
Abstract
The bone marrow microenvironment is the preferred location of multiple myeloma, supporting tumor growth and development. It is composed of a collection of interacting subniches, including the endosteal and perivascular niche. Current in vitro models mimic either of these subniches. By developing a model combining both niches, this study aims to further enhance the ability to culture primary myeloma cells in vitro. Also, the dependency of myeloma cells on each niche was studied. A 3D bone marrow model containing two subniches was created using 3D bioprinting technology. We used a bioprintable pasty calcium phosphate cement (CPC) scaffold with seeded osteogenic multipotent mesenchymal stromal cells (O-MSCs) to model the endosteal niche, and Matrigel containing both endothelial progenitor cells (EPCs) and MSCs to model the perivascular niche. Within the model containing one or both of the niches, primary CD138+ myeloma cells were cultured and analyzed for both survival and proliferation. The 3D bone marrow model with combined subniches significantly increasing the proliferation of CD138+ myeloma cells compared to both environments separately. The developed model showed an essential role of the perivascular niche over the endosteal niche in supporting myeloma cells. The developed model can be used to study the expansion of primary myeloma cells and their interactions with varying bone marrow subniches.
Introduction
B
Multiple myeloma is a B cell malignancy characterized by the clonal proliferation of malignant plasma cells within the bone marrow. 8 Bone marrow niches are utilized by invading myeloma cells that depend on this environment. 9 The prolonged presence of myeloma cells in the bone marrow leads to permanent changes in the residing MSCs that are associated with bone lesions and defect bone repair.8,10
For myeloma, it is not yet clear if there is a preferential outgrowth in either of the bone marrow subniches, and which microenvironment or interactions are most decisive for the survival of myeloma and the accompanying bone destruction. Also, to enable prolonged in vitro culture of primary myeloma cells, it should be analyzed whether the combined presence of both environments would be beneficiary. To study this, a reliable culture model mimicking these separate but interacting niches is needed. The establishment of such a model aids the understanding of complex physiological and pathophysiological processes.11,12 Even though in vitro 3D models remain experimental tools that do not reproduce the entire in vivo environment, they do span the gap between the 2D culture of cells and in vivo animal models. 13
For primary myeloma cell culture, existing models of the endosteal niche use autologous bone fragments, 14 mineralized silk scaffolds, 15 or ossified polycarbonate membrane disks. 16 Models reconstituting the perivascular niche have mimicked this microenvironment using bone marrow mononuclear cells (MNCs), MSCs, or combinations of MSCs and endothelial cells in soft extracellular matrix (ECM)-like materials.17–20 Thus far, no models have been described that combine both niches to investigate their joint effect on the in vitro survival of primary myeloma cells. Therefore, we set out to further progress these models by combining a perivascular environment with a mineralized matrix representing the endosteal niche, to study interactions between primary myeloma cells and either of these subniche microenvironments.
To create a complex porous 3D in vitro model with defined regions, additive manufacturing techniques can be used. These techniques offer the possibility to precisely control the shape and composition of a manufactured construct. 21 One of these techniques, 3D bioprinting, enables the building of 3D constructs, in which the design of the structure and the placement of components are predetermined. Such a construct is created by extruding a desired material in a layer-by-layer manner, thus, resulting in porous microtissues. This technique allows inclusion of different biomaterials at ambient temperatures, providing the possibility to incorporate cells or growth factors. 22
Commonly used biomaterials for bone mimicking scaffolds include metals, polymers with either natural or synthetic origins, and ceramics. 23 Ceramics are among the most promising biomaterials for bone tissue engineering, either used alone or as a composite combined with polymeric materials.24,25 Ceramics are both biocompatible and bioactive, because of their ability to form a hydroxy carbonate apatite layer that is chemically and structurally equivalent to the mineral phase in bone. 26 Most ceramics are processed into scaffolds at high temperatures, reducing the possibility to incorporate biological components such as growth factors. This is why recent studies have focused on ceramic materials such as pasty calcium phosphate cement (CPC), which can be processed at ambient temperatures using 3D bioprinting.27,28
The aims of the current study were (1) to test the feasibility of fabricating a model containing two separate but interacting niches, (2) to develop an in vitro 3D bone marrow myeloma culture model containing both perivascular and endosteal niches, and (3) to investigate the added value of separate osteogenic and vascular niches to the in vitro culture of primary myeloma cells. Design parameters included cytotoxicity of the CPC scaffolds for endothelial progenitor cells (EPCs) or myeloma cells, and the most optimal external shape and size of the CPC scaffolds for imaging and handling, biofunctionalization, and the ability to integrate the perivascular coculture. The stability of the combined endosteal and perivascular coculture was analyzed by tracking migration of cells from one compartment to the other. Most importantly, the 3D model containing two niches was validated by examining its potential to propagate primary CD138+ myeloma cells cultured within the model over 28 days.
