
Abstract
Fabrication of composite scaffolds is one of the strategies proposed to enhance the functionality of tissue-engineered scaffolds for improved tissue regeneration. By combining multiple elements together, unique biomimetic scaffolds with desirable physical and mechanical properties can be tailored for tissue-specific applications. Despite having a highly porous structure, the utility of electrospun fibers (EF) as scaffold is usually hampered by their insufficient mechanical strength. In this study, we attempted to produce a mechanically competent scaffold with cell-guiding ability by fabricating aligned poly lactic-co-glycolic acid (PLGA) fibers on decellularized human amniotic membrane (HAM), known to possess favorable tensile and wound healing properties. Decellularization of HAM in 18.75 μg/mL of thermolysin followed by a brief treatment in 0.25 M sodium hydroxide efficiently removed the amniotic epithelium and preserved the ultrastructure of the underlying extracellular matrix. The electrospinning of 20% (w/v) PLGA 50:50 polymer on HAM yielded beadless fibers with straight morphology. Subsequent physical characterization revealed that EF-HAM scaffold with a 3-min fabrication had the most aligned fibers with the lowest fiber diameter in comparison with EF-HAM 5- and 7-min scaffolds. Hydrated EF-HAM scaffolds with 3-min deposition had a greater tensile strength than the other scaffolds despite having thinner fibers. Nevertheless, wet HAM and EF-HAMs regardless of the fiber thicknesses had a significantly lower Young's modulus, and hence, a higher elasticity compared with dry HAM and EF-HAMs. Biocompatibility analysis showed that the viability and migration rate of skeletal muscle cells on EF-HAMs were similar to control and HAM alone. Skeletal muscle cells seeded on HAM were shown to display random orientation, whereas cells on EF-HAM scaffolds were oriented along the alignment of the electrospun PLGA fibers. In summary, besides having good mechanical strength and elasticity, EF-HAM scaffold design decorated with aligned fiber topography holds a promising potential for use in the development of aligned tissue constructs.
Introduction
N
Electrospun fibers (EF) have been used in a variety of applications, including the fabrication of tissue-engineered scaffolds. Using the electrospinning technique, ultrafine fibers with highly porous structure that resembles native extracellular matrix (ECM) can be produced. To date, many studies involving the regeneration of musculoskeletal,2,3 cardiac,4,5 and nerve tissues6,7 are using nanofibers as a carrier to deliver cells or therapeutic agents to the injury sites. Nanofibrous scaffolds possess many favorable properties such as large surface-to-volume ratios, high porosity, tunable fiber size, and orientation, as well as unique 3D structures mimicking the architecture of natural ECM. 8 Previous studies have demonstrated the biocompatibility of electrospun nanofibers with various types of cells and their ability to regulate cellular morphology, orientation, and proliferation, as well as the production of essential secretory proteins.9–12
Biological scaffolds composed of the naturally occurring ECM have the ability to modulate tissue healing and remodeling processes for functional recovery of injured tissue. Unlike synthetic scaffolds, Biological scaffolds are biologically active, biodegradable, and enriched by ECM proteins and bioactive molecules that can improve tissue healing and repair. Human amniotic membrane (HAM) is a naturally occurring scaffold used in a variety of clinical applications, especially those that relate to ocular surface reconstruction and wound management. It has unique biological and mechanical qualities that render it desirable as one of the potential scaffolding materials. Amniotic membrane patching against skin wounds,13,14 ocular surface disorders,15,16 infarcted hearts,17–19 and liver fibrosis 20 had been shown to support the healing processes as it is lowly immunogenic, anti-inflammatory, and anti-fibrotic, and releases soluble factors essential for cell growth and differentiation.20–24 HAM adheres well to wound while maintaining a moist microenvironment at the site of injury. Although HAM is relatively thin, it is sufficiently strong and elastic making it a suitable candidate biomaterial for tissue-engineered scaffold.
Creating an engineered scaffold that is mechanically competent and topographically favorable for cell alignment is crucial for the regeneration and remodeling processes of highly organized tissue. While the utility of electrospun nanofibers is mostly hampered by the weak mechanical strength and poor hydrophilicity of the scaffolds unless they are modified,25–27 HAM lacks the ability to guide cells in a desirable orientation during the formation of tissue substitutes. On the contrary, fabricating a composite scaffold composed of both HAM and aligned nanofibers can provide sufficient mechanical strength and alignment to the scaffold, respectively. In this study, we explore the potential utility of a novel composite scaffold, namely the EF-HAM, for tissue regeneration through the fabrication of electrospun poly lactic-co-glycolic acid (PLGA) fibers on decellularized HAM. The physical and mechanical properties of EF-HAM scaffold were characterized, and scaffold biocompatibility and potential of cell alignment for skeletal muscle cells were evaluated.
Materials and Methods
This study obtained ethics approval from the Research Ethics Committee, The National University of Malaysia (UKM 1.5.3.5/244/02-01-02-SF1284).
Collection, processing, and preservation of HAM
HAMs were procured with consent from healthy mothers undergoing cesarean sections at the National University of Malaysia (UKM) Medical Centre. Only donors without underlying medical problems and obstetric complications were included in this study. HAMs were handled aseptically and immediately processed after retrieval. Each amniotic membrane was separated from adjacent layer namely the chorion through blunt dissection. HAM was washed extensively in ultrapure water until all debris, mucus, and blood residues were removed and soaked with moderate shaking in 0.05% sodium hypochlorite (NaClO) solution for 1 h. HAM was washed 3 × 15 minutes by moderate shaking in Dulbecco's phosphate-buffered saline (DPBS; Thermo Fisher Scientific) to remove excess NaClO. Processed HAMs were placed in 20 mL DPBS with 1% antibiotic/antimycotic (AA; Thermo Fisher Scientific) solution and freeze-preserved at −80°C until further use.
