
Abstract
Recent advances in the development of various culture platforms are promising for achieving more physiologically relevant in vitro hepatic models using primary human hepatocytes (PHHs). Previous studies have shown the value of PHHs three-dimensional (3D) spheroid models, cultured in low cell number (1330–2000 cells/3D spheroid), to study long-term liver function as well as pharmacological drug effects and toxicity. In this study, we report that only plateable PHHs aggregate and form compact 3D spheroids with a success rate of 79%, and 96% reproducibility. Out of 3D spheroid forming PHH lots, 65% were considered stable (<50% ATP decrease) over the subsequent 14 days of culture, with reproducibility of a given PHH lot being 82%. We also report successful coculturing of PHHs with human liver nonparenchymal cells (NPCs). Crude P1c-NPC fractions were obtained by low centrifugation of the PHH supernatant fraction followed by a few days of culture before harvesting and cryopreservation. At aggregation of PHHs/P1c-NPCs (2:1 ratio 3D spheroids), liver sinusoidal endothelial cells, Kupffer cells, and hepatic stellate cells were successfully integrated and remained present throughout the subsequent 14-day culture period as revealed by mRNA expression markers and immunostaining. Increased mRNA expression of albumin (ALB), apolipoprotein B (APOB), cytochrome P450 3A4 (CYP3A4), and increased albumin secretion compared to PHH 3D spheroid monocultures highlighted that in a 3D spheroid coculture, configuration with NPCs, PHH functionality is increased. We thus achieved the development of a more integrated coculture model system requiring low cell numbers, of particular interest due to the scarcity of human liver NPCs.
Impact Statement
This is, to our knowledge, the first report on a systematic evaluation of primary human hepatocyte lots to form spheroids. It is also the first report on the success of liver nonparenchymal cells cryopreserved after a culture step to stably integrate with primary human hepatocytes as 3D spheroids.
Introduction
R
Liver cells comprise ∼60% hepatocytes that are primarily responsible for drug metabolism and have a range of functions. 11 However, the liver is a complex unit with multiple cell subtypes working together in cohesion, and the contribution of NPCs is not accounted for in monocultures of hepatocytes. NPCs, namely liver sinusoidal endothelial cells (LSECs), hepatic stellate cells (HSCs), and Kupffer cells (KCs) do not only provide support for hepatocytes, but also contribute to inflammatory responses by their own specialized functions within the liver. LSEC line the hepatocytes and play a significant role in the transport of molecules from the circulating blood to the hepatocytes. 12 HSCs are found in the space of Disse and are known to be involved in the formation of extracellular matrix,13,14 and KCs provide inflammatory cues and responses to stimuli. 15 Recent studies have highlighted the importance of NPCs and their contribution to drug toxicity and overall liver responses with PHH/NPC cocultures being shown to be a suitable in vitro model to study these interactions.6,7,16
In this study, we report that plateability is a prerequisite for PHHs to aggregate and form compact 3D spheroids using ultralow attachment (ULA) plates, a method first reported by Bell et al. 7 A success rate of 79% was observed; of those, 65% being stable over the subsequent 14 days of culture. We also report that when mixing PHHs with a crude P1c-NPC fraction, HSCs, KCs, and LSECs were successfully integrated into the PHH/P1c-NPC 3D spheroids at aggregation and remained present throughout the 14-day culture period. PHH/P1c-NPC 3D spheroids could be stimulated by lipopolysaccharide (LPS) to secrete interleukin-6 (IL-6), and this was associated with increased trovafloxacin toxicity compared to PHH monoculture 3D spheroids. Increased mRNA expression of albumin (/ALB), apolipoprotein B (/APOB), and cytochrome P450 3A4 (/CYP3A4), as well as increased ALB secretion compared to PHH 3D spheroid monocultures revealed that in a 3D spheroid PHH/NPC coculture configuration, PHH functionality is increased.
Materials and Methods
Isolation and cryopreservation of PHHs and NPCs
PHHs and NPCs were isolated from healthy human liver tissue, obtained from partial hepatectomy or nontransplantable livers. The human biological samples were acquired in agreement with French ethic regulations, and their research use was in accordance with the terms of the informed consents. PHHs and NPCs were isolated as described by Alexandre et al.
17
and Drucker et al.,
18
respectively. PHHs were cryopreserved as previously reported
17
and NPCs were either immediately cryopreserved (P0c-NPCs) or cultured (P0f-NPCs) for several days before being detached by trypsin and cryopreserved (P1c-NPCs) (Supplementary Fig. S1; Supplementary Data are available online at
PHH 3D spheroid monoculture and PHH/NPC 3D spheroid coculture
Cryopreserved PHHs from 45 donors (Supplementary Table S1) were thawed and seeded at 1500 cells per well into 96-well ULA plates as previously described.
7
Cryopreserved PHHs (lots S1391T or SSR) were cocultured with cryopreserved NPCs (P1c-NPCs, lots HC1565, HC1559, S1478, S1493, S1568). A total of 2000 viable cells were seeded at a ratio of 2:1 (PHHs:NPCs) per well into 96-well ULA plates
7
in William's E medium supplemented with 2 mM
Assessment of viability, albumin secretion, and size
At aggregation (D0) and following culture periods (D7 and/or D14), viability was determined by measuring cellular ATP using the CellTiter-Glo® Kit (Promega, France). Blank values were subtracted from all wells. At the same time points, cell culture medium was collected, and albumin concentrations were determined using an enzyme-linked immunosorbent assay (ELISA) kit (Bethyl Laboratories). Size of 3D spheroids was estimated by ImageJ software.
Immunostaining
After isolation (P0f-NPCs) or thawing (P0c-NPCs or P1c-NPCs), NPCs were seeded in 2D culture for 2–3 days, and monolayers were fixed with 4% paraformaldehyde for 10 min at +4°C. After permeabilization with 0.5% Triton-X100, cells were immunostained with specific antibodies against: CD31 for LSECs, 6 GFAP (for glial fibrillary acidic protein) for HSCs, 19 and CD68 for KCs.6,7,20 Fluorescent images were acquired using a ZOE Fluorescent Cell Imager (Bio-Rad, France).
Three-dimensional spheroid monocultures (SSR) and cocultures (P1c-NPC lot S1493) were collected at day 7 and 14 postaggregation. Samples, fixed in 4% paraformaldehyde for 16 h at +4°C, were cryoprotected in 30% sucrose and embedded in Tissue-Tek OCT compound (Sakura, The Netherlands). Three-dimensional spheroid cryosections (8 μm) were immunostained with primary antibodies against: Von Willebrand factor (vWF) for LSECs (Dako, Denmark 21 ), vimentin (VIM) for HSCs (Abcam, United Kingdom), CD68 for KCs (Santa Cruz Biotechnology), and CYP3A4 for PHHs (Cypex Limited, United Kingdom). Following secondary antibody staining with goat anti-rabbit Alexa Fluor 488 (Thermo Fisher Scientific) and donkey anti-mouse Alexa Fluor 555 (Thermo Fisher Scientific), slides were mounted with ProLong Gold Antifade Mountant with DAPI (Life Technologies). Fluorescence imaging was performed using an LSM710 confocal microscope (Zeiss, Germany) and processed using Zen 2.5 blue edition analysis software.