Materials and Methods
Cell lines and primary cells
Human myeloma cell lines OPM2 and L363 were cultured in myeloma medium: advanced RPMI 1640 media supplemented with 10% (v/v) fetal bovine serum (FBS), 2 mM of L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (all Gibco).
Human bone marrow was aspirated from the acetabulum of three patients undergoing hip replacement surgery. MSCs were isolated from the bone marrow by adherence to tissue culture plastic and expanded in MSC expansion medium: α-minimal essential media (αMEM; Gibco) supplemented with 10% (v/v) FBS, 0.2 mM L-ascorbic acid 2-phosphate, 100 U/mL penicillin, and 100 μg/mL streptomycin (all Gibco). MSCs were used at passage 2–4.
Human bone marrow was obtained from the spina iliaca posterior superior of four myeloma patients. The CD138+ cell population was isolated from the MNC fraction of the myeloma bone marrow by microbeads (Miltenyi Biotec) and used immediately in the coculture system. The CD138+ cell fraction was cultured in advanced RPMI 1640 media (Gibco) supplemented as detailed above.
Human umbilical cord blood was collected from two full-term pregnancies. MNCs were isolated from this cord blood by density-gradient centrifugation using Ficoll-Paque, and seeded on collagen I (BD Biosciences)-coated wells. Colonies were expanded in EPC expansion medium: EGM-2 media (Lonza), SingleQuots™ Kit (Lonza), 10% (v/v) FBS (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco). The isolated cells were previously characterized as late outgrowth EPCs. 29 EPCs were used at passage 2–5.
All primary samples were obtained after written informed consent, and protocols were approved by the local Ethics Committee of the University Medical Center Utrecht (NL).
CPC scaffold fabrication
Using the BioScaffolder 3.1 system (GeSiM), constructs of pasty calcium phosphate cement (CPC; Velox, InnoTERE) were produced by 3D fiber deposition. The constructs were printed using a needle having an inner diameter of 230 μm (Globaco). Printer settings can be summarized in short as follows: (1) a porous disk was printed with a diameter of 8 mm and a height of 0.48 mm; (2) a porous cylindrical construct was printed with a diameter of 5 mm and a height of 4 mm; (3) a porous hexagonal construct was printed with sides of 3 mm, a diameter of 6 mm and a height of 4 mm; and (4) a porous hexagonal construct was printed with sides of 3 mm, a diameter of 6 mm and a height of 0.72 mm. A pressure of 2–2.5 bars was used with a strand distance of 0.6 mm. The cement-setting reaction of the CPC was started in a humidified chamber and lasted 3 days. Next, the scaffolds were incubated for 1 h in acetone to remove excess oil, washed in distilled water, dried in air at 37°C, and stored at room temperature until use. The final pore size of the endosteal scaffold after setting was 330 ± 25 μm, lying within the pore size distribution measured in decellularized cancellous bone chips (440 ± 150 μm, Bislife). Scaffold pictures were taken with a Olympus SZ61 Stereo Zoom Microscope. Scaffolds used for cell culture experiments were sterilized by gamma irradiation (25 kGy).