HAM decellularization
Processed HAMs were cut into smaller pieces with size of 3 × 3 cm each and decellularized using thermolysin (TL; Sigma-Aldrich) and sodium hydroxide (NaOH; BDH Laboratory Supplies, England) to remove the epithelium from the membrane. TL working solutions at 12.5, 18.75, and 25.0 μg/mL of concentration were preheated at 60°C for 10 minutes before treatment. HAMs were incubated in the respective TL working solutions at 37° for 10 mins and subsequently pulse vortexed for 60 times. HAMs were later washed 3 × 15 mins by moderate shaking in DPBS to remove excess TL. HAMs were then soaked in 0.25 M NaOH for 1 min, immediately pulse vortexed for 30 times, and washed 3 × 15 mins by moderate shaking in DPBS. HAMs were later air-dried under biosafety cabinet. Three biological samples were randomly selected for ultrastructural analysis of HAM by scanning electron microscopy and 4′,6-diamidino-2-phenylindole (DAPI) staining on HAM to confirm decellularization of the tissue. The remaining air-dried HAMs were sent to the Malaysian Nuclear Agency for gamma sterilization. Sterilized HAMs were stored at room temperature in a dry storage area and used for EF-HAM scaffold fabrication and mechanical testing on HAM.
Visualization of amniotic epithelium on decellularized HAM
Decellularized HAMs were counterstained with DAPI (Thermo Fisher Scientific) to evaluate the effectiveness of the TL-NaOH decellularization process. Briefly, decellularized HAMs were fixed with 4% paraformaldehyde (Sigma-Aldrich) at 4°C overnight and were later washed three times in DPBS. Samples were incubated with DAPI (1:20,000 dilution) for 40 mins at room temperature and were washed three times in DPBS. Images of the HAMs were captured using fluorescence microscope (Nikon, Japan) to assess the presence of remaining epithelial cells on decellularized amniotic membranes.
Fabrication of composite EF-HAM scaffolds
PLGA EF were fabricated with different thicknesses on decellularized, gamma-sterilized HAMs via electrospinning to produce composite EF-HAM scaffolds. PLGA 50:50 polymer (DURECT) was dissolved at 20% (w/v) concentration in a 7:3 solvent mixture of dichloromethane (Sigma-Aldrich) and dimethylformamide (Sigma-Aldrich), loaded into a 1-mL syringe equipped with a 21-gauge blunt needle and spun onto decellularized HAMs attached on a mandrel collector at ∼1000 rpm of rotating speed, 15 kV of applied voltage, 0.3 mL/h of polymer flow rate, and 15 cm of deposition distance. Different fiber thicknesses on HAMs were created by varying the deposition times to 3, 5, and 7 mins. The resulting EF-HAM scaffolds were air-dried under biosafety cabinet to allow for complete solvent evaporation and UV-irradiated for 30 min. Three biological samples were randomly selected for ultrastructural analysis and mechanical characterization of EF-HAM scaffolds. HAMs and EF-HAM scaffolds were rehydrated overnight in DPBS with 1% AA solution before being seeded with skeletal muscle cells.
Ultrastructural analysis of scaffolds
To study the effects of decellularization process on the structural integrity of HAM and to understand the physical properties of electrospun PLGA fibers in EF-HAM scaffolds with regard to fiber diameter, morphology, and alignment, sample analysis was run on Ultra-high Resolution Field Emission Scanning Electron Microscope (UHR-FESEM; Hitachi, Japan) and Phenom ProX desktop SEM (Phenom-World, The Netherlands), respectively. For analysis on native and decellularized HAMs, samples were fixed in 2.5% glutaraldehyde (Ajax Finechem, Australia) at 4°C overnight to preserve the biological structure of the specimens, washed 3 × 5 mins in DPBS to remove excess glutaraldehyde, frozen at −80°C, and freeze-dried at −56°C and 5 mTorr. Samples were sputter-coated with platinum at 30 mA for 60 s on specimen stubs before visualization under UHR-FESEM at 2 kV of acceleration voltage. Meanwhile, EF-HAM scaffolds did not undergo any pretreatment before visualization under desktop scanning electron microscope (SEM) at 10 kV of acceleration voltage.
Measurement of EF-HAM fiber diameter
Representative electron micrographs from five different positions were obtained from each sample. The mean diameter of PLGA fibers on different EF-HAM scaffolds was manually determined using Fibermetric image analysis software on desktop SEM. In total, more than 50 individual fibers have been measured for each scaffold group. For each data set, all measured diameters were combined and plotted onto a histogram. A nonlinear line was then fitted to the histogram to form a distribution curve. The same data sets were plotted on box and whisker diagrams, through which both mean and median of the fiber diameters were being determined.
Mechanical characterization of scaffolds
To determine the mechanical strength of HAM and different EF-HAM scaffolds, uniaxial tensile strength test was performed using 20 N load cell (Model UUK 5, Korea) at 0.5 mm min−1 of crosshead velocity. Before the test, scaffolds were cut into small strips of 10 mm length and 3 mm width and their thicknesses measured using digital indicator (Mitutoyo Corp., Japan). For measurement under wet condition, strips would be rehydrated with DPBS for 10 mins before the test. A minimum of six strips were tested for each scaffold group. The strips, either dry or wet, were then subjected to an axial load until broken and the force (in kgf) and elongation (in mm) were recorded using a ZR-RX25 data logger (OMRON, Australia). To calculate tensile stress, the force unit was converted to load (in N) and then divided by the cross-sectional area (in mm2) of the strip. Meanwhile, tensile strain was calculated by dividing the elongation value with the length of the strip. The ultimate tensile strength (UTS) (in MPa) and Young's modulus (in MPa) of the scaffolds were later determined from the stress/strain curve plots. UTS was determined from the highest point on the curve, which indicates the maximum stress the material can bear before breaking. Meanwhile, Young's modulus was determined by calculating the slope of the straight line on the curve, which demonstrates the stiffness of an elastic material.