Interleukin-6 secretion
Functionality of KCs was assessed by measuring LPS-induced IL-6 secretion. At the day of aggregation (D0), LPS was added to the medium of PHH 3D spheroid monocultures or PHH/NPC 3D spheroid cocultures at a concentration of 10 μg/mL and supernatants were collected after 48 h and stored at −80°C. IL-6 concentration was determined using an ELISA kit (Euromedex, France). Results are expressed as a percentage over the PHHs monocultured in absence of LPS.
Cytotoxicity assays
At the day of aggregation (D0), trovafloxacin at various concentrations was added to the medium of PHH 3D spheroid monocultures or PHH/NPC 3D spheroid cocultures with or without LPS (10 μg/mL). After 48 h of exposure, cell viability was assessed by the CellTiter-Glo kit and expressed as the percentage of ATP content in the treated cells compared with vehicle control.
At the day of aggregation (D0), fialuridine at various concentrations was added to the medium of PHH 3D spheroid monocultures. Treatment was repeated every 2–3 days and cell viability was assessed after 3, 7, and 14 days of exposure by the CellTiter-Glo kit and expressed as the percentage of ATP content in the treated cells compared with vehicle control.
Reverse transcriptase and quantitative polymerase chain reaction analysis
Total RNA was extracted from cells at different stages using the Direct-zol™ RNA MiniPrep Kit (Zymo Research). RNA was quantified by fluorimetry using the QuantiFluor RNA System (Promega) and cDNAs were obtained from 0.07 μg total RNA using the iScript Reverse Transcription Supermix for RT-qPCR from Bio-Rad. Expression levels of PHH markers ALB/ALB, 22 APOB/APOB and CYP3A4/CYP3A4, LSEC markers CD31/PECAM1 and lymphatic vessel endothelial hyalurinan receptor 1 (LYVE1/LYVE1), HSC makers vinculin (VCL/VCL), 20 VIM/VIM 7 and alpha smooth muscle actin (αSMA/ACTA1), 23 and KC markers CD163/CD163 20 and CD68/CD68 7 were determined with a set of primers designed (Supplementary Table S2) and marketed by Eurogentec (Belgium). All markers were normalized with the expression of the housekeeping gene actin B (ACTB). At thawing, the expression of PHH gene markers was compared to the level obtained in NPCs and the expression of respective NPC gene markers was compared to the level obtained in PHH monocultures. At 3D spheroid aggregation (D0) and 14 days thereafter (D14), the expression of PHH and NPC markers in PHH/NPC cocultures was compared with PHH monocultures from same day. Expression was normalized taking into account the ratio 2:1 of PHHs/NPCs.
Results
PHH 3D spheroid monocultures
Success rate of aggregation of cryopreserved PHHs
We and others have noticed that PHHs, freshly isolated or cryopreserved, do not systematically aggregate to form 3D spheroids. We therefore wanted to assess if the aggregation potential of a given cryopreserved PHHs lot was dependent on its plateability in 2D culture and tested 45 lots known for their plateability on collagen-coated culture wells (Supplementary Table S1). We found that none of the six lots with <80% confluency in 2D culture wells aggregated when seeded in ULA plates, while 31/39 with ≥80% confluency in 2D culture wells aggregated to form 3D spheroids. Figure 1 depicts an example of the difference in performance of PHH lots during the aggregation process. Reproducibility in aggregation for seven PHH lots was 96%.

Bright-field images of plateable or suspension PHHs (S1518T and HC1559T, respectively) during the aggregation process. Between 1330 and 1500 PHHs per well were seeded, the aggregation process takes 5–7 days. 7 Scale bar = 100 μm. PHHs, primary human hepatocytes.
Morphological and functional stability of 3D spheroids over time in culture
The 31 lots which formed compact single 3D spheroids were maintained in culture over an additional period of 14 days. Lots for which ATP content did not decrease over 50% during the 14 days of culture postaggregation (20/31) were considered stable. 7 Figure 2 shows that for stable cultures, on D14 the average 3D spheroid size was 92% ± 6% relative to D0, while ATP content decreased to 64% ± 6%. Reproducibility in stability over time in culture postaggregation for seven tested PHH lots was 82%.

Morphological and functional stability of 3D spheroids over time in culture. Size (gray bars, n = 18) and ATP content (black bars, n = 20) after 7 days (D7) and 14 days (D14) of culture postaggregation (D0). Data represent mean ± SEM compared to D0. 3D, three-dimensional; SEM, standard error of the mean.
Properties and responses to toxic compounds of 3D spheroids cultivated in ULA plates compared to hanging-drops
Figure 3 shows that 3D spheroid morphology and size were similar when obtained using either ULA plates or the GravityTRAP™ plates for the same PHH lot (B1370T). In both cases, ATP content/3D spheroid was stable from aggregation and subsequent culture time up to 14 days of culture. In addition, as depicted in Figure 4, the response to fialuridine at various concentrations over time was similar in both 3D culture formats. This observation was confirmed with five additional training compounds tested within the EU-funded MIP-DILI consortium (data not shown).
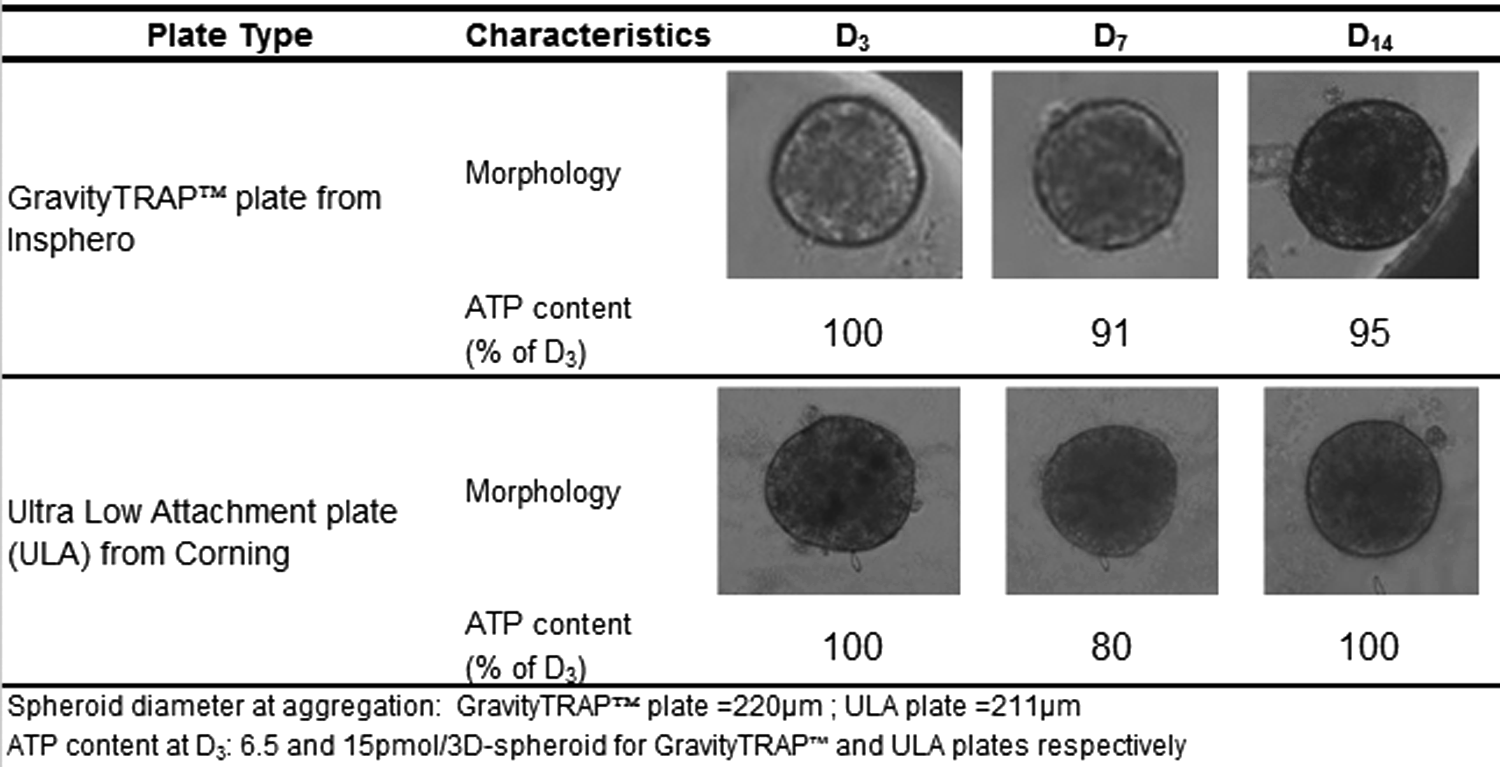
Typical morphology and relative ATP content of 3D spheroids from lot B1370T seeded in GravityTRAP™ or ULA plates 3 days (D3), 7 days (D7) and 14 days (D14) after aggregation. ULA, ultralow attachment.