Biochemical analysis of cell proliferation
Both EPC donors and two myeloma cell lines were cultured on top of the CPC disks for 14 days (Fig. 1). On each scaffold, 105 cells were seeded. The number of cells grown on the scaffolds (EPCs) or present in the well (myeloma cells or control EPCs) was determined after 1, 7, and 14 days of cultivation by measurement of the intracellular lactate dehydrogenase (LDH) activity as described earlier. 30 In short, the frozen samples were thawed and lysed by 1% Triton X-100 in PBS on ice, supported by sonication. Lysates were used for measurement of LDH activity with the CytoTox96 Non-Radioactive Cytotoxicity Assay (Promega) and correlated with the cell number using a calibration line.

Methods overview of the constructed compartments and final two-compartment model. MM, multiple myeloma; MSCs, multipotent mesenchymal stromal cells; EPCs, endothelial progenitor cells; CPC, calcium phosphate cement. Color images available online at
Cell culture on CPC scaffolds
MSCs of three donors were added on top of the cylindrical CPC scaffolds (Fig. 1) and incubated statically or dynamically on a plate shaker for 6 h. MSCs were dynamically seeded on top of the high and low hexagonal scaffolds for 6 h. On each scaffold, 105 MSCs were seeded. The seeded scaffolds were cultured in osteogenic medium (MSC-medium plus 10 nM dexamethasone (Sigma) and 10 mM β-glycerophosphate (Sigma). Cell viability and distribution were analyzed at days 1, 7, 10, and 14 using the Live/Dead Viability/Cytotoxicity Kit for mammalian cells according to the manufacturer's protocol (ThermoFisher).
Two compartment 3D culture model
MSCs labeled with lipophilic Cell-Tracker dye DiI (Invitrogen) were added on top of the cylindrical CPC scaffolds and incubated dynamically on a plate shaker for 6 h. The seeded scaffolds were cultured in either MSC expansion medium or osteogenic medium for 14 days before coculture. MSCs and EPCs, in a 4:1 ratio (2.0 × 106 MSCs/mL, 0.5 × 106 EPCs/mL), were mixed into 50% (v/v) growth factor-reduced Matrigel (Corning, diluted by an equal volume of MSC-medium). The MSCs were labeled with lipophilic Cell-Tracker dyes DiO and the EPCs with DiD (Invitrogen). Each MSC or osteogenic MSC (O-MSCs) seeded cylindrical CPC scaffold received 30 μL of Matrigel cell suspension (Fig. 1). The constructs were statically cultured in mix medium: MSC expansion medium and EPC expansion medium in a 1:1 ratio. Medium was refreshed twice a week. The stability and integration of the two compartments were imaged on day 2, 7, 10, and 14. Before imaging, the samples were fixed for 30 min in formalin; the nucleus of the cells was stained with Hoechst 33342 (Invitrogen). The scaffolds were cut in half before imaging for a cross-sectional view.
Culture of primary myeloma cells in the two compartment 3D model
MACS selected CD138+ myeloma cells of four donors were labeled with DiD (Invitrogen) and mixed with MSCs and EPCs (both DiO labeled; Invitrogen) in a 4:1:1 ratio. Fifty percent (v/v) Matrigel was added to the cells and the cell suspension was either cultured alone or added to the hexagonal scaffolds. The scaffolds were seeded with either MSCs or O-MSCs, both cell types labeled with DiI (Invitrogen) (Fig. 1). The four different culture conditions are detailed in Table 1. The constructs were statically cultured in myeloma mix medium: MSC expansion medium, EPC expansion medium, and myeloma medium in a 1:1:1 ratio. Medium was refreshed twice a week. The cells cocultured in the two compartment model were live imaged on day 1, 7, 14, 21, and 28. Cells were tracked for compartment interactions, cell migration, and myeloma outgrowth.
Condition one contains the perivascular coculture in Matrigel. Condition two combines the perivascular coculture in Matrigel with the CPC scaffold seeded with undifferentiated MSCs. Condition three combines CD138+ cells only in Matrigel with the CPC scaffold seeded with O-MSCs. Condition four combines the perivascular coculture in Matrigel with the CPC scaffold seeded with O-MSCs.
MSC, multipotent mesenchymal stromal cell; EPC, endothelial progenitor cell; CPC, calcium phosphate cement; O-MSC, osteogenic differentiated MSCs.