Skeletal muscle tissue collection, skeletal myoblast enrichment, and culturing
Redundant skeletal muscle tissues were obtained aseptically during surgery from debridement or amputated limbs of patients with written consents. Skeletal muscle cell harvesting and culturing were performed using protocols described in our previous study with some modifications. 28 Briefly, skeletal muscle tissue was rinsed in DPBS, and any connective tissue, fat, or blood vessels present were removed from the tissue. Muscle tissue was minced and enzymatically digested with 0.25% trypsin-EDTA (TE; Sigma-Aldrich) in a 37°C shaker incubator for 10 mins to isolate muscle cells containing a mixed population of myoblasts and fibroblasts. During the incubation period, tissue fragments were gently triturated using a 10-mL pipette every 3 mins. Cell suspension was then separated from the undigested tissue through centrifugation at 500 rpm for 5 mins before being transferred into a 50-mL tube containing an equal volume of a 1:1 mixture of Nutrient Mixture F-10 Ham (F10; Sigma-Aldrich) and Dulbecco's modified Eagle's medium—high glucose (DMEM; Sigma-Aldrich) supplemented with 20% fetal bovine serum (FBS; Biowest, South America origin). These digestion steps were then repeated twice with the undigested tissue obtained from previous centrifugation. All cells in suspension were later pelleted by centrifugation at 1000 rpm for 10 mins, resuspended in F10:DMEM +20% FBS culture medium, and cultured at 37°C with atmospheric condition of 5% CO2. Waste medium was replaced with fresh culture medium every 48 h. From passage 1 onward, skeletal muscle cells were cultured in F10:DMEM culture medium supplemented with 10% FBS and grown on culture surface coated with 10 μg/mL of laminin (Sigma Aldrich) to enrich skeletal myoblast population.
Cell viability assay
The viability of skeletal muscle cells on HAM and EF-HAM scaffolds was assessed using PrestoBlue® cell viability assay (Thermo Fisher Scientific) as per the manufacturer's instructions. PrestoBlue, a resazurin-based assay, is rapidly reduced by metabolically active cells into a red and highly fluorescent by-product, which can be used to measure cell viability by either absorbance or fluorescence. Briefly, subconfluent skeletal muscle cells at passage 3 were dissociated with 0.05% TE at 37°C for 5 mins. After checking for complete cell detachment, the TE solution was immediately neutralized using equal volume of F10:DMEM +10% FBS culture medium. Cell suspension was centrifuged at 3500 rpm for 7 mins and supernatant was later removed. Cells were resuspended in fresh F10:DMEM +10% FBS culture medium and viable cell count was obtained through trypan blue exclusion assay. Skeletal muscle cells were seeded on HAM and EF-HAM scaffolds at 3000 cells/cm2 of seeding density and cultured for 7 days. At day 7 of culture, cell-seeded HAM and EF-HAM scaffolds were incubated in the dark for 24 h in 10% PrestoBlue reagent diluted with F10:DMEM +10% FBS culture medium. The cell culture supernatant from each scaffold was transferred into a 96-well plate before quantification with a spectrophotometer. The absorbance readings at 570 nm were normalized to 600 nm reference wavelength and plotted against the experimental conditions. The cell-seeded scaffolds were then fixed with 4% paraformaldehyde at 4°C overnight and were later washed in DPBS before being subjected to immunofluorescence staining for morphological observations.
Cell migration study
The dynamic cell behavior on HAM and EF-HAM scaffolds such as cell migration was monitored in real time using live imaging tool. Subconfluent skeletal muscle cells at passage 3 were incubated in the dark at 37°C and 5% CO2 with 5 μg/mL of Hoechst 33342 dye (Thermo Fisher Scientific) and 5 μM of CellTracker™ Green CMFDA dye (Thermo Fisher Scientific) for 20 min and later replaced with fresh culture medium. After 24 h, skeletal muscle cells were trypsinized and seeded on the scaffolds at 3000 cells/cm2 of seeding density and cultured for another 48 h in F10:DMEM +10% FBS culture medium before being captured by fluorescence live-cell imaging. Sample images at three predetermined positions were captured at a 10-min interval for the next 12-h and were subsequently compiled by imaging software to create time-lapse videos. For each scaffold group, a minimum of 15 cells were tracked and their migration rates evaluated using NIS-Elements D Ver4.30.00 image processing software, through which the x and y coordinates of the center of the mass of each cell were determined at each time point. The migration rate of individual cells between two time points was calculated using the following formula:
where x1 = x coordinate of cell at first time point
x2 = x coordinate of cell at second time point
y1 = y coordinate of cell at first time point
y2 = y coordinate of cell at second time point
t = time elapsed
before the average migration rate of cells on each scaffold was determined.
Immunofluorescence staining of cells on EF-HAM scaffolds
Skeletal muscle cells on EF-HAM scaffolds were previously fixed in 4% paraformaldehyde at 4°C overnight and washed in DPBS. Cells were then permeabilized with 0.5% of Triton X-100 solution (Sigma-Aldrich) for 20 mins at room temperature. After masking nonspecific binding epitopes with 10% goat serum (Gibco® Thermo Fisher Scientific, New Zealand) at 37°C for 1 h, skeletal myoblasts were incubated with the mouse monoclonal anti-human desmin antibody (diluted 1:300 in 1% goat serum; Dako Agilent Technologies) at 37°C for 2 h. After incubation with a primary antibody, cells were incubated at 37°C for 2 h with the goat anti-mouse IgG (H+L) secondary antibody (Alexa Fluor® 488; Thermo Fisher Scientific) diluted 1:300 in 1% goat serum. This was followed by counterstaining with DAPI (1:15,000 dilution) and CF™594 Phalloidin (1:70 dilution; Biotium, Inc.) for 40 mins at room temperature. Images of labeled cells were captured using fluorescence microscope and used to study cellular morphology and orientation on scaffolds.
Fiber and cell alignment
The alignment of electrospun PLGA fibers and skeletal muscle cells on HAM and EF-HAM scaffolds was determined by measuring the angle of fibers and cells relative to the horizontal axis on SEM micrographs and immunofluorescence images, respectively. For each sample, a minimum of 20 individual fibers or cells were measured for their angles using NIS-Elements D Ver4.30.00 image processing software. In total, more than 70 individual fibers and 60 individual cells have been measured for each scaffold group. Fiber and cell alignment angles were then normalized to 0° (horizontal axis) and plotted as histograms with moving average line. Data set with low angular standard deviation (SD) represents fibers and cells that were highly aligned. The positive and negative values of normalized angle indicate the opposite orientation of fibers and cells relative to the horizontal axis.
