
Abstract
Mesenchymal stem cells/marrow stromal cells (MSCs) are attractive for applications ranging from research and development to use in clinical therapeutics. However, the most commonly studied MSCs, adult bone marrow MSCs (A-MSCs), are limited by significant donor variation resulting in inconsistent expansion rates and multilineage differentiation capabilities. We have recently obtained permission to isolate pediatric MSCs (P-MSCs) from surplus iliac crest bone chips. Here, we developed a simple and easily replicable isolation protocol yielding P-MSCs, which adhere to MSC defining guidelines. After confirming immunophenotypic marker expression, we compared expansion rates, senescence, morphology, and trilineage differentiation of P-MSCs to A-MSCs for multiple donors. We found P-MSCs have faster in vitro replication, consistently show significantly lower senescence, and are capable of more reproducible multilineage differentiation than A-MSCs. We, therefore, believe P-MSCs are a promising candidate for use in research applications and potentially as part of an allogeneic therapeutic treatment.
Introduction
O
In order for MSCs to be used as part of AMTPs, a sufficient quantity of cells must be obtained that are capable of producing consistent outcomes that satisfy the regulatory requirements set by the European Medicines Agency (EMA) and the Food and Drug Administration (FDA).17–19 When removed from the environment of their in vivo niche and expanded in vitro, MSCs rapidly lose their ability to replicate and differentiate, meaning their characteristics change unpredictably over time in culture.16,20–22 This variation is often observed in bone marrow (BM) MSC populations, which as of now is still considered the gold standard when it comes to MSCs.23,24 Other MSC sources such as umbilical cord and adipose tissue are being actively characterized with promising but conflicting results.11,25–27 An ideal MSC source would allow isolation with minimal patient discomfort and yield cells capable of reproducibly meeting EMA/FDA regulatory requirements. 28 Although BM MSCs are currently the gold standard for MSCs, their isolation is associated with a painful procedure and harvesting of such material results in substantial patient discomfort and recovery time when used.27,29 Many researchers, us among them, use surplus clinical material obtained from patients undergoing surgical procedures (total hip or knee replacement, for example). However, the age and disease status of the donors often negatively influence MSC performance. 20 Kretlow et al. found that cell attachment, proliferation, and differentiation were all affected as donor age increased.30,31 MSCs from aged donors were less capable of secreting and maintaining a chondrogenic matrix, 32 and had a decreased bone-forming potential in vivo. 33 It has also been reported that cells from the elderly often exhibit cellular dysregulation, which negatively impacts stem cell populations.31,34,35 In addition, MSCs from elderly patients have been shown to have age-induced gene expression changes and earlier replicative senescence that further negatively effects MSC performance.31,36 Cellular dysregulation in aging populations has also been hypothesized to add to the pathogenesis of these diseases, which results in a damaged stem cell population.31,34 As neither cell dysregulation nor senescence is useful for research or the clinics, MSCs isolated from adult or geriatric populations are not an ideal cell source. MSCs isolated from younger patients have shown promise. 11
Recently we have gained access to small quantities of surplus bone from pediatric patients undergoing craniofacial reconstruction surgery from which we can easily isolate pediatric MSCs (P-MSCs). The resulting MSCs are plastic adherent, maintain MSC-related immunophenotype, and are capable of consistent differentiation. Here we outline how these cells are obtained, isolated, and cultured as well as describe the morphological and phenotypic characteristics of these novel MSCs to allow others in the scientific community to utilize them for their own applications. We compare P-MSCs with adult MSCs (A-MSCs) isolated from BM and find P-MSCs to be capable of more consistent multilineage differentiation. We believe P-MSCs to be a promising candidate for use in both research and clinical applications.
Materials and Methods
Mesenchymal stem cell isolation and expansion
All samples were harvested with the approval of the medical ethics committee at Erasmus Medical Centre (Erasmus MC, The Netherlands). A-MSCs were isolated and expanded as previously described (MEC-2004-142 and MEC-2015-644).
