
Abstract
Intrahepatic islet transplantation is a promising therapy for treatment of type 1 diabetes. During islet isolation, collagenase is used to extract islets from the pancreas, leading to loss of important cell–matrix interactions. Loss of the native pancreatic microenvironment is associated with apoptosis of islet cells, early graft failure, and poor islet function. The islet extracellular matrix (ECM) is composed of a specific combination of collagen (Col), laminin (LN), and fibronectin (FN) molecules. Reintroducing these molecules has been shown to boost the function, viability, morphology, and proliferation of β-cells. In this research, the effect of combinatorial ECM on islet function and survival was investigated. Specifically, thin-film microwell array scaffolds made from two distinct biomaterials were coated with FN, collagen type IV (Col4), LN111, LN332, or a combination thereof. We found that coatings containing a single type of ECM molecule, for example, FN or Col, can improve short-term islet function. However, these single proteins do not prevent loss of morphology and subsequent loss of islet function afterward. In contrast, combining Col4 with LN111 at a ratio of 8:2 not only improved short-term islet function but also preserved islet structure and islet function on a longer term. This effect was reproducibly shown on poly(ester-urethane) and poly(ethylene-glycol-terephthalate-poly(butylene-terephthalate) microwell islet delivery devices as well as tissue culture polystyrene. We concluded that biofunctionalization of inert biomaterials regardless of their molecular composition with a specific combination of islet ECM molecules can support and improve islet function over longer time periods. Our data suggested that creating a biomimetic islet niche by biofunctionalization of biomaterials can significantly improve the endocrine function of β-cells. The creation of islet mimicking niches in islet delivery devices leads to an improvement of islet function by restoring part of the islet's ECM in these devices.
Impact Statement
This research deals with finding a proper bioengineering strategy to improve the outcome of islets transplantation for treatment of type 1 diabetes. It is focused on the mimicking of islet extracellular matrix niche in microwell islet delivery devices to improve their endocrine function.
Introduction
Intrahepatic islet transplantation is a minimally invasive therapy to treat type 1 diabetes by replacing lost β-cells. This treatment results in insulin independence in 50% of the transplanted patients for a period of 3–5 years. 1 The success of islet transplantation is significantly influenced by the process of islet isolation from the pancreas. Current islet isolation methods result in separation of islets from their surrounding microenvironment by enzymatic digestion of the pancreatic extracellular matrix (ECM). Degradation of the ECM not only affects islet physiology but also negatively affects insulin gene expression, 2 insulin secretion, 3 proliferation, 4 differentiation,4,5 adhesion, and survival of β-cells.6–8 Therefore, degradation or removal of islets ECM molecules such as collagen (Col), fibronectin (FN), and laminin (LN) during islet isolation can reduce the efficacy of islet transplantation by providing a suboptimal β-cell microenvironment.
To overcome problems associated with the removal of ECM molecules after islet isolation, several researchers have investigated the addition of a new ECM to isolated islets. Results of these studies revealed the beneficial short-term effects of a new ECM on islet function. For instance, the mechanical support and topological arrangement of the islet's endocrine cells were improved in the presence of certain ECM molecules. Consequently, insulin secretion and islet survival were enhanced in these studies. 9 More specifically, it was reported that a combination of collagen type I (Col1) and collagen type IV (Col4) improves the insulin secretion of β-cell. 10 In another study, it was shown that culturing islets in Col1 hydrogels mixed with Col3, Col4, and LN improves the survival of the islets. 11 The ECM molecules secreted by 804G cells increase the insulin release of β-cells. 12 The specific sequence of LN511 promotes islet viability as well as insulin release. Moreover, the synergistic beneficial effect of the leu-arg-glu sequence of LN in combination with Col4 was reported. 13 In addition, some studies focused on the interaction of ECM molecules with integrin and nonintegrin receptors that are expressed by human islets. Such interactions regulate adhesion, differentiation, insulin secretion, and survival of β-cells. For instance, human β-cell adhesion, motility, and insulin secretion were associated with the interaction of Col4 and its specific integrin receptors (α1β1). 3 The β-cells are also known to bind arginine–glycin–aspartic acid (RGD) sequences present in ECM molecules by expressing integrin α3β1, α5β1, and αvβ3 and thereby promoting cell survival. 7 Therefore, it is highly important to increase knowledge about the response of human islet cells upon their interaction with ECM molecules.