Statistical analyses
Each assay or analysis had been performed on three independent biological samples and run on triplicate unless stated otherwise in the methodology sections. All statistical analyses were conducted on GraphPad Prism 7 software. Analysis of variance with Tukey's post hoc test was used to calculate the significant difference for pairwise comparison. Mean values were reported with SD, and the significant differences were assigned when the value of p < 0.05.
Results
Decellularizing effects of TL and NaOH treatment on HAM
To remove intact epithelium from HAM, we incubated HAM with various concentrations of TL enzyme followed by a brief treatment in 0.25 M NaOH solution. Compared with untreated control (Fig. 1A), HAM treated with 12.5 μg/mL of TL (Fig. 1B) showed partial detachment of epithelial cells, whereas treating HAM with a higher concentration of TL at 25.0 μg/mL (Fig. 1D) caused smearing of fragmented DNAs across the membrane. At an intermediate TL concentration of 18.75 μg/mL (Fig. 1C), complete removal of the epithelium was achieved and the presence of DNA smear on the membrane was negligible. Ultrastructural analysis of decellularized HAM was performed using SEM to study the structural integrity of the membrane after decellularization. Decellularization using 18.75 μg/mL of TL and 0.25 M of NaOH (Fig. 1F) removed the basement membrane of HAM and exposed a fibrous and porous ECM underlying the epithelium. The ECM of decellularized HAM appeared structurally intact, and the decellularization process did not cause any major perforation to the membrane.
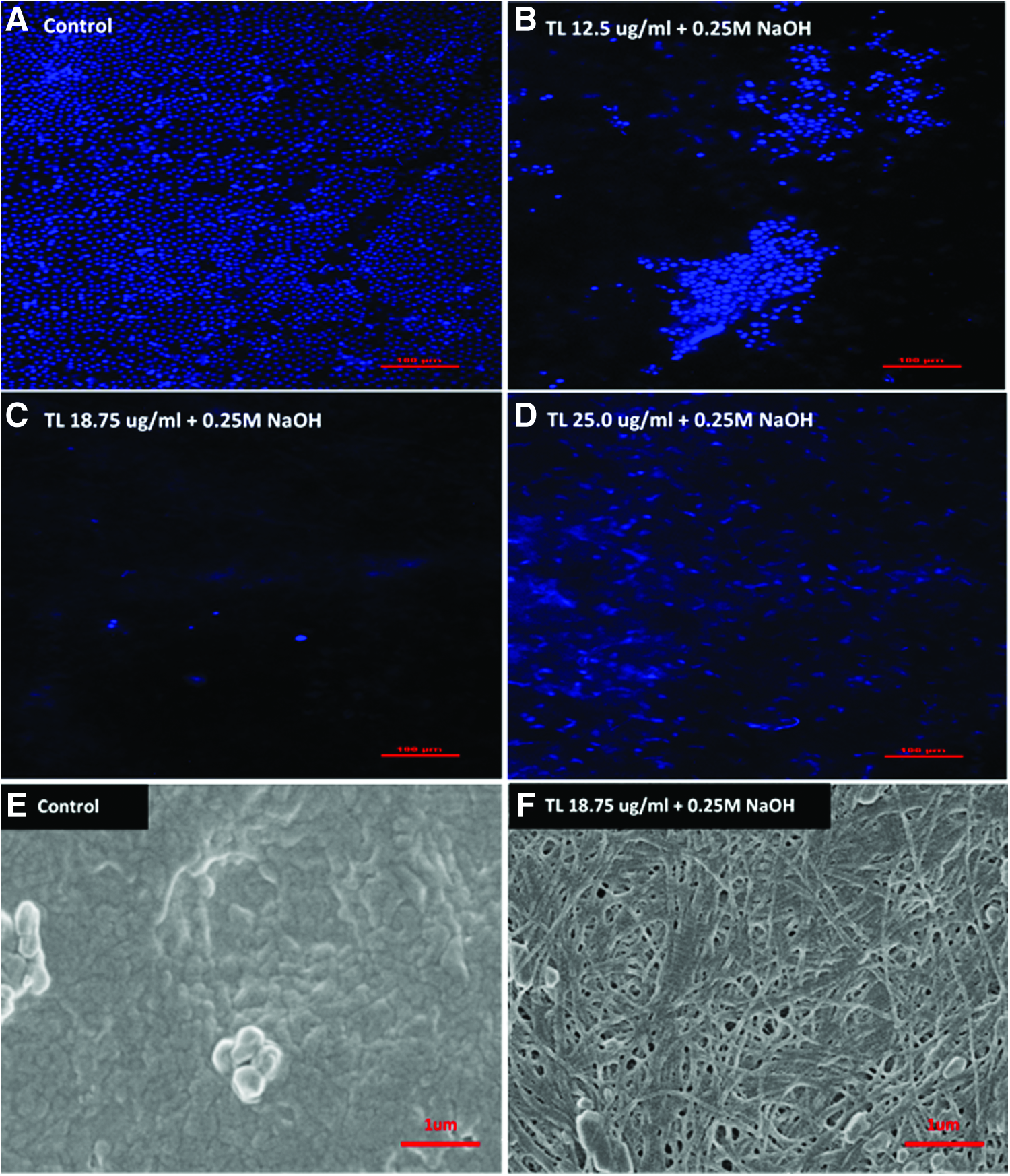
Morphological, physical, and mechanical characterization of EF-HAM scaffolds
The fabrication of 20% (w/v) PLGA 50:50 fibers on decellularized HAM, which was conducted at optimized electrospinning parameters, produced fibers of straight morphology without bead defects (Fig. 2A, C, E). The majority of fibers in EF-HAM scaffolds showed a relatively aligned arrangement as illustrated by the narrow peaks in the histograms (Fig. 2B, D, F). Nevertheless, the spinning of EFs for 3 min on HAM (Fig. 2A, B) exhibited higher alignment as evidenced by the lowest angular SD value (16.77 degrees versus 25.22 degrees for 5 min and 26.28 degrees for 7-min spinning).
