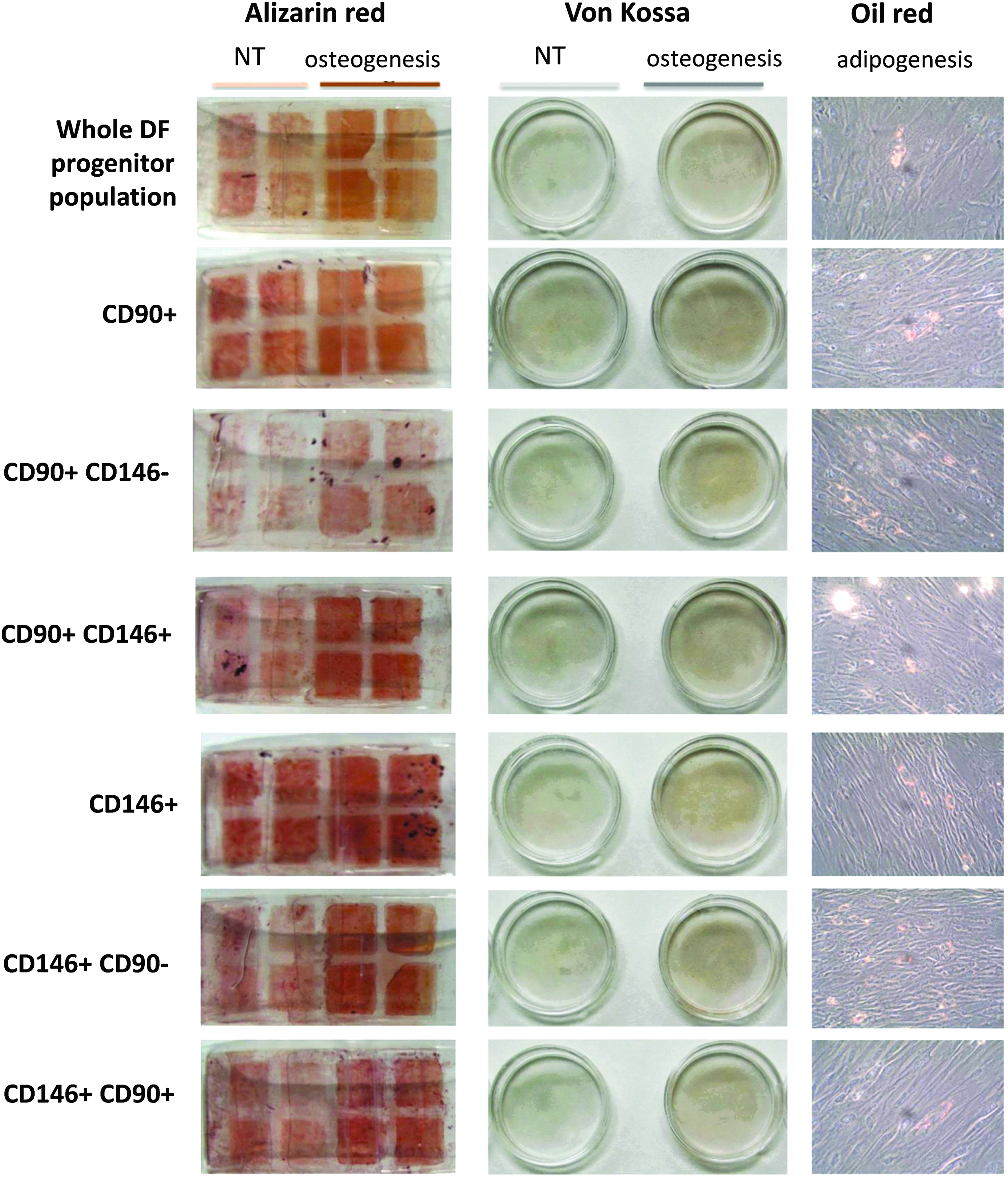
Multipotency of dental progenitor cells. The capacity of progenitor cells to differentiate into osteogenic or adipogenic lineages was assessed in static 2D culture conditions. Depending on the lineage, cells were cultured in specific media, as described in Materials and Methods section. Photograms were taken with an Olympus CAMEDIA C-5050 zoom digital camera. 2D, two-dimensional.
For the dynamic 3D culture, hDFCs were first amplified in monolayer and then injected into the RCCS culture chamber. Cells promptly began to aggregate but, depending on the medium used, multicellular aggregates varied in size and in ECM content. In particular, cells induced to differentiate into the adipogenic lineage produced very little ECM, and cell aggregates were smaller than those observed in chondrogenic- or osteogenic-induced cells.
In comparison with the 2D culture, the dynamic 3D culture markedly favored accumulation of proteoglycans, lipids, or mineral foci in hDF precursors exposed to chondrogenic, lipogenic, or osteogenic medium, respectively (Fig. 7).