13
P-MSCs were isolated from leftover iliac crest bone chip material obtained from patients undergoing alveolar bone graft surgery (MEC-2014-16; 9–13 years). P-MSCs were isolated by gently swirling 10 mL of expansion medium (αMEM containing 10% serum [lot no. 41Q204K; Gibco], 50 μg/mL gentamycin, 1.5 μg/mL fungizone, 25 μg/mL
FACS analysis
A-MSCs and P-MSCs were trypsinized at passage 5 and rinsed in FACS flow. Cells were incubated for 30 min in 100 μL FACS buffer (BD Biosciences) containing antibodies against CD90 (APC), CD105 (FITC), CD73 (PE), CD271 (APC), CD166 (PE), HLA-DR (PerCP), HLA-ABC (FITC), or CD45 (PerCP). MSCs were washed with FACS flow, centrifuged at 689 g for 5 min, resuspended in 200 μL of FACS flow, and analyzed on a FACS Jazz flow cytometer (all antibodies BD Biosciences). Postanalysis was completed using FlowJo software version 10.0.7 (Treestar, Inc.).
Senescence staining and quantification
The percentage of senescent cells was determined by staining for senescence-associated lysosomal β-galactosidase using a modification of Debacq-Chainiaux et al.'s protocol.
37
A-MSCs and P-MSCs were seeded at 2300 cells/cm2 in complete expansion medium (as described in Mesenchymal stem cell isolation and expansion). On day 3, cells were refreshed and after 6 days, cells were fixed in 1% (v/v) formaldehyde (Sigma) and 0.5% glutaraldehyde (v/v) (Sigma) in PBS for 15 min at 4°C. After washing with distilled water, cells were incubated for 24 h at 37°C with 250 μL/cm2 staining solution (1 mg X-gal [5-bromo-4-chloro-3-indolyl-β-
Chondrogenic differentiation
A total of 2 × 105 A-MSCs or P-MSCs were suspended in 500 μL of chondrogenic medium (high-glucose Dulbecco's modified Eagle's medium [DMEM] supplemented with 50 μg/mL gentamycin [Invitrogen], 1.5 μg/mL fungizone [Invitrogen], 1 mM sodium pyruvate [Invitrogen], 40 μg/mL proline [Sigma], 1:100 v/v insulin-transferrin-selenium [ITS+; BD Biosciences], 10 ng/mL transforming growth factor β3 [Peprotech], 25 μg/mL
Osteogenic differentiation
A-MSCs or P-MSCs were plated at a density of 3 × 103 cells/cm2 in expansion medium (as described in Mesenchymal stem cell isolation and expansion). Twenty-four hours after seeding, medium was replaced with osteogenic induction medium (high-glucose DMEM supplemented with 10% serum, 50 μg/mL gentamycin, 1.5 μg/mL fungizone, 10 mM glycerol phosphate [Sigma], 0.1 μM dexamethasone [Sigma], and 0.1 mM
Adipogenic differentiation
A-MSCs or P-MSCs were plated at a density of 2.1 × 104 cells/cm2 in adipogenic induction medium (high-glucose DMEM supplemented with 10% serum, 50 μg/mL gentamycin, 1.5 μg/mL fungizone, 0.2 mM indomethacin [Sigma], 0.01 mg/mL insulin [Sigma], 0.5 mM 3 iso-butyl-1-methyl-xanthine [Sigma]). The medium was refreshed as previously described and cells were maintained at 37°C and 5% CO2. Samples were prepared for histology (fixed in 4% [w/v] formalin) after harvest.
Hematoxylin–eosin staining
Pediatric bone chips were fixed for 24 h in 4% (w/v) formalin, decalcified in 10% EDTA (w/v) for 30 days, and paraffin embedded. Chondrogenic MSC pellets were fixed in 4% (w/v) formalin for 2 h and paraffin embedded. Six micrometer-thick sections were cut, deparaffinized, and stained with hematoxylin–eosin (H&E). H&E staining was performed by incubating deparaffinized samples in Gill's hematoxylin (Sigma) for 5 min, washed in nondistilled water for 5 min, washed in distilled water, and counterstained for 45 s with 2% eosin (Merck; in 50% ethanol, 0.5% acetic acid). Samples were fixed in 70% EtOH for 10 s and rehydrated (sequentially in 96% EtOH, 100% EtOH, and xylene for 1 min). Samples were mounted in Entellan (Depex).