Previous research has mainly focused on the effect of specific individual ECM proteins on the function and survival of islet cells. However, studies on the combinatorial effects of different ECM molecules at different ratios on islets are lacking. Considering that the islet microenvironment is a complex system and the success of islet transplantation relies on better simulation of the islet microenvironment, a profound investigation of this complexity is necessary. It is important to discover the most effective combination of ECM molecules in a topologically similar microenvironment.
Here, we report about thin-film microwell array scaffolds using a poly(ethylene-glycol-terephthalate-poly(butylene-terephthalate) (4000PEOT30PBT70) abbreviated as (PEOT-PBT) and a poly(ester-urethane) to deliver islets. The poly(ester-urethane) was synthesized by reaction of an α,ω-isocyanate end-functionalized prepolyester and an α,ω-hydroxy biscarbamate. The scaffolds were coated with combinations of FN, Col4, as well as various LNs, and afterward, the glucose responsiveness of human islet inside these devices was investigated. We showed that a combination of laminin-111 (LN111) and Col4 has a reproducible synergistic positive effect on the function of the human islets. Furthermore, the shape and dimensions of the microwell scaffolds used offer a potential advantage to support the three-dimensional (3D) structure of islets. Interestingly, the positive effect on endocrine function was independent of the biomaterial used to manufacture the microwell scaffold, and not only the amount of insulin secreted by the islets was significantly increased but also their stimulation index was higher in the presence of Col4 and LN in a ratio of 8:2.
Materials and Methods
Fabrication of PEOT-PBT and poly(ester-urethane) films
The PEOT-PBT block copolymer is composed of a soft hydrophilic segment poly(ethylene-glycol-terephthalate) and a hard hydrophobic segment poly(butylene-terephthalate) and was produced by PolyVation BV (Groningen, The Netherlands, under the trade name PolyActive™). PEOT is synthesized using poly(ethylene-glycol) with a molecular weight of 4000, whereas the weight percentage of PEOT and PBT is kept at 30:70 wt%, respectively. 14 Films with a thickness of 40 μm were created using a solvent casting method. Shortly, PEOT-PBT was dissolved in a 65:35 (w/w) mixture of chloroform (Merck, Darmstadt, Germany) and 1,1,1,3,3,3-hexafluoro-2-isopropanol (Biosolve, Valkenswaard, The Netherlands) at a concentration of 15 wt%. After casting the polymer solution on a silicon wafer at room temperature, a nitrogen flow was applied for 4 h to dry the resulting film. Residual solvent was removed by immersing the polymer film in ethanol. Complete evaporation of residual solvents was achieved by placing the films in a vacuum oven (Heraeus, Hanau, Germany) at 30°C for 3 days. The same protocol was applied to fabricate poly(ester-urethane) (Polyganics BV, Groningen, The Netherlands) films with a thickness of 40 μm from a 5 wt% polymer solution in chloroform. The poly(ester-urethane) was synthesized by reaction of an α,ω-isocyanate end-functionalized prepolyester and an α,ω-hydroxy biscarbamate.