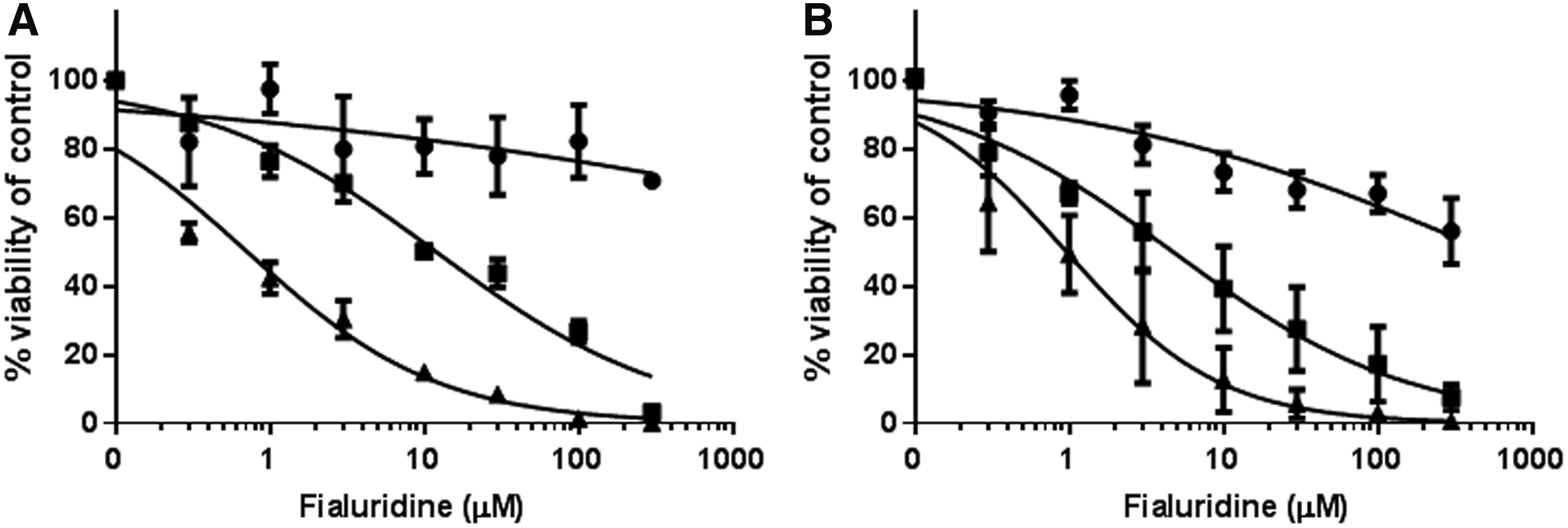
Cell viability profile of 3D spheroids from lot B1370T seeded in GravityTRAP
PHH and liver NPC 3D spheroid cocultures
NPC fraction preparation and cell type characterization
Crude fractions of liver NPCs were isolated from liver resections as described in Materials and Methods section (Supplementary Fig. S1) and processed as depicted in Figure 5. The enriched P0f-NPC fractions were contaminated with hepatocytes at seeding in culture flasks. After 5–10 days of culturing, hepatocytes progressively disappeared. Immunostaining showed that markers for LSECs (CD31), 6 HSCs (GFAP), 19 and KCs (CD68) 20 were maintained throughout the culture period. After cryopreservation, P0c-NPC fractions did not attach to culture plates, while P1c-NPCs did with immunostaining confirming the presence of the three cell types.