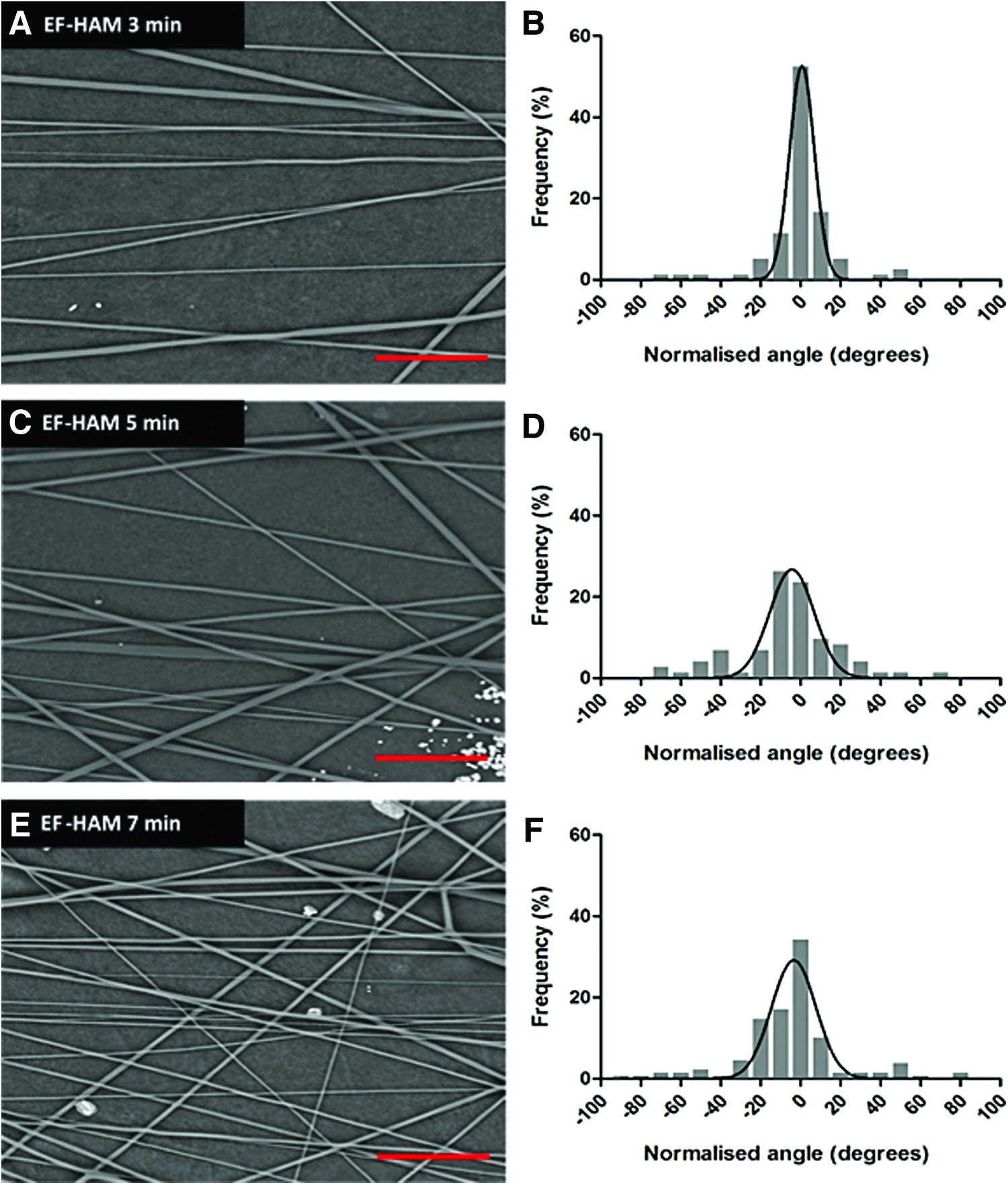
Electron micrographs and histograms showing the alignment of electrospun PLGA fibers and the normalized (to 0°) angle of PLGA fibers in EF-HAM 3-min
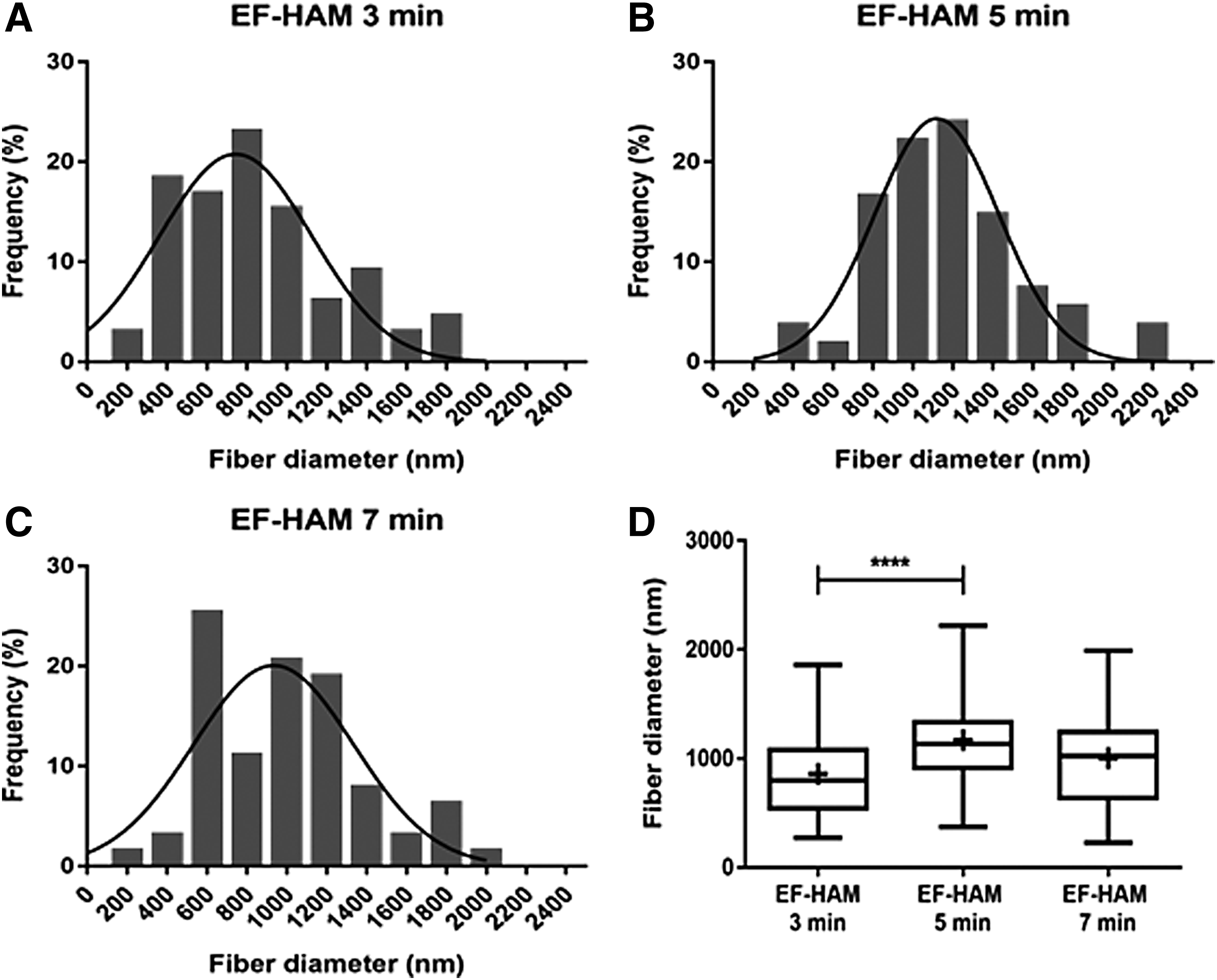
The mean diameter of electrospun PLGA fibers in EF-HAM scaffolds was determined by analyzing SEM images. As illustrated in Figure 3D, the mean diameter of electrospun PLGA fibers in EF-HAM 3-min scaffold (858.3 ± 400.3 nm) was significantly lower than that in EF-HAM 5-min scaffold (1170 ± 370.5 nm; p = 0.0001) but was comparable with EF-HAM 7-min scaffold (1005 ± 386.7 nm; p = 0.0852).
The UTS of HAM and different EF-HAM scaffolds was determined by uniaxial tensile testing. As displayed in Figure 4A, the increased deposition of PLGA fibers on decellularized HAM did not result in any significant change in the UTS of EF-HAM scaffolds when measured in dry condition. Notably, the UTS of EF-HAM scaffolds measured in hydrated condition was significantly reduced in EF-HAM scaffolds with thicker PLGA fiber coating (33.92 ± 16.4 MPa in EF-HAM 3 min vs. 21.48 ± 9.385 MPa in EF-HAM 5 min; p = 0.0353 and 17.25 ± 7.12 MPa in EF-HAM 7 min; p = 0.0043). In particular, EF-HAM 7-min scaffold was mechanically stronger in dry condition than when it was hydrated (29.96 ± 13.78 MPa vs. 17.25 ± 7.12 MPa; p = 0.0434). The Young's modulus of HAM and EF-HAM scaffolds was also evaluated to determine the stiffness of the scaffolds. Materials with high Young's modulus have higher stiffness compared with those with lower Young's modulus, which are physically more elastic. As demonstrated in Figure 4B, HAM and EF-HAM scaffolds become more elastic once hydrated (from 557.4 ± 111.8 MPa to 198.9 ± 124.1 MPa in HAM; p = 0.0019, from 799.3 ± 378.3 MPa to 267.9 ± 175.8 MPa in EF-HAM 3 min; p < 0.0001, from 604.1 ± 198.5 MPa to 202 ± 99.38 MPa in EF-HAM 5 min; p < 0.0001, and from 783.9 ± 242.5 MPa to 129.5 ± 84.1 MPa in EF-HAM 7 min; p < 0.0001). Nevertheless, the addition of PLGA fibers on HAM did not cause any significant change in the Young's modulus of EF-HAM scaffolds either in dry or wet condition, suggesting that the elasticity of HAM and EF-HAM scaffolds was mainly derived from the amniotic membrane.