Thionine staining
Deparaffinized samples were incubated in 0.04% thionine (prepared in 0.01 M sodium acetate, pH 4.5) for 5 min, differentiated in 70% EtOH for 10 s, and then rehydrated as previously described. Samples were mounted in Entellan (Depex).
von Kossa staining
Osteogenically differentiated MSCs were fixed in 4% (w/v) formalin for 1 h. After a rinse with ultrapure water, samples were incubated in 5% w/v silver nitrate (Sigma) for ∼30 min under direct light provided by a light box. After incubation, samples were washed in ultrapure water and counterstained with nuclear fast red (Merck) for 5 min. Samples were dehydrated in 70% EtOH for 10 s followed by 96% EtOH and 100% EtOH for 1 min. Samples were imaged in 100% EtOH directly after staining.
Oil red staining
After a 15-min fixation in 4% (w/v) formalin, samples were rinsed in distilled water for 10 min. Then 0.5% w/v Oil-red O (in 2-propanol; Sigma) was added to samples for 10 min followed by further rinsing with distilled water. Samples were imaged in distilled water.
P-MSC–peripheral blood mononuclear cells coculture
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats from healthy male donors (Sanquin, Rotterdam) by a Ficoll-Paque PLUS gradient separation as previously described 38 (GE Healthcare). PBMCs were resuspended in human serum-conditioned medium (RPMI-1640 medium, 1% GlutaMAX [Life Technologies], 50 μg/mL gentamycin, 1.5 μg/mL fungizone, 10% human serum [Sigma-Aldrich]) and stored at −80°C until use. P-MSCs were trypsinized as