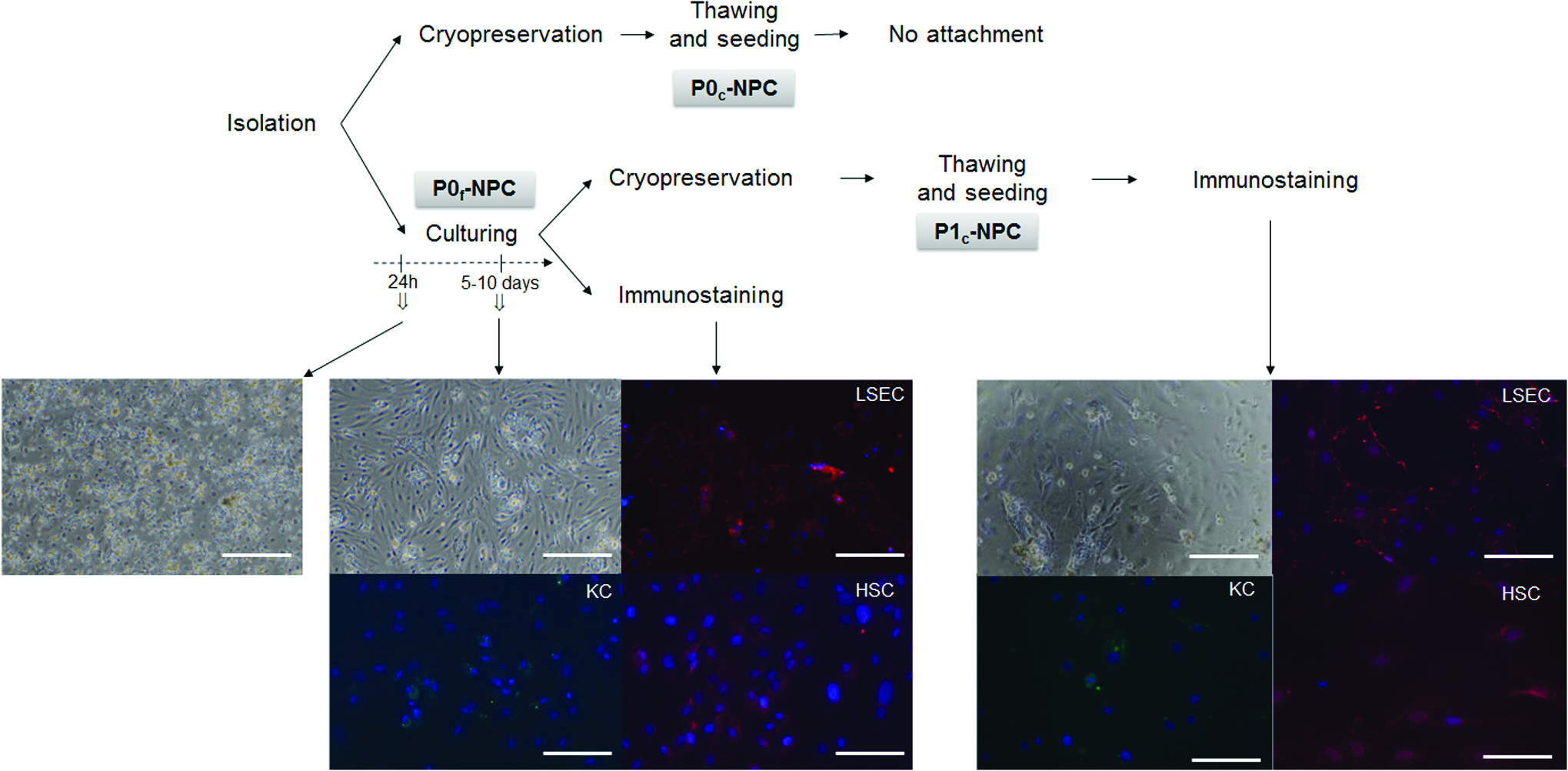
Seeding after isolation, cryopreservation, and thawing of NPCs. After isolation, NPCs were seeded (P0f-NPCs) and/or cryopreserved (P0c-NPCs). After 5–10 days in culture, part of the P0f-NPCs was immunostained and the other part was cryopreserved. The latter was thawed (P1c-NPCs), seeded, and after 3–4 days of culture, immunostained. Cell morphology was observed by phase contrast microscopy using a Nikon Diaplot TMD. NPCs were identified by immunofluorescence staining of the different cell types: CD31 for LSECs (red), CD68 for KCs (green) and GFAP for HSCs (red) using a ZOE Fluorescent Cell Imager (Bio-Rad, France). Nuclei were counterstained with DAPI (blue). Scale bar = 100 μm. GFAP, for glial fibrillary acidic protein; HSCs, hepatic stellate cells; KCs, Kupffer cells; LSECs, liver sinusoidal endothelial cells; NPCs, nonparenchymal cells. Color images available online at
RT-qPCR quantification of ALB, APOB, and CYP3A4 gene expression confirmed that P0c-NPC fractions were largely contaminated by PHHs, while this was substantially reduced in P1c-NPC fractions (Fig. 6). KC markers CD163 and CD68 20 were present in the P1c-NPC fractions, although enrichment was less compared to the P0c-NPC fractions. Expression of the LSEC marker PECAM16,20 and HSC marker VIM 7 were present in both P0c-NPC and P1c-NPC fractions, their expression being higher in the latter, while the expression of LSEC marker LYVE1 and of HSC marker VCL 20 were almost lost in P1c-NPC fractions. The same was observed with ACTA1, a marker of activated HSCs24,25 that was much lower in P1c-NPC compared to P0c-NPC fractions. A marked lot-to-lot variability in the enrichment of all NPC types was observed.
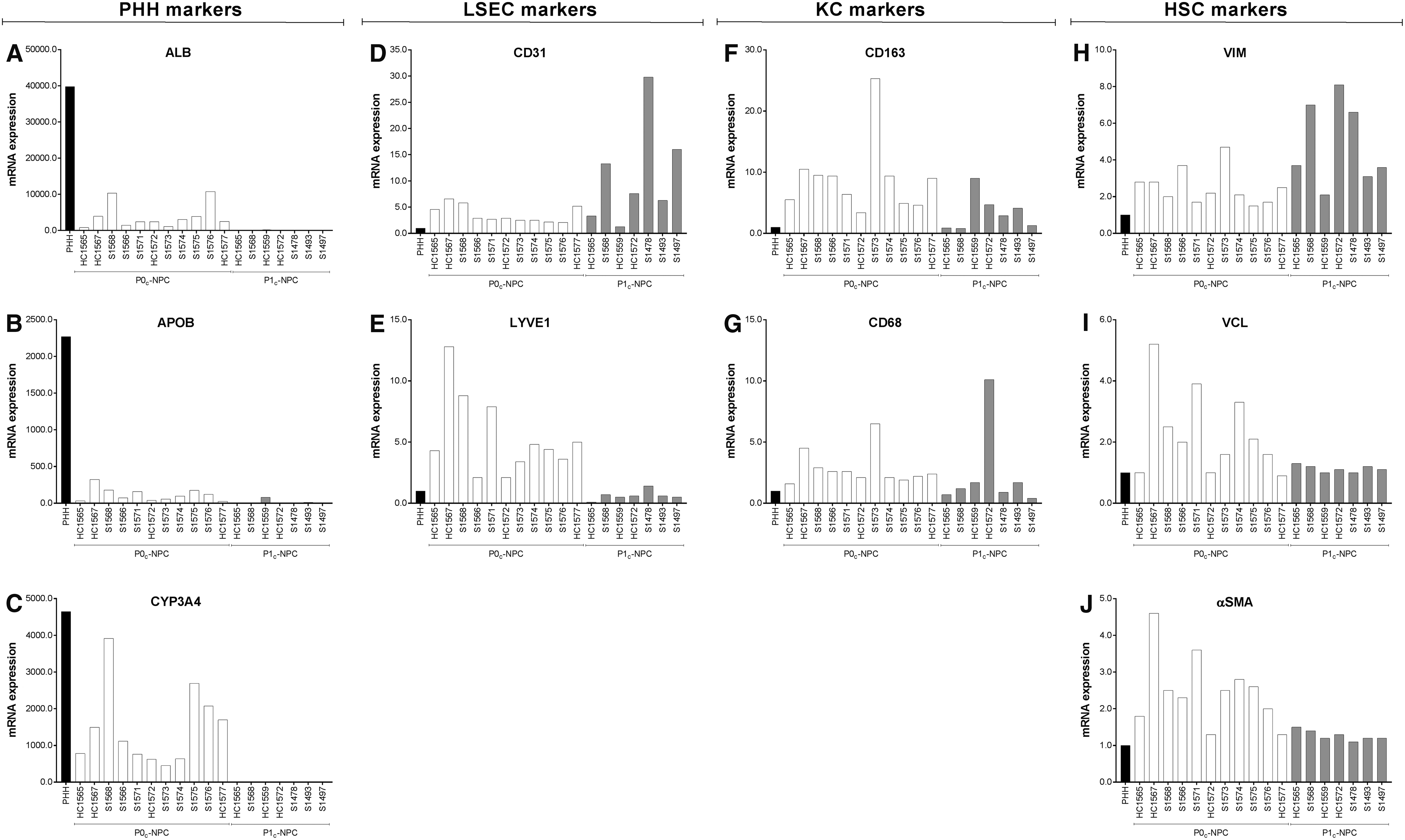
Freshly thawed PHHs (black bar), P0c-NPCs (n = 11, white bars) and P1c-NPCs (n = 7, gray bars) were characterized by RT-qPCR in simplicate. All markers were normalized with the expression of the housekeeping gene actin B (ACTB).