Biocompatibility evaluation of EF-HAM scaffolds with skeletal muscle cells
The viability of skeletal muscle cells cultured on HAM and different EF-HAM scaffolds was evaluated using PrestoBlue cell viability assay to assess the cell growth. Our results demonstrated the viability of cells on HAM and all EF-HAM scaffolds that were similar to that of control (Fig. 5A), indicating the absence of scaffold cytotoxic effect on cells.

On the contrary, the migration rate of skeletal muscle cells on HAM and EF-HAM scaffolds was analyzed using live imaging of fluorescently labeled cells. No significant difference was observed for the migration rate of skeletal muscle cells on HAM or EF-HAM scaffolds compared with control (Fig. 5B).
Alignment of skeletal muscle cells on EF-HAM scaffolds
As shown in Figure 6, skeletal muscle cells grown on HAM alone appeared to be randomly oriented and less organized. Meanwhile, the presence of PLGA fibers on EF-HAM scaffolds provides some degree of orientation and contact guidance to the cells. Cells on EF-HAM scaffolds appeared to be more elongated and oriented along the directional axis of the fibers. Histograms in Figure 6, showing the normalized angle of cells, clearly demonstrate the distinct alignment patterns of cells on HAM and EF-HAM scaffolds. A broad range of cell angles on decellularized HAM were indicative of cells lacking a specific directional order. Meanwhile, cells grown on EF-HAM 5-min scaffold exhibited a slightly higher cell alignment compared with cells on other PLGA-coated scaffolds as revealed by the slightly lower angular SD value (13.11 degree vs. 16.28 degree in EF-HAM 3-min scaffold and 18.39 degree in EF-HAM 7-min scaffold).