previously described and seeded in low-evaporation round-bottomed 96-well plates. Twenty-four hours after seeding, 1 × 107 PBMCs were labeled with 20 μL of carboxyfluorescein diacetate succinimidyl ester (CFSE). T cells were stimulated by adding anti-CD3/CD28 antibodies (1 μL/106 cells) to PBMC suspension with an antigoat linker antibody (2 μL/106 cells). Stimulated 100,000 PBMCs were cocultured with P-MSCs at P-MSC:PBMC ratios of 1:2.5, 1:5, 1:10, or 1:20. PBMCs were harvested 5 days later and stained with CD3-PerCP (clone SK7), CD8- PE-Cy7, and CD4-APC (BD Biosciences). Samples were fixed in 3.6% paraformaldehyde and analyzed using a FACS Jazz flow cytometer (BD Biosciences) and postanalysis was completed using FlowJo software version 10.0.7 (Treestar, Inc.). Data are represented as reciprocal of the mean fluorescence intensity.
Statistical analysis
Mann–Whitney U analysis was performed using SPSS (Ver 21.; SPSS, Inc., Chicago) on data shown in Figures 1–3. Kruskal–Wallis analysis with Dunn's multiple comparison was performed as shown in Figure 4. Data are shown as mean ± SD, except for Figure 5 which is mean ± SEM, p-values under 0.05 were considered significant.

P-MSCs are isolated from illiac crest rest material containing both bone and bone marrow. Hematoxylin and eosin-stained sections of the illiac crest chips from which P-MSCs are isolated. The cell source environment is rich in both bone marrow elements and bone (black arrows indicate bone, white arrows indicate bone marrow). MSC, mesenchymal stem cells/marrow stromal cells; P-MSCs, pediatric MSCs. Color images available online at