Preparation of PEOT-PBT and poly(ester-urethane) microwell scaffolds
Micro thermoforming was applied to manufacture microwell scaffolds with a 3D microwell structure as was reported before. 15 The backing film materials used for micro thermoforming of PEOT-PBT and poly(ester-urethane) microwell scaffolds consisted of either polyethylene (560 μm thickness) or paraffin (540 μm thickness) placed on top of a 20-μm-thick polyethylene film, respectively. The films were placed on stainless steel mold created by laser micromachining (Lightmotif BV, Enschede, The Netherlands). For the poly(ester-urethane) microwell scaffold, the initial mold pressure was set at 30 kN at 25°C, after which the temperature was gradually increased over time. During this process, the pressure was stepwise increased at 30°C, 33°C, and 41°C to 40, 50, and 60 kN, respectively. The pressure was released as soon as the temperature reached 41°C, followed by immersion in ice-cold ethanol for 15 min. Subsequently, the films were gently peeled off their molds. Any residual traces on the poly(ester-urethane) scaffolds of paraffin were removed by immersing the microwells in xylene for 1 min followed by five times rinsing with 70% ethanol and then water. Fabrication of PEOT-PBT microwell scaffolds was performed in a slightly different way. In short, the mold pressure was set at 40 kN at 30°C. The pressure was kept constant at 40 kN, whereas the temperature was increased to 85°C. At 85°C, the pressure was increased to 65 kN, and directly thereafter, the temperature was set to 20°C. As soon as the press device was cooled to 20°C, the pressure was released. Microwell films were then placed in 70% ethanol for 15 min at room temperature before removing the scaffold from the mold. Before seeding the islets, the microwell scaffolds were disinfected by immersion in 70% ethanol overnight. The dimensions of individual microwells in the PEOT-PBT and poly(ester-urethane) films were determined by measuring 10 random microwells of every third polymer scaffold manufactured.
Immobilization of ECM molecules on culture substrates
The thin-film PEOT-PBT and poly(ester-urethane) microwell array devices were shortly exposed to oxygen plasma at an oxygen pressure of 1.0 bar, at 40 mA, and 200 mTorr for 10 and 20 s, respectively. The oxygen plasma treatment was used to modify the surface chemistry of the microwell scaffold and generates, for example, aldehyde groups that can react with molecules containing free amine groups present in the ECM proteins. Subsequently, microwell scaffolds and tissue culture polystyrene plates were coated with 10 μg/mL of ECM proteins. The ECM proteins include FN, Col4, LN111 (Merck), LN332 (Biolamina, Sweden), and a mixture of LN111, 211, 121, 311, 411, and 511 (Merck). With a total protein concentration of 10 μg/mL, the pairwise compositions of varying ratios of FN, Col4, and LN (5:5, 2:8, and 8:2) were coated at room temperature for 2 h. After three times rinsing in phosphate-buffered saline (PBS) to remove any unbound proteins, the scaffolds were placed in cell culture inserts and transferred to ultra-low-attachment cell culture plates (Corning).
Primary human islet cultures
Human islets of Langerhans were isolated from donor pancreas at the Human Islet Isolation Laboratory, Leiden University Medical Center, Leiden, The Netherlands. Human donor islets were only used if not deemed suitable for clinical islet transplantation and if research consent was present according to national laws and regulations.
Isolated human donor islets were cultured in CMRL-1066 culture medium (Cellgro, Mediatech, VA) containing 5.5 mM
Human islets from two donors (a 49-year-old man and a 75-year-old woman) were carefully handpicked, and then, 50 islets were seeded per microwell scaffold (an islet per well), which was located in tissue culture plate (n = 3). Medium was refreshed every other day. Human islets were seeded on nonadherent polystyrene plates, PEOT-PBT, and poly(ester-urethane) microwell scaffolds coated with ECM proteins. Several controls were prepared by seeding islets on ultra-low adhesive plates (free-floating control), bovine serum albumin (BSA)-coated tissue culture plates, and noncoated and BSA-coated microwell scaffolds (nonrelevant protein controls) made from both individual polymers.