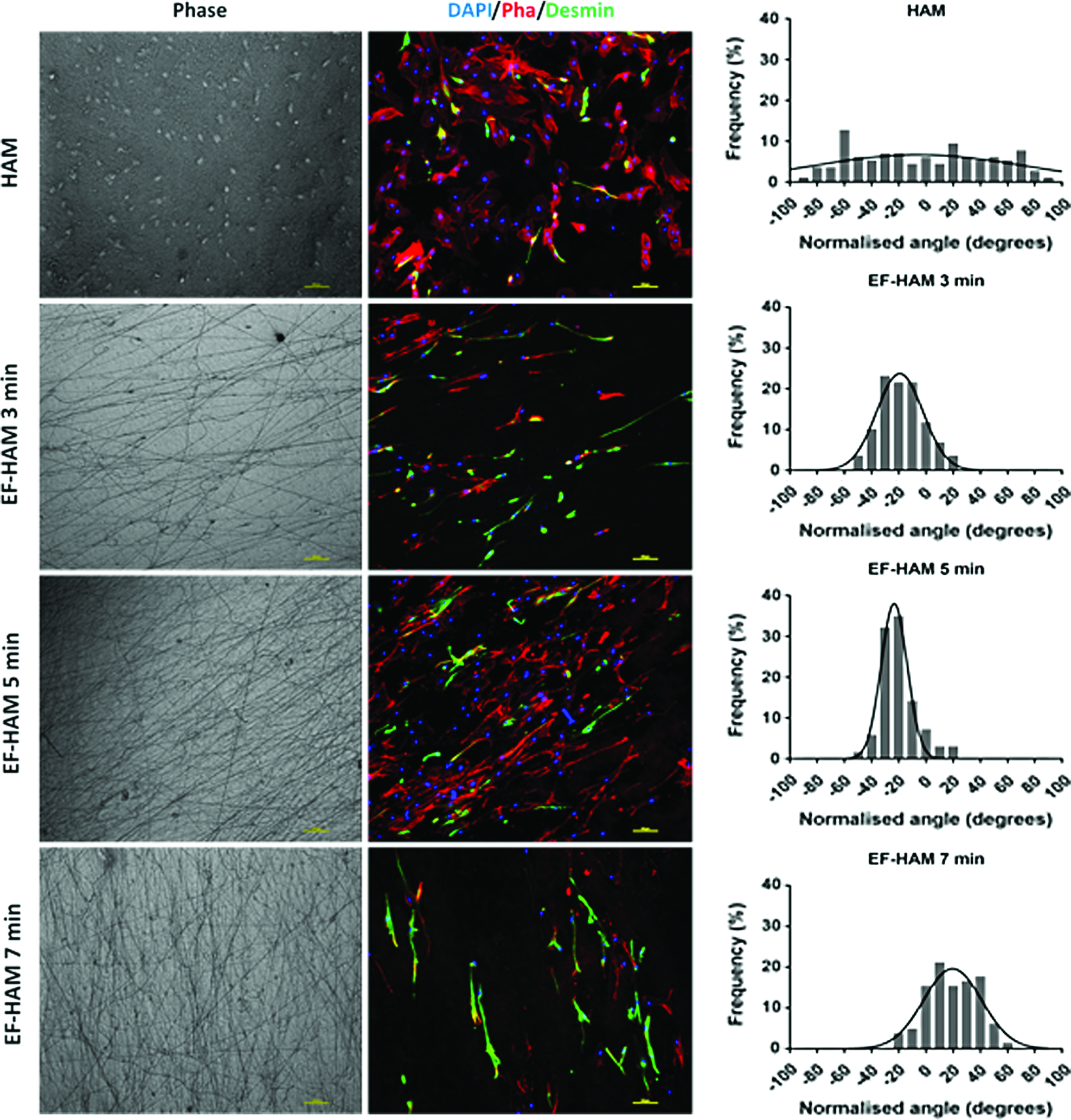
Immunofluorescence images were showing the attachment and orientation of skeletal muscle cells on HAM and EF-HAM scaffolds. Histograms were showing the normalized (to 0°) angle of cells on scaffolds. Cells grown on HAM were randomly oriented, whereas cells grown on all EF-HAM scaffolds were oriented along the alignment of the electrospun PLGA fibers. Color images available online at
Discussion
The functionality of many tissues such as in the musculoskeletal system, heart, nerve, cornea, and blood vessels depends on the cell alignment and tissue organization, which are directed by the anisotropic architecture of ECM. This unique structural feature of the matrix also contributes to the greater mechanical properties of these tissues.29–32 In recent years, different techniques such as electrospinning and self-assembly have been used to fabricate nanofiber scaffolds mimicking the aligned matrix structure, which is crucially required to facilitate cellular orientation. Extensive investigations have been performed using aligned EF to develop skeletal muscle, 33 neural, 34 bone, 35 tendon, 36 ligament, 9 and vascular 37 tissue substitutes. In this study, we have fabricated aligned PLGA fibers via the electrospinning process on decellularized HAM, which were shown to regulate skeletal muscle cell alignment on the composite scaffold, namely the EF-HAM. The use of decellularized HAM resulted in a scaffold of higher mechanical strength. Moreover, we described a simple procedure to decellularize HAM before the construction of EF-HAM scaffold.