20
For PHH gene markers [ALB/ALB
NPC incorporation into PHH/NPC 3D spheroids
Size and viability of 3D spheroid cocultures
Thawed crude fractions of P1c-NPCs were mixed with thawed PHHs (lot S1391T), seeded in ULA plates, and assessed for aggregation as described in Materials and Methods section. Figure 7 shows that the addition of P1c-NPC fractions of five lots to PHHs slightly increased the size of 3D spheroids, which ranged between 106% and 121% of PHH 3D spheroids. ATP content ranged between 86% and 114% of PHH 3D spheroids at aggregation. After 14 days of culture postaggregation, PHH/NPC 3D spheroid size and ATP content ranged, respectively, between 102–113% and 97–149% of PHH 3D spheroids (data not shown).
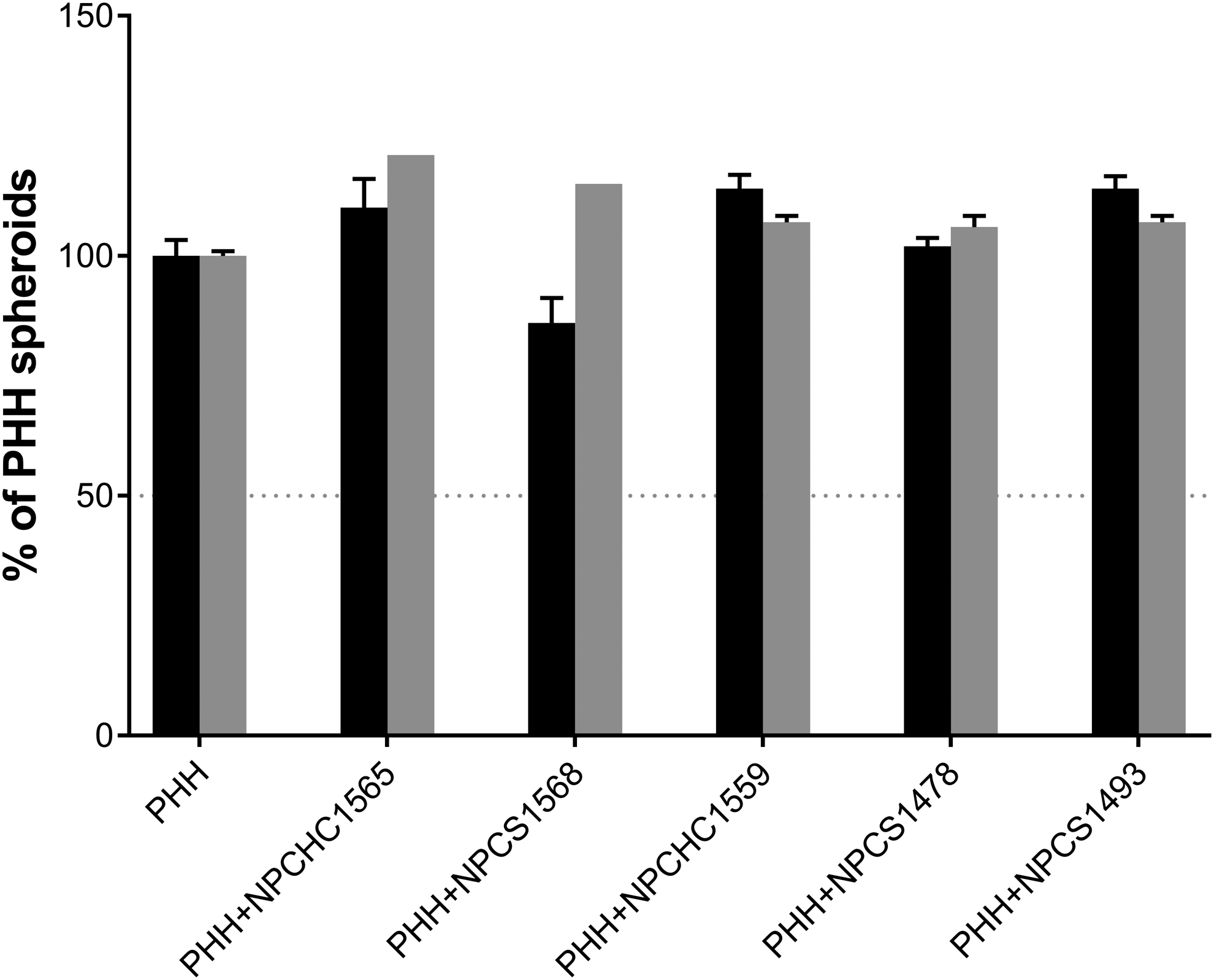
Properties of PHH/P1c-NPC 3D spheroid cocultures at aggregation (D0). ATP content (black bars) and size (gray bars) of 3D spheroids (n = 1–9) were expressed as % ± SEM of PHH monoculture 3D spheroids (n = 12–30).
Enrichment of 3D spheroids with various NPC types
The expression of the various NPC markers shows that an enrichment of all cell types occurred in 3D spheroid PHH/P1c-NPC cocultures at aggregation (D0) and that this enrichment remained altogether stable over the culture period, up to day 14 postaggregation (D14) (Fig. 8). There was some variability between the P1c-NPC lots tested, but enrichment was at least threefold compared to PHH monocultures. Immunostaining confirmed the presence of all cell types in 3D spheroid PHH/P1c-NPC cocultures up to D14 postaggregation.