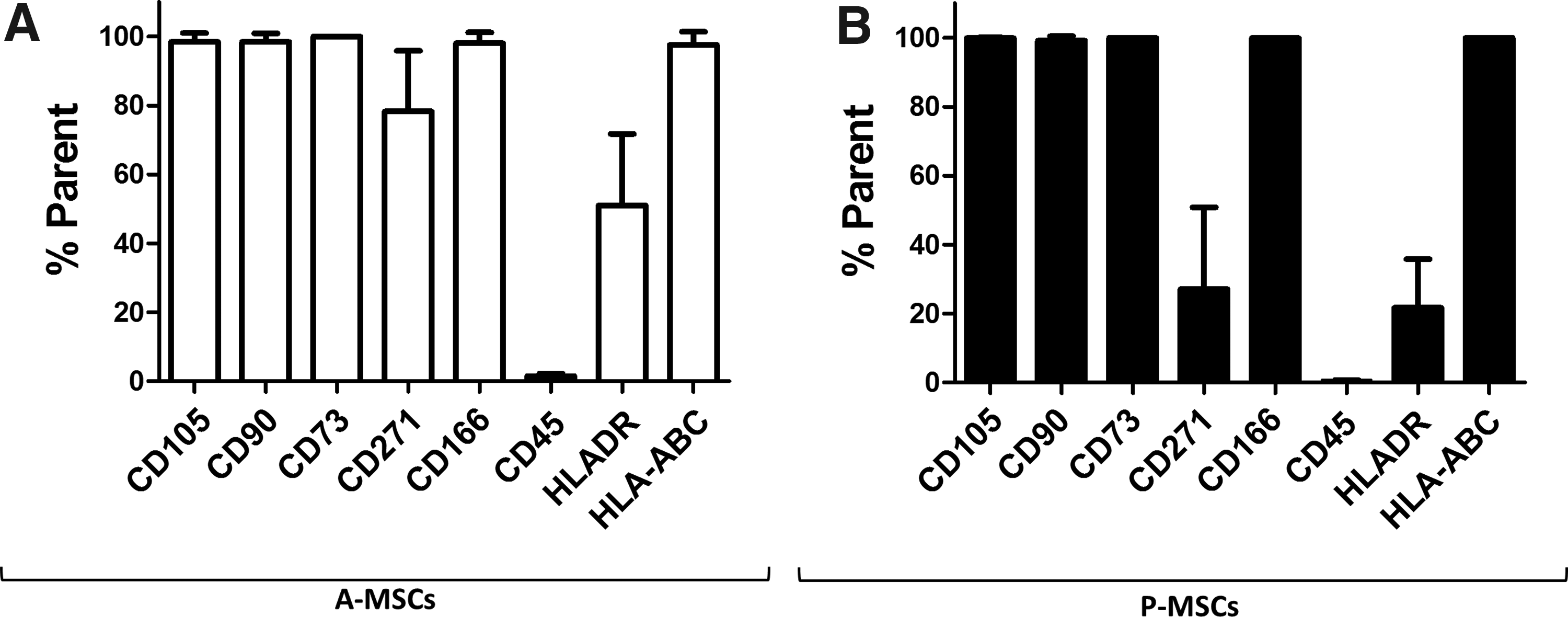
P-MSCs and A-MSCs express similar levels of general stem cell markers. General MSC-related markers that are commonly expressed on A-MSCs
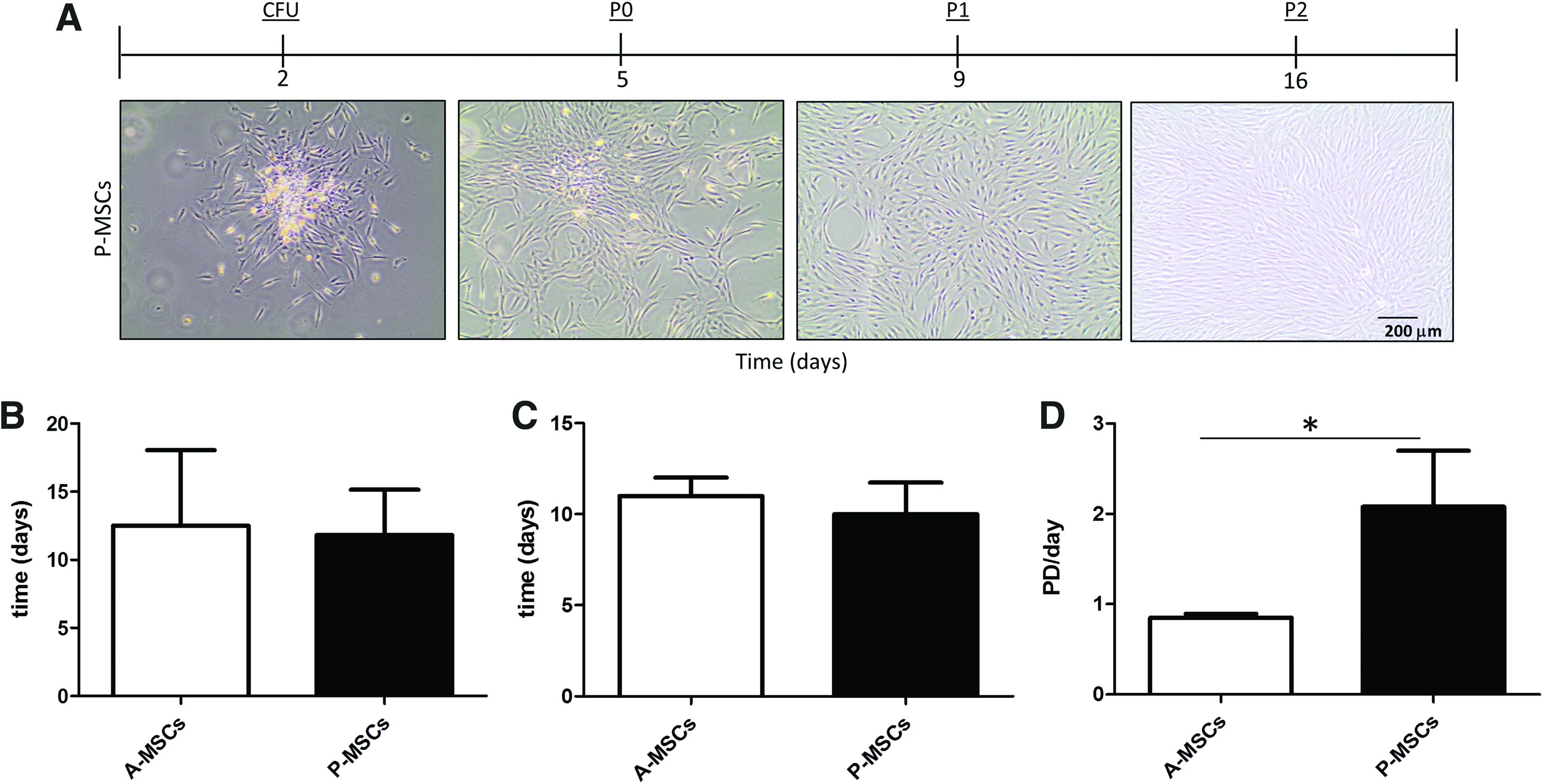
P-MSCs and A-MSCs have a similar rate of expansion but P-MSCs undergo more population doublings.
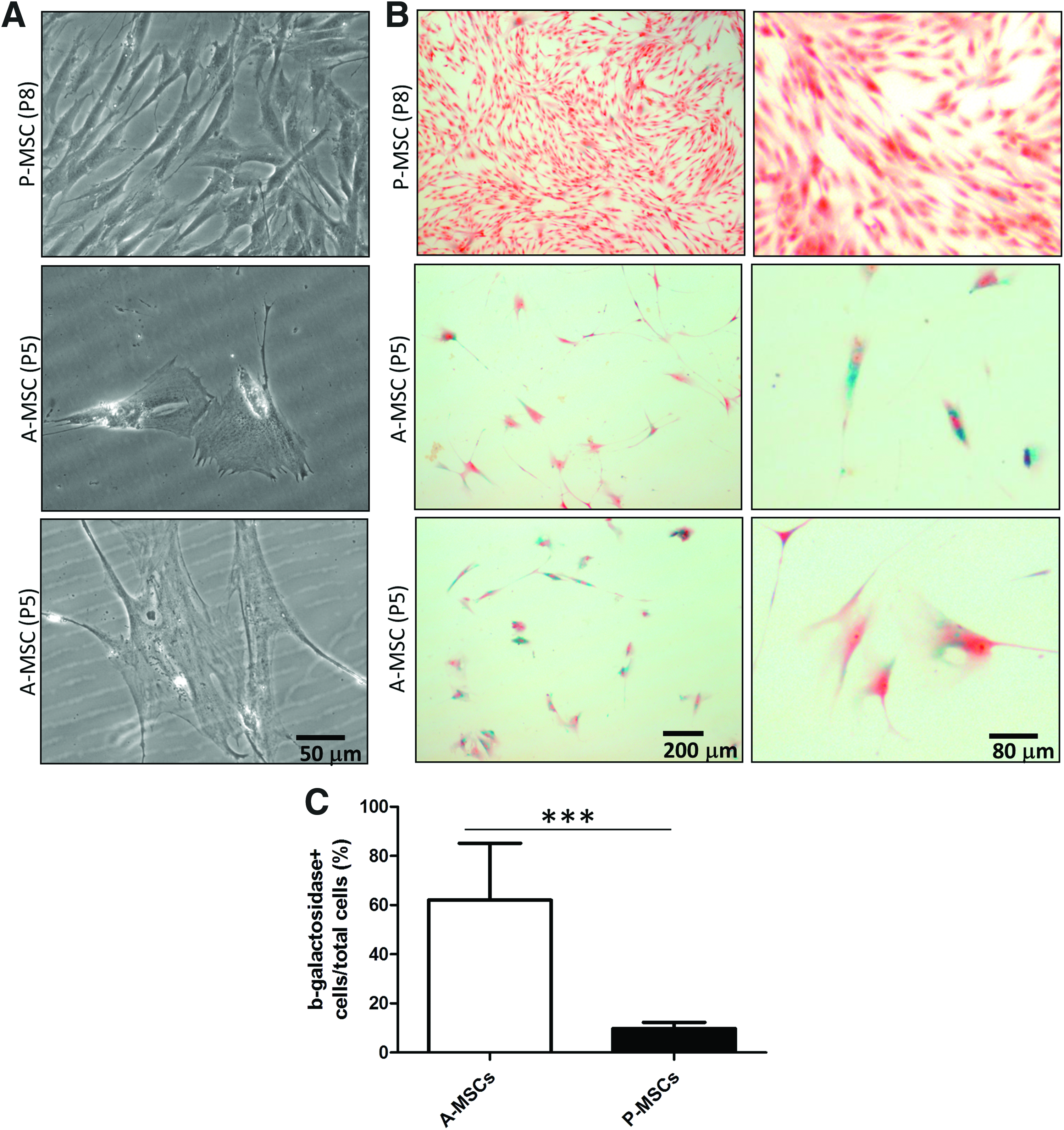
P-MSCs contain significantly less senescent cells than A-MSCs.
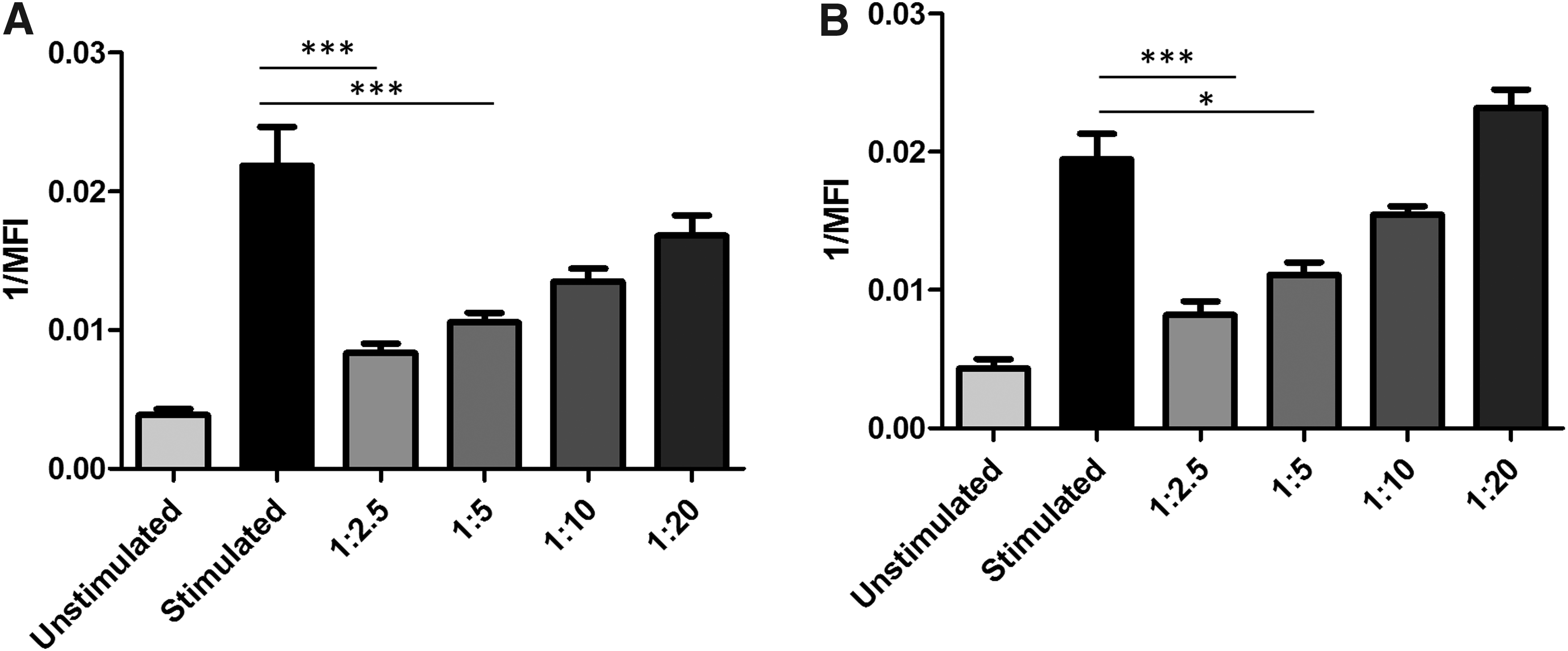
P-MSCs reduce allogeneic CD4+ and CD8+ T cell proliferation in monolayer. Stimulated (+CD3/CD28) PBMCs cocultured with P-MSCs at different MSC:PBMC ratios (1:2.5, 1:5, 1:10, 1:20). CD4+
Results
P-MSCs express a panel of established MSC surface markers
P-MSCs were isolated from small quantities of surplus bone biopsies from patients undergoing cleft pallet reconstruction surgery. The environment from which the P-MSCs are isolated contains both bone and BM elements (Fig. 1). To prove that isolated cells were indeed true MSCs, we characterized the immunophenotypic expression of common MSC markers. These markers included a panel that are known to be expressed on MSCs, including CD105, CD90, CD73, CD271, CD166, and HLA-ABC as well as a commonly used negative marker, lymphocyte-associated CD45.12,39 Both A-MSCs and P-MSCs were analyzed after five passages. MSC markers were expressed at a similar level in P-MSCs and A-MSCs (Fig. 2). Both populations were negative for CD45 and positive