Immunofluorescence staining
The morphology of islets was evaluated by histological analysis and microscopy. Following fixation in 4% (w/v) paraformaldehyde and permeabilization by 0.3% Triton for 3 h, human islets were immunofluorescent labeled for glucagon and insulin. 16 The islets were blocked with 10% normal goat serum/0.15% Triton-X 100/10 mM PBS overnight at 4°C. The primary and secondary antibodies were diluted in 1% BSA/0.2% Triton X-100/10 mM PBS. Staining for glucagon was performed using primary rabbit anti-glucagon (1:100; Abcam, The Netherlands), and two different secondary antibodies including biotin anti-rabbit (1:200; Abcam) and streptavidin-Alexa Fluor 488 (1:200; Abcam) to distinguish alpha from β-cell. In case of insulin-positive β-cells, guinea pig anti-insulin (1:200; Abcam) was used as the primary antibody, whereas Alexa Fluor 647 goat anti-guinea pig (1:500; Abcam) was applied as the secondary conjugated antibody. Nuclei were stained using 4′,6′-diamidino-2-phenylindole dihydrochloride (DAPI) (Abcam) for 10 min. Each step was followed by subsequent washing in 1% BSA/0.2% Triton X-100 in 10 mM PBS. Samples were subjected to optical sectioning in axial (z) dimension using a Zeiss LSM510 confocal laser scanning microscope.
Glucose-stimulated insulin secretion tests
Glucose-stimulated insulin secretion (GSIS) tests were performed using triplicate samples of 50 human islets for each condition at 3 and 7 days after seeding. Each sample was first incubated for 1 h in a glucose-free incubation buffer, in a modified Krebs-Ringer bicarbonate buffer with HEPES (KRBH) containing 115 mmol/L NaCl, 5 mmol/L KCL, 24 mmol/L NaHCO3, 2.2 mmol/L CaCl2, 1 mmol/L MgCl2, 20 mmol/L HEPES, and 2 g/L human serum albumin (Sanquin, The Netherlands), pH 7.4. Samples were then exposed to low glucose (1.6 mmol/L), high glucose (16.7 mmol/L), and another low glucose (1.6 mmol/L) KRBH buffer each for 90 min at 37°C. After each incubation step, samples' media were collected for insulin concentration measurements by ELISA (Mercodia, Uppsala, Sweden). Finally, the islets from each sample were collected for DNA quantification using a Quant-iT Picogreen dsDNA assay (Invitrogen). Amount of insulin release was measured after low and high glucose stimulation and normalized to the DNA of islets. Moreover, the stimulation index was calculated as a ratio of insulin value after high glucose stimulation divided by the insulin value after low glucose stimulation.
Statistical analysis
Results are presented as the mean ± standard deviation. Statistical analysis was performed using one-way ANOVA, the least significant difference multiple comparison test, and Tukey's post hoc test by the application of SPSS statistic software (Chicago). Statistical significance was considered at a p-value of ≤0.05. The statistical analysis was performed to compare the glucose responsiveness of human donor islets in high glucose (16.7 mmol/L), between control groups (noncoated and BSA coated) versus functionalized culture-substrate, PEOT-PBT, and poly(ester-urethane) microwell scaffolds at day 3 and day 7. In addition, the statistical difference between control groups was calculated.
Results
Microwell scaffold morphology
The poly(ester-urethane) and PEOT-PBT microwell scaffolds were fabricated using micro thermoforming, which is an adapted method14,15 (Fig. 1A). Scanning electron microscopy analysis indicated similarly shaped microwells mask-aligned across the surface (Fig. 1). The process was highly accurate and reproducible, and all microwell scaffolds remained stable during handling. PEOT-PBT microwells had a circular and tapered appearance with aperture diameter of 380 ± 5 μm and bottom diameter of 300 ± 5 μm, and the distance between the aperture and base was 300 ± 10 μm. Poly(ester-urethane) microwells had a similar aperture diameter, but a different base diameter of 250 ± 5 μm.