The overall cell viability of the two compartment model was analyzed during 28 days using the Live/Dead Viability/Cytotoxicity Kit for mammalian cells according to the manufacturer's protocol (ThermoFisher). This staining was performed on scaffolds seeded with non-DiO, DiI, or DiD labeled cells.
Osteocalcin immunocytochemistry
MSCs were cultured on top of the hexagonal scaffolds in osteogenic or expansion medium for 28 days. At 7, 14, 21, or 28 days, scaffolds were fixated for 30 min in formalin. The scaffolds were incubated for 10 min in 0.2% (v/v) Triton X-100 in PBS, and thereafter blocked for 30 min with 5% (v/v) bovine serum albumin/PBS. The samples were incubated overnight at 4°C with 10 μg/mL mouse monoclonal antibody recognizing human osteocalcin (clone OCG4; Enzo Life Sciences). This was followed by incubation with 10 μg/mL goat-anti-mouse polyclonal antibody conjugated to Alexa Fluor 488 (Invitrogen). All samples were also stained for F-actin (TRITC) and DAPI (both FAK100 Kit; Merck Millipore), according to the manufacturer's protocol.
Confocal imaging
The stability and migration of the combined perivascular and endosteal model were imaged using a Leica TCS SP5 confocal microscope. PMT detectors collected fluorescence signal from Hoechst (405/460–480), DiO (484/500–540), DiI (549/565–605), and DiD (644/665–705), respectively, represented in the images by pseudocolors blue, green, yellow, and cyan.
All other fluorescence images were taken with a Leica SP8X Laser Scanning Confocal Microscope using a white light laser (470–670 nm) and Leica LASX acquisition software. The cell distribution of MSCs seeded on CPC scaffolds was live imaged: hybrid detectors collected fluorescence signal from calcein (494/500–525) or ethidium homodimer-1 (528/600–640), which were given the pseudocolors green and red.
The primary myeloma cells cultured in the two compartment model were live imaged: hybrid detectors collected fluorescence signal from DiO (484/500–540), DiI (549/565–605), and DiD (644/665–705), which were given the pseudocolors green, yellow, and red.
The osteocalcin staining was imaged using hybrid detectors collecting fluorescence signal from DAPI (405/430–480), Alexa Fluor 488 (488/490–525), and TRITC (532/540–575), which were given the pseudocolors blue, green, and red.
All z-stack images were processed using ImageJ 1.51 h software to create single maximum projections. Images of large scaffolds used the mosaic function of the Leica LASX software, stitching the images together using smooth and linear blending.
Statistical analyses
All experimental groups were handled in technical triplicates. Results are presented as mean ± standard deviation for the indicated number of donors. p values are based on Student's t tests (2-tailed) for two-way comparisons or a two-way repeated measurements ANOVA for multiple hypothesis testing, using post hoc Bonferroni correction. Statistical analyses were performed using IBM SPSS 20 (IBM SPSS, Inc.), differences being statistically significant when p < 0.05.
Results
Viability of EPCs and myeloma cells maintained when cultured with a CPC scaffold
For MSCs, it is known that their viability is not affected when cultured on top of a CPC scaffold.28,30 In the previous studies, CPC scaffolds were not used to culture primary endothelial cells or myeloma cells. In this study, porous CPC disks (Fig. 2B) were cultured together with EPCs or myeloma cell lines, and cell expansion was analyzed over time. With or without CPC disks, the number of viable EPCs showed no significant differences on day 1, 7, or 14. Also, whether cultured with or without CPC disks, the number of viable myeloma cells was unaffected on day 1, 7, or 14 (Fig. 2A).