HLA-ABC. About half the P-MSCs and A-MSCs population were positive for HLA-DR, which was not surprising as HLA-DR expression can increase with in vitro culture of MSCs. 40 We found no significant difference in HLA-DR expression between P-MSCs and A-MSCs (Fig. 2).
P-MSCs have enhanced expansion properties compared with A-MSCs
During expansion, P-MSCs exhibit a typical MSC morphology similar to that observed in A-MSCs (Fig. 3A). Although the total number of days A-MSCs and P-MSCs took to establish the initial culture (Fig. 3B) and reach passage 3 (Fig. 3C) did not change, P-MSCs expanded significantly faster than A-MSCs, yielding more cells after the same time in culture (Fig. 3D; p < 0.02). This difference in cell number could be contributed to cell size. A-MSCs enlarged the longer they were in culture, whereas P-MSCs remained small (Figs. 3A and 4A).
P-MSCs are a less senescent cell source compared with A-MSCs
As increased cell size is a hallmark of senescence, a permanent cell cycle arrest that A-MSCs have been shown to undergo in vitro, we compared senescence between A-MSCs and P-MSCs.41,42 We observed cell enlargement of A-MSCs compared with P-MSCs (Fig. 4A). Senescence-associated lysosomal β-galactosidase staining showed that P-MSCs, even after extensive passage (p8; Fig. 4B, C), contained significantly less senescent cells than A-MSCs at an earlier passage (p5; Fig. 4B, C).
In monolayer, P-MSCs reduce T cell proliferation at a similar level as A-MSCs