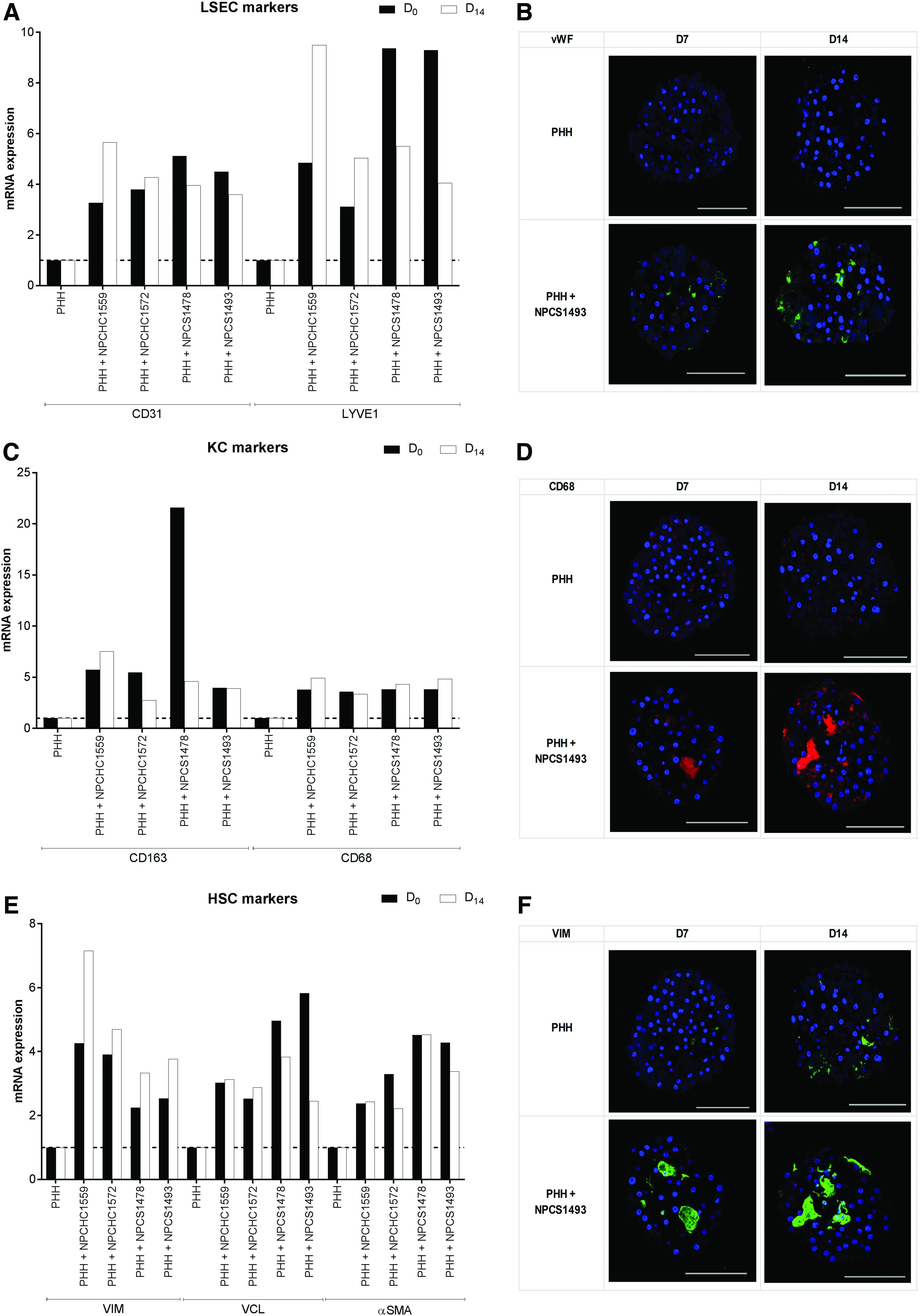
RT-qPCR and immunostaining of NPC markers in PHH/P1c-NPC 3D spheroid cocultures. mRNA expression of LSEC markers PECAM1/CD31 and LYVE1/LYVE1
Functions of PHH/NPC 3D spheroids
PHHs S1391T in monoculture showed a progressive decrease of ALB, APOB, and CYP3A4 mRNA expression as well as a decline in ATP and albumin secretion over time (data not shown), which is in line with the results obtained with other PHH monocultures (Fig. 2). These markers, however, remained in the range of 0.5–1 of D0 values up to 14 days postaggregation, thus being considered acceptable in terms of stability over duration of culture. 26 CYP3A4 was present up to D14 as revealed by immunostaining (Fig. 9). This figure also shows that ALB, APOB, and CYP3A4 gene expression were higher in PHHs S1391T cocultured with P1c-NPCs compared to PHHs S1391T in monoculture, in 3/4 cocultures from aggregation (D0) up to D14 postaggregation. Albumin secretion was also systematically higher in these three PHH/NPC cocultures compared to PHH cultures alone on D14 postaggregation. Figure 10A highlights that the five PHH/P1c-NPC cocultures tested, when stimulated with LPS, showed