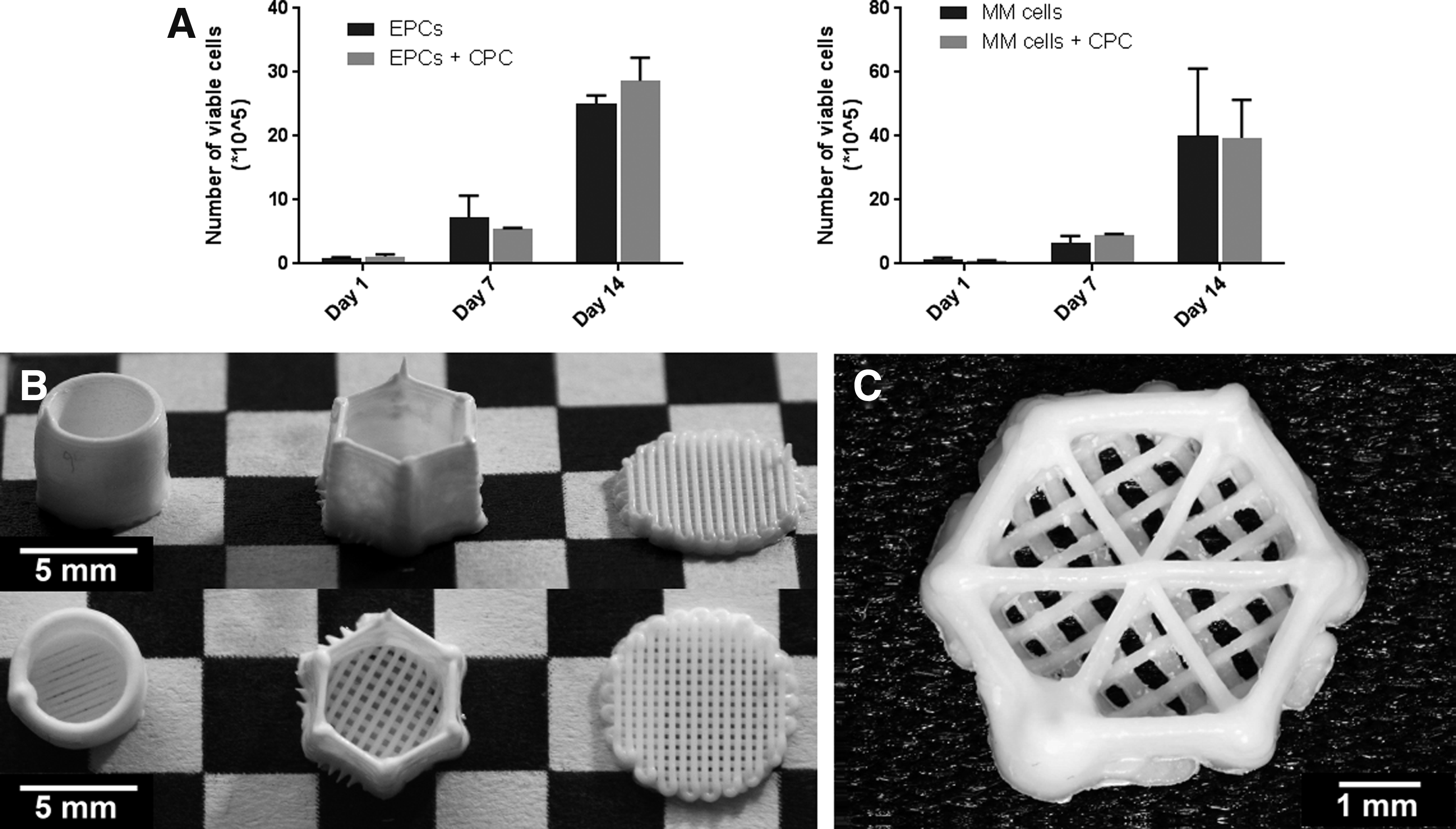
Plotted CPC scaffolds and their cytotoxicity.
MSC distribution and outgrowth dependent on seeding, culture, and scaffold design
The perivascular compartment was modeled using 50% (v/v) Matrigel, MSCs and EPCs, based on previous research. 20 To create a two-compartment bone marrow model, the endosteal compartment needed to be designed and optimized. A cylindrical, cup-like scaffold was designed for the endosteal CPC scaffold, to which the perivascular component was added (Fig. 2B). By incorporating the perivascular model within the endosteal model, the confined space ensured enough gelling time to give it the desired shape.