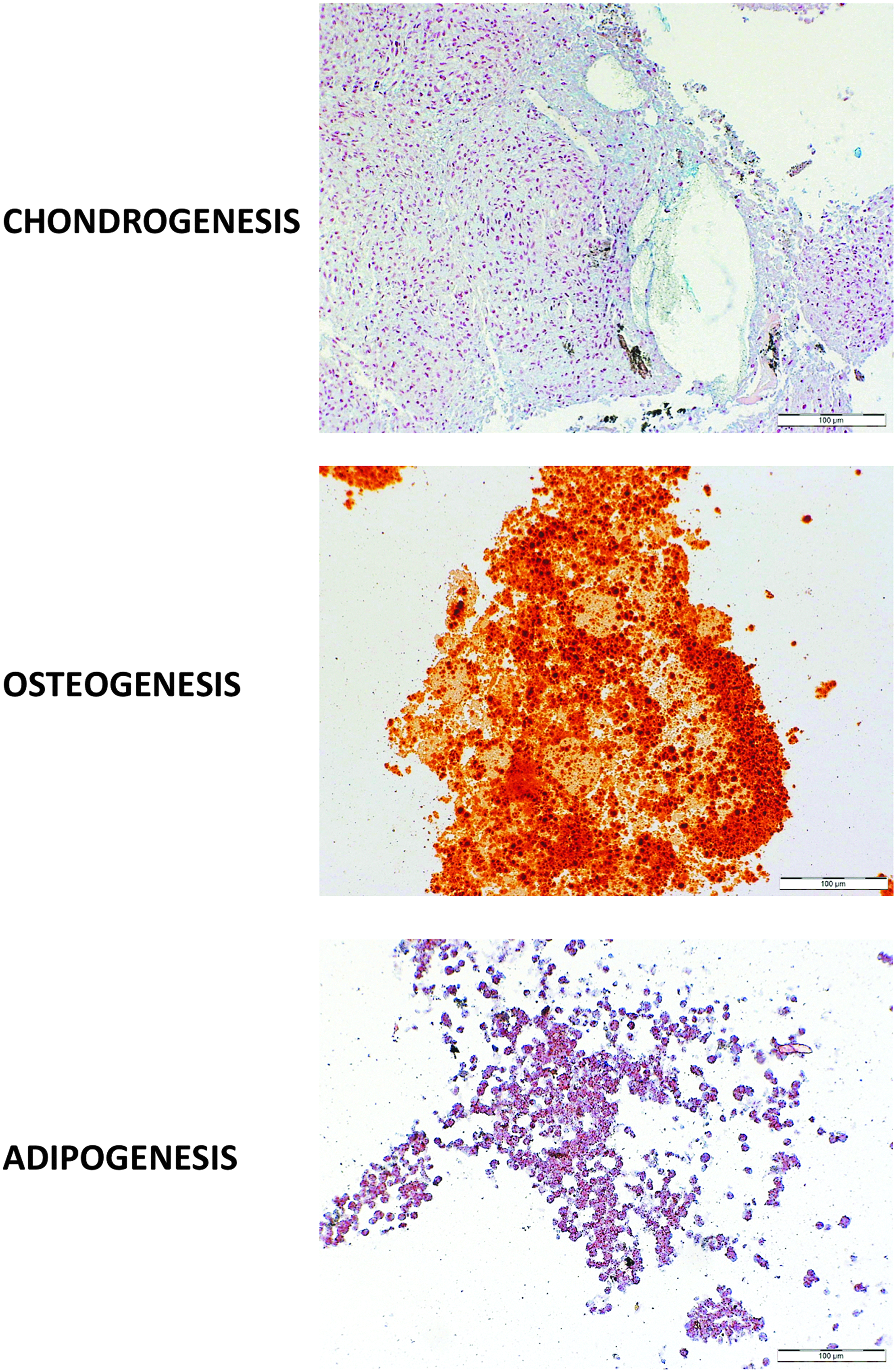
Multipotency of hDF-derived cells maintained in dynamic 3D culture conditions. Cells cultured in the RCCS bioreactor were maintained in different culture media that, specifically, favor the differentiation of cells into chondrocytes, osteoblasts, or adipocytes, as described in Materials & Methods section. Samples were treated with Alcian blue, Alizarin red/Von Kossa, or oil red staining to identify their chondrogenic, osteogenic, or adipogenic potential, respectively.
Discussion
The present study characterized two 3D ex vivo models of hDF, derived either from follicular tissue explants or from hDF-isolated cells. Both models were based on the use of the RCCS dynamic bioreactor; the first provided long-term preservation of the original structural and biochemical characteristics of tissue-specific microenvironments (tissue culture), while the second led to neogenesis of appropriate contexts, suitable for cell survival and expression of differentiated phenotypes (cell culture),20,21 or to reproduce specific 3D tissue morphogenesis. 11
The first model applied the RCCS-based culture method to healthy hDF explants, maintained for up to 10 days in 3D dynamic culture. The physiological relevance of this hDF explant-based model was confirmed by the maintenance of the original tissue composition, organization, and architecture, showing the presence of the characteristic cell types (neuronal, endothelial, epithelial, and myofibroblastic cells) of the in vivo condition (freshly harvested hDF). The expression of differentiation markers, typical of normal hDFCs, also demonstrated that, in the cultured tissue, cell components maintain their viability and differentiated status, thus confirming that preservation of the original, tissue-specific microenvironment is essential to sustain cell homeostasis, as has already been demonstrated for other cell types.11,16,22 Moreover, maintenance of the original tissue features appears also to favor neovascularization of the cultured tissue, as suggested by the increased expression of CD31 and CD34 markers, specific for endothelial progenitors and/or mature cells.