increased IL-6 secretion compared to PHH monocultures. In accordance with the increased IL-6 secretion induced by LPS in all tested cocultures, the cytotoxicity of trovafloxacin was significantly increased in LPS-stimulated cocultures compared with nonstimulated cocultures (Fig. 10B).
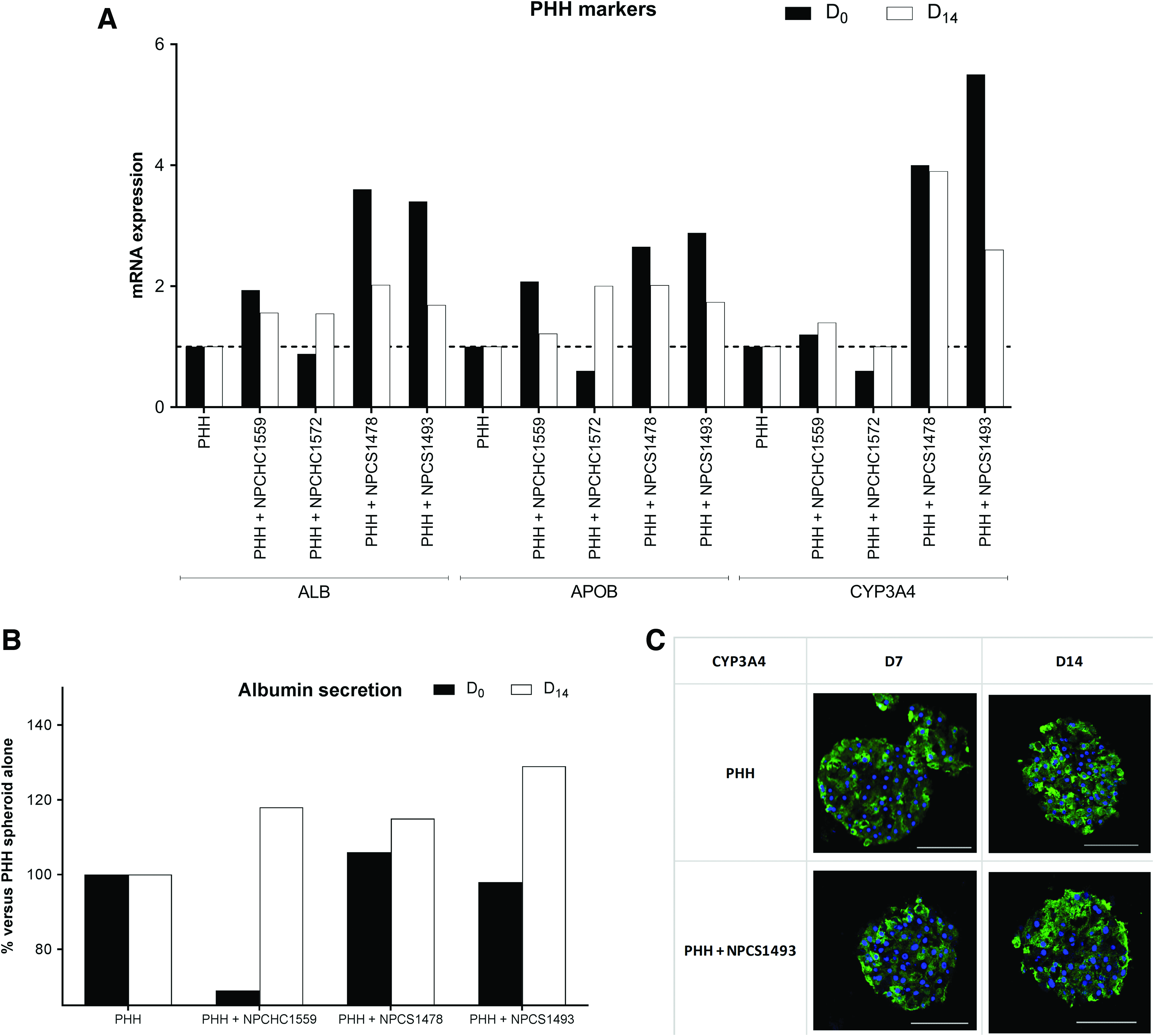
ALB/ALB, APOB/APOB, and CYP3A4/CYP3A4 mRNA expression
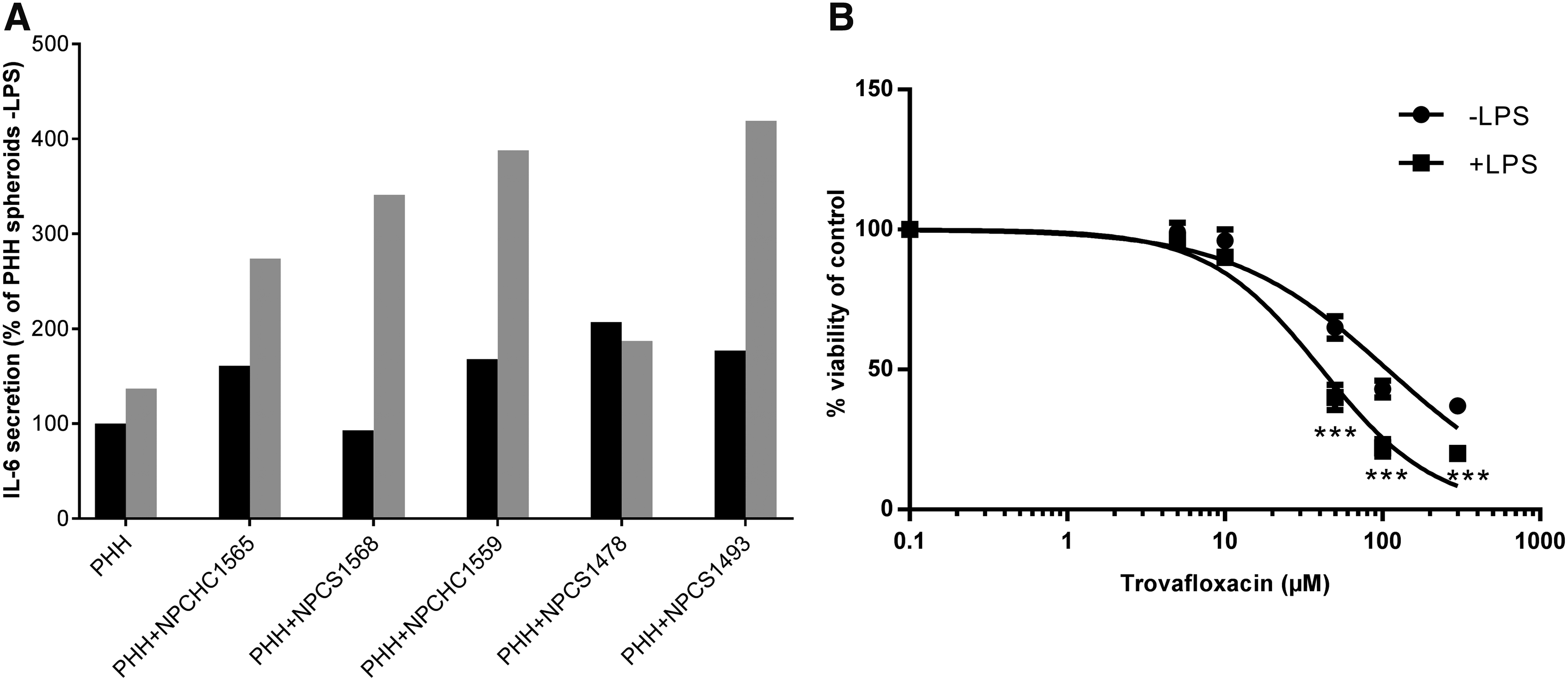
Functional characterization of KCs in 3D spheroids of PHHs/P1c-NPCs at aggregation.
Discussion
The scarcity in availability of human NPCs due to low yield of cells after isolation for human liver (∼2.5 × 10 6 NPCs/g liver and ∼20 × 10 6 PHHs/g liver according to the procedure of Werner et al. 20 and Kegel et al. 27 ) makes the use of 3D multicellular liver spheroids described here a particular interest for coculturing PHHs with NPCs, thanks to the low cell number (1330–2000 cells/3D spheroid) required. It thus allows the development of more integrated coculture model systems, with addition of LSECs, KCs, and HSCs to PHHs and indeed, some success in integration of either purified KCs 28 or crude NPCs7,9 together with PHHs into 3D spheroids have been reported.