To biofunctionalize the cylindrical CPC scaffold, MSCs were seeded on top/inside the CPC scaffold. The bottom of each scaffold could be imaged, but the curved sides of the cylindrical scaffolds hindered imaging by confocal microscopy, due to depth limitations in the z-direction (maximum of 1 mm). This technical problem was addressed by switching to a hexagonal design (Fig. 2B), with six straight sides suitable for confocal imaging.
A confluent layer of O-MSCs was found in the bottom of both the static and dynamically seeded constructs after 7 days when cultured in osteogenic medium (Fig. 3B). This layer also covered the pores of the construct (Fig. 3C, D). To maximize contact between the endosteal and perivascular compartments, MSC adherence to the sides of the scaffold was also optimized. Static seeding of MSCs in the hexagonal scaffolds did not result in MSC attachment to the sides. Some adherence of MSCs to the sides of the scaffolds resulted from dynamic seeding of MSCs. However, varying cell densities were observed in the upper and lower part of the scaffold during 14 days of culture (Fig. 3A, E). No reproducible results could be obtained using the 4 mm high scaffolds.

Seeding efficiency and O-MSC viability on 4 mm height CPC scaffolds after dynamic cell seeding.
A third design decreased the height of the scaffold from 4 to 0.72 mm (Fig. 2C). The lower outside border still enabled the addition of the Matrigel in its liquid phase, confining it until gelation. As previous scaffold designs only facilitated contact between the endosteal and perivascular compartments at the bottom and outside border of the construct, three CPC strands were added in the middle of the construct to increase the contact area between the endosteal and perivascular compartments. This construct could be imaged entirely using confocal imaging, in the x, y, and z direction. A reproducible, confluent layer of O-MSCs was obtained 7 days after dynamic seeding and remained present at later time points (Fig. 4A–G).

Seeding efficiency and MSC viability on 0.7 mm height CPC scaffolds.
Combining perivascular and endosteal compartments
The bone marrow model containing two separate but interacting compartments was evaluated for (1) stability over time, (2) reproducibility, and (3) effective confocal imaging. Its perivascular component could be incorporated after fabrication of the endosteal CPC/O-MSC scaffold. A homogeneous incorporation of the perivascular culture into the endosteal scaffold was obtained in all constructs (Fig. 5A), and prevascular MSC/EPC structures formed in between the CPC/O-MSC strands (Fig. 5B). The Matrigel coculture remained located within the CPC/O-MSC scaffold during 28 days, but decreased in height over time. The integration of the two compartments at microscopic level could be tracked over time using confocal imaging.

Combining the perivascular and endosteal compartments. Confocal images, day 10, showing
Interactions and migration in the two compartment bone marrow model
After integrating both environments into one model, the cultures were fixed and analyzed at different time points, imaging cell interactions and migration. At day 7, a stable layer of O-MSCs could be found covering the CPC strands (Fig. 6A-I, B-I). Some cells from the perivascular culture (both MSCs and EPCs) migrated toward this confluent O-MSC layer, indicating interactions between the two compartments. No O-MSCs were found in the perivascular compartment. At day 10, prevascular networks of MSCs and EPCs were formed in the vicinity of the O-MSC layer covering the CPC (Fig. 6A-II, B-II). At day 14, networks of combined MSCs, EPCs, and O-MSCs could be found in the region adjacent to the CPC strand and O-MSC layer (Fig. 6A-III, B-III). The region most distant from the O-MSC layer consisted only of MSCs and EPCs networks (Fig. 6A-III), capable of forming luminal structures. 31

Formation of interactions between the perivascular and endosteal compartments. Confocal images showing the side view of the CPC scaffolds (in blue, nonspecific Hoechst staining), with an added Matrigel coculture. The CPC scaffolds were seeded with O-MSCs (DiI, yellow) before adding the Matrigel containing both MSCs (DiO, green) and EPCs (DiD, cyan). Nuclei in blue (Hoechst).