Our laboratory has previously shown that A-MSCs repress the proliferation of allogeneic T cells, a feature essential for many anti-inflammatory MSC-based therapeutics. 38 To examine whether P-MSCs repress T cell proliferation, allogeneic T cells from PBMC fractions were CD3/CD28 stimulated and added in suspension to P-MSC monolayers. In both CD4+ and CD8+ T cell subsets, P-MSCs inhibited T cell proliferation in a dose-dependent manner and to a similar extent as we have previously reported for A-MSCs (Fig. 5). Here we found that P-MSCs exhibited a similar level of inhibition to what was reported by A-MSCs. 38
P-MSCs exhibit more consistent multilineage differentiation capacity compared with adult donors
A-MSCs are known to exhibit inconsistent differentiation capabilities that vary greatly between donors. This severely limits their use in applications wherein consistency is essential. 43 Compared with A-MSCs donors (passages 2–5; Fig. 6A), the trilineage differentiation potential of P-MSCs (passage 5; Fig. 6B) was found to be more consistent. Even though P-MSCs were used several passages beyond that used for the A-MSCs, P-MSCs more consistently underwent adipogenesis, osteogenesis, and chondrogenesis, with only 1 out of 12 donors not being able to make bone or cartilage after treatment (Fig. 6B). A-MSCs showed much more variability in their differentiation potential. Out of the 14 A-MSC donors tested, 3 donors failed to undergo adipogenesis, 5 donors failed to osteogenically differentiate, and 5 donors were unable to chondrogenically differentiate (Fig. 6A).