Taken together, the findings show that the innovative tissue culture system for hDF explants presented in this study might be a powerful tool for a number of investigative approaches, including the study of dental follicle behavior (homeostatic/regenerative processes) and follicle response to various physiopathological and pharmaco-toxicological stimuli (e.g., new biomaterials, dental implants, and drugs used for regenerative dentistry and implant therapy).
Another significant outcome of the study is that, among the heterogeneous cell populations present in the 3D-cultured hDF explants (namely, the abundance of specific markers detected), dental stem/progenitor cells were also detected. Although such precursors in hDF have already been reported in isolated cells,1,23 the precise location of these cells in situ is determined in this study. The majority of hDF precursors were detected in perivascular areas. These cells were small in size and expressed CD44 (but not cytokeratins), CD31, and CD34; they expressed the SMA Ag very weakly. Their specific location and the pattern of expressed markers suggest that these cells could be stromal progenitors and/or pericyte-derived cells.24,25
The second model of hDF is based on the 3D culture of hDFCs. These cells express CDs specific to MSC precursors (CD90, CD146, CD105, and CD44) but not markers of hematopoietic precursors (CD34) or mature cells (CD31, CD14). Analysis of the hDFC proliferation rate also confirmed these cells' self-renewal capability. Interestingly, depending on the procedure used for the double selection for CD90 and CD146 expression (CD90+/CD146+ or CD146+/CD90+), it was found that the two hDFC populations behave differently, although they are identical in regard to CD expression. This observation, in addition to confirming that hDF consists of a heterogeneous cell population, 8 shows the importance of the technique used for isolating/selecting specific cell types.
Furthermore, the results indicate that the hDF tissue may encompass a mixture of precursors, whose phenotypes could vary from undifferentiated/uncommitted stem cells to more committed precursors6,8,26 to pericyte-originating cells. 27 When progenitors were isolated from different donors, the proportion of cells expressing CD90 and CD146 varied greatly, as did their growth properties and sensitivities to differentiating media. These data suggest that, when tooth-derived cells are considered, in addition to the age of the donor, the maturation state of the dental follicle may also give rise to differently competent cells. In addition, hDFCs maintain multipotency, since they were able to progress toward osteogenic and adipogenic lineages, even if the number of cells able to differentiate into adipocytes was limited to sparse areas.
Conversely, the fluid-dynamic 3D microenvironment provided by the RCCS boosted the specific expression of the differentiated osteoblastic/adipocytic phenotypes more markedly. This finding was even more evident for cells exposed to the osteogenic medium, confirming the greater propensity of progenitors isolated from hDF to differentiate into bone-specialized cells. Moreover, besides guaranteeing optimal conditions for osteoblastic/adipocytic differentiation, the 3D cell culture also provided the necessary conditions to guide hDFC differentiation toward the chondrogenic lineage. In contrast to reports by Honda et al.8,28 in the case of DFCs, and by Balic 29 for dental pulp, in the present experimental conditions, cells also differentiated into chondrogenic cells, suggesting that these isolated hDFCs were in a less committed state than those used in the studies cited.
We observed a marked expression of cartilage/bone-matrix components on the RCCS-based culture and a weaker tendency of hDFCs to follow the adipogenic lineage. This could be explained simply by the “history” of these cells, which, in the hDF, are normally fated to generate the periodontium and not the adipose tissue. This predisposition to differentiate into a specific cell type, depending on the origin of the tissue, has already been demonstrated for bone marrow-derived, adipose tissue-derived, and dental-derived stem cells.30–33
Conclusions
In conclusion, both of the fluid-dynamic, RCCS-based, 3D ex vivo models of hDF presented in this study provide new opportunities to investigate hDF features in the fields of basic and applied research. The tissue-based model, by preserving the tissue peculiarity in toto, may provide a powerful tool for studying hDF physiopathology and response to therapeutic agents. The hDF precursor-based 3D model could be used in these contexts as well. In addition, because this model is based on the culture of hDF precursor cells, it could also be used to study tooth development and novel tissue-engineering strategies, designed for dental/periodontal tissue regeneration.
Footnotes
Acknowledgments
The authors thank Franco Alpi, Irene Scarabelli, and Lucia Fontana (Pathological Anatomy Unit, University of Brescia, Italy) for their technical assistance. The authors also acknowledge and thank Dr. William Giannobile (University of Michigan, Ann Arbor, MI) for the excellent critical reviewing of the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.