The structure of microwell scaffolds.
Function of primary human islets
The effect on glucose responsiveness of human donor islets of the culture-substrate (nontreated tissue culture plastic), PEOT-PBT, poly(ester-urethane), and flat films versus microwells islets functionality was studied at 3 and 7 days. Islets cultured on PEOT-PBT and poly(ester-urethane) microwell scaffolds released significantly more insulin compared with free-floating controls (Fig. 2). Moreover, the increase in insulin secretion showed a consistent, yet discrete, tendency to be highest in islets cultured on PEOT-PBT groups. In addition, islets seeded in microwells secreted more insulin than islets seeded on flat membranes of the same materials. Furthermore, while insulin release of islets on a flat surface declined over time, insulin secretion of islets cultured in microwells increased over time.

Glucose-stimulated insulin secretion test of human islets on ultra-low adhesive plate (control), PEOT-PBT, and poly(ester-urethane) polymer film and microwell scaffold at day 3 and day 7. Values represent mean ± SEM. *Indicates statistically significant differences (p < 0.05) for the glucose responsiveness of islets in high glucose when compared with free-floating controls at day 3 and day 7. SEM, standard error of the mean.
The PEOT-PBT and poly(ester-urethane) microwell scaffolds were functionalized with FN, Col4, LN111, or LN332. Additionally, these ECM molecules with a total concentration of 10 μg/mL were pairwise combined at a mass ratio of 5:5, 2:8, and 8:2. After seeding the human islets into the biofunctionalized microwells, the effect of the different ECM molecules on insulin secretion at day 3 and day 7 were assessed and values were normalized to the amount of insulin secreted per DNA. The results were compared with control groups consisting of human islets cultured on nontreated and BSA-functionalized PEOT-PBT and poly(ester-urethane) microwell scaffolds. Human islets significantly increased their glucose secretion level at day 3 when cultured on biofunctionalized tissue culture polystyrene surfaces with FN, Col4, LN332, Col4:LN111 (5:5, 2:8, and 8:2), and FN: LN111 (8:2) (Fig. 3). Similar results were observed for islets from the same donors seeded into the ECM-functionalized microwell scaffolds. We observed that in general the amount of insulin secreted by islets seeded on ECM protein-functionalized PEOT-PBT scaffolds was higher than that on poly(ester-urethane) scaffolds, and significantly higher compared with islets seeded on standard tissue culture polystyrene (Fig. 4). We observed that insulin secretion in the two microwell scaffold groups (Figs. 3 and 4) in the presence of FN and Col4 shows significantly increased insulin secretion compared with their control groups (noncoated and BSA coated) on both day 3 and day 7 of culture. The insulin secretion from islets seeded on PEOT-PBT and poly(ester-urethane) microwell scaffolds, and tissue culture polystyrene coated with LN111 was lower than their control groups (noncoated and BSA coated) on day 3. However, insulin secretion increased in islets cultured in microwell scaffolds after 7 days, whereas in the tissue culture polystyrene group, no change was observed. Islets cultured on LN332-coated scaffolds show an enhanced insulin secretion only after 3 days but not after 7 days. Comparison of insulin secretion of islets cultured on Col4 at day 7 indicates a significant better glucose response in the two microwell scaffold groups, then the polystyrene controls.