Over time, the model maintained varying environments, starting with an O-MSC layer bound to the CPC, flowing into a mixed region with O-MSCs, MSCs, and EPCs, then flowing into a region with only MSC and EPC networks. Interactions and migration of the different cell types were observed and quantified assessing the presence of the each cell type in either the perivascular or endosteal compartment (Fig. 6C).
The two-compartment bone marrow model enhances primary myeloma cell propagation
To investigate the additive effect of the two combined environments on the in vitro expansion of myeloma cells, primary CD138+ myeloma cells (n = 4) were cultured within the developed two-compartment bone marrow model. This model was, in different forms (Table 1), compared to a single perivascular coculture or single endosteal coculture. The perivascular coculture was added to either the CPC/O-MSC scaffold mimicking the endosteal compartment or to a nondifferentiated CPC/MSC scaffold.
The two-compartment model had a sustained viability during the 28 day culture. The endosteal scaffold had a sustained osteogenic differentiation during the 28 day culture (Fig. 7). The CPC/O-MSC scaffold significantly increased the number of CD138+ myeloma cells within the culture at day 14, 21, and 28 compared to the perivascular environment alone (Fig. 8A). Outgrowths of myeloma cells were observed both in the perivascular coculture, as on top of the CPC/O-MSC scaffold (Fig. 8D-IV). The myeloma cells were seeded in the perivascular coculture at day 0. Over time, the average relative contribution of myeloma cells in the endosteal compartment increased from 6.8% at day 7 to 40.8% on day 28 (Fig. 8B). The combined perivascular and CPC/MSC scaffold showed similar growth of the CD138+ myeloma cells compared to the perivascular coculture alone. No outgrowth of myeloma cells was observed on top of the CPC/MSC scaffold (Fig. 8D-III). Thus, osteogenically differentiated MSCs seem to provide signals that complement the vascular niche in stimulating myeloma cell expansion. Interestingly, the endosteal compartment alone (CPC/O-MSC scaffold, Fig. 8D-II) was not capable of supporting the survival of the CD138+ myeloma cells, in contrast to the single perivascular coculture (Fig. 8D-I). Thus, the MSCs and EPCs in the perivascular environment seem to be an essential element for primary CD138+ myeloma cell survival, further enhanced by adding an osteogenic compartment.

Osteogenic differentiation and viability of the model followed over time.

Culture of primary CD138+ myeloma cells in the two-compartment model.
Discussion
Using 3D bioprinting, we developed a reproducible bone marrow model that mimics both the endosteal and perivascular niche. This two-compartment bone marrow model provides a novel tool to study the biology and interactions of primary myeloma cells within varying microenvironments. Primary myeloma cells are known to depend on bone marrow niches for their survival and progression,32,33 with the precise processes and interactions involved still being investigated. By analyzing the outgrowth of primary myeloma cells within these separate or combined environments, their dependency on each other can be studied. The developed model has shown an essential role of the perivascular niche over the endosteal niche in supporting myeloma cells, with the best support of primary myeloma cells when combining both environments.
To fabricate the perivascular niche, Matrigel was used as a bioactive 3D environment, encapsulating MSCs and EPCs. 20 Not only Matrigel but also collagen, PuraMatrix, and fibrin have been used before to support 3D bone marrow models, for the in vitro culture of myeloma cells.17–19,34 To fabricate the endosteal environment, a ready to use, printable CPC paste was used. 28 Calcium phosphates have a composition similar to bone mineral, with chemical properties suitable for bone remodeling.35,36 CPCs have also been used for their low-temperature setting reaction and porosity that allows for the incorporation of drugs or growth factors.30,37,38 In addition to a growth factor or drug delivery system, CPCs can also be used as a base for an in vitro bone or endosteal model. Previous in vitro bone models have used different calcium phosphate ceramics, allowing cell attachment, perfusion, and mechanical loading of the cultures.39,40 However, these ceramics do not allow growth factor addition, nor can be printed into every shape desired, in contrast to the CPC paste used for our study.