Systematic evaluation and validation of these systems is limited and is necessary before such a platform can be more widely adopted for pharmacological and toxicological applications. In the present work we found that 79% (31/39) of cryopreserved plateable PHHs (with ≥80% confluency in 2D culture) successfully aggregate and form 3D spheroids when using ULA plates, while none of six nonplateable PHH lots aggregate. Once aggregated, 65% (20/31) of 3D spheroids were considered stable over the subsequent 14 days, using ATP maintenance as a benchmark. 7 On an average, size decreased about 8% and ATP about 36%. The reproducibility was, respectively, 96% for aggregation and 82% for stability over culture time. We thus confirm our recent results within the EU-funded MIP-DILI consortium that PHH lots prequalified for aggregation and stability in culture also successfully formed 3D spheroids when used by the other partners. 29 In the latter, we reported a good between-laboratory reproducibility in the cytotoxicity profiles of drugs with a given PHH–3D spheroid lot when independently generated using ULA technology. In this study, we show that size, morphology, stability of ATP content over culture time, and cytotoxicity profiles of PHHs (B1370T) were comparable in 3D spheroids obtained using ULA plates or applying the microtissue technology as described by Messner et al. 6
Hepatocytes account for about 60% of liver mass, whereas NPCs constitute ∼40% of the liver. 11 It was reported that coculturing PHHs with crude NPC fraction (2:1 ratio) as 3D spheroids results in interlot variability in the different NPC types incorporated. 28 It was, however, unclear if this was due to the absence of a given cell type in the NPC fraction or reduced/absence of incorporation efficiency into the 3D spheroid coculture. We report that all P1c-NPC fractions obtained using a low centrifugation step of the PHH supernatant fraction described by Werner et al. 20 (Supplementary Fig. S1) and followed by a culture step before harvesting and cryopreservation, maintain expression markers of LSECs, KCs, and HSCs at thawing. The reduction of PHH markers revealed only a minor contamination of PHHs, compared to P0c-NPC fractions immediately cryopreserved after isolation. This was confirmed after plating on plastic, by morphological observation and immunofluorescence revealed presence of protein markers of the three NPC types in P1c-NPCs, in accordance with our previous observations. 5
Although LSECs have a significant role in drug exposure and toxicity, their use in drug screening alongside hepatocytes is challenging, since they are not viable in classical 2D long-term culture. 30 In the present study, we found that at thawing, P1c-NPCs had higher expression of PECAM1/CD31 compared to P0c-NPCs. CD31, a widely used maker for endothelial cells, has been shown to be expressed both in normal and diseased liver, 31 but its increased expression has been associated with the development of a more vascular morphology, suggesting that this is occurring during the few days of culture before harvest and cryopreservation of P1c-NPCs. Expression of LYVE1, a newer marker of the endothelial phenotype, and a member of the link family of proteins recently described as scavenger receptors responsible for clearance of a variety of proteins by LSECs, 31 was barely present in P1c-NPCs, while highly expressed in P0c-NPCs. However, at aggregation, PHH/P1c-NPC 3D spheroids stably reexpressed this marker of functional LSECs, together with PECAM1, over the subsequent 14-day culture period. Immunostaining confirmed enrichment of vWF-positive cells in the PHH/P1c-NPC 3D spheroids up to D14.
KCs constitute the largest resident macrophage population in the body and are crucial for the regulation of immune-mediated hepatotoxicity and liver injury. 32 It is currently acknowledged that indirect hepatocellular injury caused by immune cell (KCs) activation and hepatic inflammation accounts for idiosyncratic drug-induced liver injury, 33 and that in vitro coculture systems are imperative to study these events. We report that after thawing, P1c-NPC fractions express markers of KCs, although the expression was lower than in P0c-NPC fractions. At aggregation, KCs were successfully integrated into the PHH/P1c-NPC 3D spheroids and remained present throughout the 14-day culture period as revealed by both mRNA expression and immunostaining of KC markers.
When activated, KCs release IL-6, 6 which was observed to be greater in most cocultures than the corresponding PHHs. It has been reported that HSCs also release inflammatory, proliferative, and fibrogenic cytokines such as IL-6 through direct contact with their neighboring cells. 34 We found that PHH/P1c-NPC 3D spheroids stimulated by LPS present increased IL-6 secretion and increased trovafloxacin toxicity compared with PHH 3D spheroids and suggest that PHH/P1c-NPC 3D spheroids represent an excellent model, in which to study drug-induced liver injury.
HSCs are pluripotent cells that are involved in the pathogenesis of liver fibrosis. Under physiological conditions, HSCs reside in the space of Disse, exhibiting a quiescent phenotype. Also, their main function is storing vitamin A and lipid droplets. It has been described that vinculin specifically stains human liver lobular HSCs. 35 There is some controversy about the pericellular matrix proteins such as αSMA and vimentin staining of HSCs in normal liver. Some reports indicate that at isolation, HSCs display vitamin A autofluorescence, with presence of numerous lipid droplets and absence of αSMA and vimentin staining, while others report staining of both vimentin and αSMA in HSCs in normal liver. 25 The results from the present study show that the crude P0c-NPC fraction isolated from human liver and immediately cryopreserved express VCL, VIM and ACTA1 genes. Expression of VCL and ACTA1 was lower, while that of VIM was higher in the P1c-NPC fraction. At aggregation, however, HSCs expressing VCL were successfully integrated into the PHH/P1c-NPC 3D spheroids and remained present up the 14 days culture period as revealed by the mRNA expression of all HSC markers. Immunostaining confirmed enrichment of vimentin-positive cells in the PHH/P1c-NPC 3D spheroids up to D14.
As a response to chronic injury, HSCs are activated and change from quiescent into myofibroblast-like cells, which are characterized by increased proliferation and high contractility with increased expression of pericellular matrix proteins αSMA and vimentin, and secretion of abundant extracellular matrix proteins (fibronectin, collagen type I and III). Such a phenomenon occurs during fibrosis, due to a continuous excessive production of extracellular matrix and collagen by HSCs, 36 which leads to permanent scarring and to organ malfunction that ultimately results in organ failure and death. Additional work is needed to assess if the HSCs incorporated in PHH/P1c-NPC 3D spheroids cocultures further respond to activation stimuli and could serve as a model for the investigation of pathophysiology of fibrosis, as has been recently reported in 3D bioprinted human liver tissues. 37
Hepatocytes, HSCs, and LSECs are the main cell types occupying the space of Disse in the liver. In vivo HSCs bridge between hepatocytes and LSECs. Such continuous hepatocyte–HSC–LSEC connections may lead to effective signal transfer among these cell types. Our results suggest that in PHH/P1c-NPC 3D spheroids, such connections are existing, with as a consequence, an increased functionality of PHHs, as revealed by increased mRNA expression of ALB, APOB, CYP3A4 proteins, as well as albumin secretion compared to PHH 3D spheroids. This result is in accordance with previous studies,30,38 suggesting that functionality of PHHs was better maintained in the presence of LSECs.
Conclusion
We report that 3D spheroid-qualified plateable PHH lots reproducibly form compact 3D spheroids, which maintain phenotypically stable in culture for at least 14 days. PHHs also aggregate into spheroids when mixed with a crude NPC fraction that prior to harvesting for cryopreservation had been in culture for a few days (PHHs/P1c-NPCs at 2:1 ratio). These 3D spheroid cocultures successfully and stably integrate LSECs, KCs, and HSCs and present increased PHHs functionality. The results of the present study suggest that this integrated coculture model system could present a valuable tool both for pharmacological and toxicological mechanistic studies as well as for the development of diseased liver models.
Footnotes
Acknowledgments
We thank M. Untrau (KaLy-Cell) and A. Fardellas (Karolinska Institutet) for technical assistance, InSphero for providing 3D spheroids in GravityTRAP™ plates with PHH lot B1370T, Orion, Karolinska Institutet, Janssen Pharmaceutical, GSK, and AstraZeneca for assessing cytotoxicity in PHH 3D spheroid monocultures and Hepacult for providing NPC HC1565, HC1567, HC1577, HC1559, and HC1572 batches. This work was supported by funding from European Community grants under the Innovative Medicines Initiative (IMI)—Programme (MIP-DILI Project No. 115336), the European Union Eurotransbio HepaCells ETB-2015-113 project, funded by the European Regional Development Funding (ERDF), “Investment strategy for growth and jobs” ALSACE 2014-2020 under grant agreement number AL0006907, and a grant from the Grand Est Region (France) and the Eurostars Programme (Cryosphere E! 11034).
Disclosure Statement
No competing financial interests exist. Magnus Ingelman-Sundberg is co-founder of HepaPredict AB.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.