Decellularized allogeneic and xenogeneic tissues are gaining popularity as scaffolds for tissue engineering applications as they can preserve the complex matrix architecture of target tissues or organs after the removal of residing cells. However, the choice of decellularization methods is crucial for the efficient removal of cells from the tissues; preservation of structural, biochemical, and mechanical properties of the decellularized tissues; and successful interaction of newly seeded cells with decellularized tissues consequently. In this study, we performed a simple and practical decellularization procedure on HAM using both TL and NaOH to remove HAM cellular components without causing any severe damage to the underlying ECM of the membrane. In previous studies, TL had been used at high concentrations to effectively remove the amniotic epithelium.38,39 Nevertheless, combining TL treatment with NaOH helps to lower the amount of TL being used, thus preserving the structural integrity of the membrane. Also, instead of subjecting the membrane to mechanical scraping as commonly practiced, we pulse vortexed our samples to mechanically dissociate the epithelial cells from the membrane without causing any local tears. The TL-NaOH decellularization procedure may produce DNA smearing on the membrane due to DNA fragmentation, but DNA remnants could be easily removed with the use of DNase (unpublished results). The presence of essential ECM proteins in decellularized HAM such as collagen type IV, laminin, nidogen, and fibronectin, was not being determined. However, previous studies reported that HAM treatment with either TL or NaOH did not affect the presence of these essential proteins after decellularization.38,40
The SEM images demonstrated that the protein matrix in decellularized HAM was randomly oriented. To extend the applications of decellularized HAM in aligned tissue regeneration, we fabricated a composite scaffold of HAM containing aligned EF of PLGA. Several factors have been reported to affect the alignment of electrospun nanofibers such as rotational speed, electrical fields, width of insulating gap, and applied voltage.41,42 Our results showed that increasing the deposition of fibers on decellularized HAMs would result in the decrease of fiber alignment. It is poorly understood how the increase in thickness of fiber layer could lead to the loss of fiber alignment. However, previous studies have reported similar observations and attributed them to the accumulation of residual charges in the thick fiber mat.43,44 Since the amniotic membrane is a nonconducting material, residual charges quickly accumulated as more fibers were deposited. These residual charges might cause the subsequent depositing fibers to be repelled thus altering the orientation of fibers on HAM. Polymer deposition time could also affect the diameter of EF. We found that the diameter of EF in EF-HAM significantly increased when spun from 3 to 5 mins. We postulated that a drop in the electric field gradient at the early phase of electrospinning might occur as more fibers were deposited on HAM thus reducing fiber stretching and thinning. Consequently, this would lead to an initial increase in the diameter of the EF. Electrospinning of viscous polymers has been shown previously to produce fibers with thicker diameter at low electric field, supporting our results from this study. 45
The tensile strength of our decellularized HAMs, both in dry (20.46 ± 7.477 MPa) and wet (26.34 ± 8.991 MPa) conditions, were higher than what had been reported previously.46–48 Mechanical characterization of EF-HAMs demonstrated that the increasing deposition of PLGA fibers on decellularized HAM did not significantly alter both the UTS and Young's modulus of scaffolds in dry condition. While we had not measured the tensile strength of the electrospun PLGA fibers alone, many previous studies have reported the tensile strength of not more than 13 MPa for both random and aligned PLGA fibers.7,49–51 The EF-HAM 3-min scaffold had the highest tensile strength of all, at 33.92 ± 16.4 MPa when it was hydrated, which was almost triple the strength of electrospun PLGA fibers. On the contrary, an increase in the thickness of electrospun PLGA fibers in wet EF-HAMs resulted in the reduced strength of scaffolds. Previous studies have reported the reduced tensile strength of PLGA fibers in wet condition.50,52 Due to the hydrophobic nature of PLGA polymer, the presence of thick PLGA fibers might have lowered the water absorption capacity of HAM, thus further reducing the overall strength of the scaffolds. On the contrary, HAM and EF-HAM scaffolds became more elastic when hydrated as indicated by the significant decrease in the Young's modulus of the scaffolds regardless of fiber thickness.
Topographic features such as fiber alignment, diameter, and surface roughness are known to influence cell attachment, morphology, proliferation, and differentiation.33,53,54 When skeletal muscle cells were grown on EF-HAMs, cells exhibited a higher degree of alignment than on plain and HAM surface. Surprisingly, skeletal muscle cells were most aligned on EF-HAM 5-min scaffold, even though the EF-HAM 3-min scaffold had the highest fiber alignment suggesting that cell orientation could be dictated by other topographic elements such as fiber diameter and fiber/fiber spacing.10,55 While EF-HAM scaffolds have the ability to influence cell morphology and orientation, biocompatibility analysis performed on EF-HAMs shows that no significant elevation with regard to cell viability and migration was observed compared with control or HAM alone. Thus, further studies are warranted to enhance the biocompatibility of EF-HAM scaffold, which can be achieved through several strategies, including fiber surface coating, polymer blending, incorporation of growth factors, surface functionalization with cell-adhesive peptides, and surface treatment by radiation.26,56–58
For the scope of this study, human myoblast-enriched cells have been used to study basic cellular responses toward EF-HAM morphology and topographic structure. Nevertheless, further research is warranted to establish a functional EF-HAM scaffold for use in actual muscle regeneration. Notably, the choice of candidate cells with remarkable regenerative capacity is as critical as the functional requirements of the scaffolding biomaterials. While primary myoblasts can be extensively propagated in culture and are more traditionally used for tissue engineering, satellite cell transplantation seems a more promising approach for long-term regeneration of muscle tissue.59,60 However, to date, human muscle satellite cells have not been definitively characterized and the clinical approach for the isolation and transplantation of human satellite cells is lacking. On a positive note, emerging technologies have currently focused on creating an in vitro microenvironment that could mimic the key properties of satellite cell niches so that their stem cell phenotype could be retained in culture, thus paving the way toward clinical translation of satellite cell therapy.61,62
Conclusions
In summary, besides having good mechanical strength and elasticity, EF-HAM scaffold also possesses uniaxially aligned fibers, which could facilitate guided orientation and migration of skeletal muscle cells. We also demonstrated that EF thickness does have an influence on fiber alignment, diameter, and tensile properties. On further improvement on the biocompatibility of EF-HAM scaffolds, we could prospectively explore a wider range of applications where EF-HAM could be potentially used for the reconstruction of any bodily tissue with a highly organized structure.
Footnotes
Acknowledgments
This work was supported by the Ministry of Science, Technology and Innovation, Malaysia (Science Fund: 02-01-02-SF1284) and Ministry of Higher Education, Malaysia (Fundamental Research Grant Scheme: FRGS/2/2013/SG05/UKM/03/1). We also thank Mrs. Nor Kamalia binti Zahari and Mr. Mohd Asyraf bin Mat Afandi for helpful discussions and guidance, and Mr. Cosmos George and Dr. Ruzalina binti Baharin from the Malaysian Nuclear Agency for providing assistance for gamma sterilization.
Disclosure Statement
No competing financial interests exist.