In previous studies, MSCs cultured on CPC scaffolds showed good biocompatibility. 28 In this study, the viability of the EPCs and myeloma cells cultured together with the CPC scaffolds was also shown to be unaffected, which was expected since CPCs are known to be highly biocompatible. 31 However, in addition to biocompatibility, the model requires that cells have the ability to attach and proliferate on the scaffold material. 41 In our study, the formation of a confluent MSC layer on top of the CPC scaffolds was affected by the architecture of the scaffold, the seeding, and the culture method. We were able to obtain a stable distribution and outgrowth of MSCs on 0.72 mm high hexagonal CPC scaffolds, dynamically seeded and cultured in osteogenic medium. This osteogenically differentiated MSC layer also forms the biological component of the endosteal niche in the developed model. Apart from a limitation in the height of the scaffolds, different shapes and sizes are feasible for the model. This provides the possibility to up or down scale the model for future applications.
After the fabrication of an endosteal scaffold, a previously developed perivascular coculture was added, known to support primary myeloma cells in vitro. 20 After combining both envi ronments, a stable layer of O-MSCs remained covering the CPC scaffold at all time points. Cell migration from one compartment to the other was observed, through the surrounding Matrigel matrix, known to facilitate migration. 42 Over time, a gradient developed in the model, creating a heterogeneous in vitro model, which is in general difficult to produce. 43 However, this gradient developed spontaneously and could potentially differ from one culture to the other. By using multichannel bioprinting, different cell types could be deposited in varying but controlled ratios per layer, creating a controlled gradient. Such a model would further enable the study of the varying bone marrow environments, as it is not yet clear whether endosteal and perivascular microenvironments function separately or are one interacting entity. 44
Once the two compartment model was developed, the added biological value of such a model became apparent. The two-compartment bone marrow model enhances primary myeloma cell propagation, compared to the perivascular environment alone as well as the perivascular environment combined with a nonosteogenic MSC/CPC scaffold. Interestingly, when using the endosteal O-MSC/CPC scaffold, no myeloma cell survival was observed. Previous models using only an osteogenic compartment did see primary myeloma survival.15,16,45 In these, primary myeloma cells were shown to be viable after 11 days 15 or 21 days, 45 and decreased viability was shown after 28 days. 16 The endosteal system maintaining primary myeloma viability over the course of 21 or 28 days used a perfused environment, as opposed to our static system. It has also been shown that in vivo, the endosteal region has a higher oxygen tension compared with the perivascular region. 46 In this light, it remains to be seen whether different results can be obtained using our endosteal environment in a perfused system. Second, it has been suggested that quiescent myeloma cells reside in the endosteal niche, where they are protected from apoptotic stimuli. 5 The quiescent nature of the supported myeloma cells within the endosteal niche could also explain the lack of outgrowth observed in our system.
In general, engineered 3D in vitro models provide a controllable environment for the incorporation of specific cell types, ECM molecules, growth factors, and other biochemical cues that can potentially better simulate a native microenvironment. 47 However, challenges and limitations will remain associated with in vitro models. Also, the developed, more complex, two-compartment model does not fully represent the native myeloma microenvironment, for instance, lacking natural occurring hematopoiesis and immune system interactions.
Conclusions
The developed 3D two-compartment bone marrow model provides a novel tool to study the biology and interactions of primary myeloma cells within varying microenvironments. By using 3D bioprinting, a reproducible model was created, which recapitulates important characteristics of the native tumor microenvironment. The mere outgrowth of primary myeloma cells, which does not occur in a 2D setting or by providing feeder cells, indicates that the most essential bone marrow-derived signals are present. This in vitro BM model, mimicking both the endosteal and perivascular niche, is particularly interesting to study myeloma–bone or myeloma–MSC interactions that lead to increased bone resorption and the development of osteolytic lesions. By studying and better understanding these interactions, new treatments can be developed aiming to reverse myeloma inflicted changes in the surrounding bone marrow microenvironment. The BM model offers the possibility to analyze chemotherapeutic drug response and other tumor characteristics and could be further developed toward a drug-screening tool.
Footnotes
Acknowledgments
This study was supported, in part, by a University Utrecht Seed Grant (2013) and the Dutch Arthritis Foundation.
Disclosure Statement
No competing financial interests exist.