P-MSCs show more consistent capacity for multilineage differentiation.
Discussion
Identifying cell sources with enhanced and reliable differentiation capabilities and expansion properties is necessary in order for MSCs to be more readily utilized both in research and in ATMPs. Here we have established a simple isolation protocol to obtain P-MSCs from surplus iliac crest bone chip material (Fig. 1). We confirmed P-MSCs expressed cell surface markers typically used to properly identify MSC populations. P-MSCs showed similar expression levels of general stem cell markers CD105, CD90, CD73, and CD166 compared with A-MSCs, as well as the absence of the hematopoietic marker CD45 (Fig. 2). As these markers are conventionally used to identify MSCs,44,45 we are confident that this marker expression in combination with other results confirms their phenotype. In this study, A-MSCs displayed a higher expression of CD271 and HLA-DR than P-MSCs. It has been previously reported that in MSCs, CD271 as well as HLA-DR expression decrease over time in culture.46,47 As P-MSCs undergo more population doublings than A-MSCs at the same passage (Fig. 3D), this increased cellular division could have contributed to the decreased expression of both CD271 and HLA-DR we observe in P-MSCs.
P-MSCs expanded more rapidly than A-MSC donors, which might be attributed to the relatively low senescence in P-MSCs (Fig. 4). Senescent cells are much larger than nonsenescent cells. 48 A-MSCs have more enlarged senescent cells that do not divide,49,50 making it easy to understand why the population doublings in A-MSCs are effected. In addition, by preventing proliferation, senescence can also block differentiation pathways requiring proliferation, such as chondrogenic differentiation. 51 Being able to obtain cells with higher proliferation and differentiation capacity in a shorter time than is possible with A-MSCs makes P-MSCs an attractive cell source.
If P-MSCs are to be utilized in an allogeneic setting, it is important to show P-MSCs maintain immunomodulatory capabilities typically observed in A-MSCs.38,52 MSCs are known to be able to manipulate T cell proliferation and phenotypic behavior, and their immunosuppressive nature makes them an interesting candidate from a clinical perspective.52–54 In this study, we found P-MSCs were capable of inhibiting T cell proliferation at a level similar to what we previously reported with A-MSCs using a 1:5 ratio (MSC:PBMCs). 38 For use in an allogenic model, it is advantageous for P-MSCs to inhibit T cell proliferation to prevent an unwanted immune reaction after transplantation. However, how P-MSCs interact with other immune cell types including antigen presenting cells needs to be determined to further understand how they would respond to a fully functional immune system.
A-MSCs have been reported to have inconsistent multilineage differentiation capabilities.15,20,55 P-MSCs were capable of more consistent multilineage differentiation than the A-MSCs in this study. Senescence could have contributed, in part, to the difference we observed here as senescent MSC populations undergo phenotypic changes56,57 and exhibit chromosomal abnormalities,50,58 which ultimately could influence their differentiation capacity. 51 In this study, it is plausible that a combination of factors influenced the differentiation capacity of these cell populations. It is logical that cells from a younger, healthier patient who contain less senescent cells would be capable of better multilineage differentiation than senescent cells obtained from elderly patients. Having a cell source with more consistent differentiation capacities is ideal as it allows for more reproducible results.
Here we have described an easy isolation protocol that allows access to a P-MSC population with enhanced expansive and differentiation potential compared with A-MSCs. P-MSCs with their rapid expansion, low senescence, and consistent multilineage differentiation are, therefore, prime candidates for applications from drug screening and development to use in ATMPs.
Footnotes
Acknowledgments
The authors thank Dr. Niamh Fahy at the Erasmus MC for assistance revising this article. This research was supported by the AO Foundation, Switzerland (AOCMF-15-27F). R.N. was further supported by the VENI grant by STW (13659). J.L. was further supported by the Erasmus MC grant.
Disclosure Statement
No competing financial interests exist.