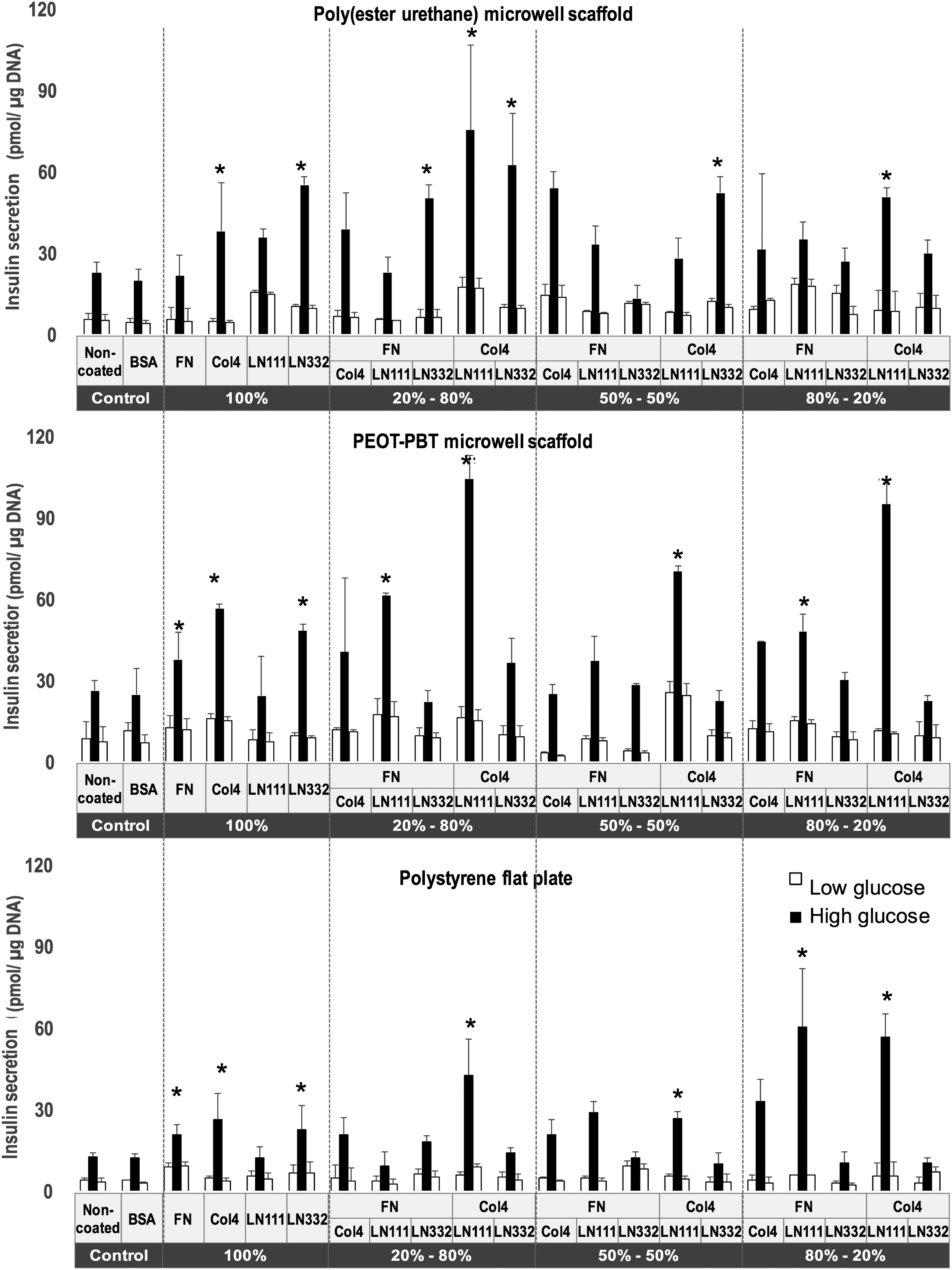
Glucose-stimulated insulin secretion test of human islets on monolayer-coated ECM proteins on poly(ester-urethane) microwell films (top panel), PEOT-PBT microwell films (middle panel), and polystyrene tissue culture plastic (bottom panel) with different ratios of ECM proteins at day 3. Values represent mean ± SEM. *Indicates statistically significant differences (p < 0.05) for the glucose responsiveness of islets in high glucose when compared with the controls (nontreated and BSA-coated controls) at day 3. BSA, bovine serum albumin; ECM, extracellular matrix.
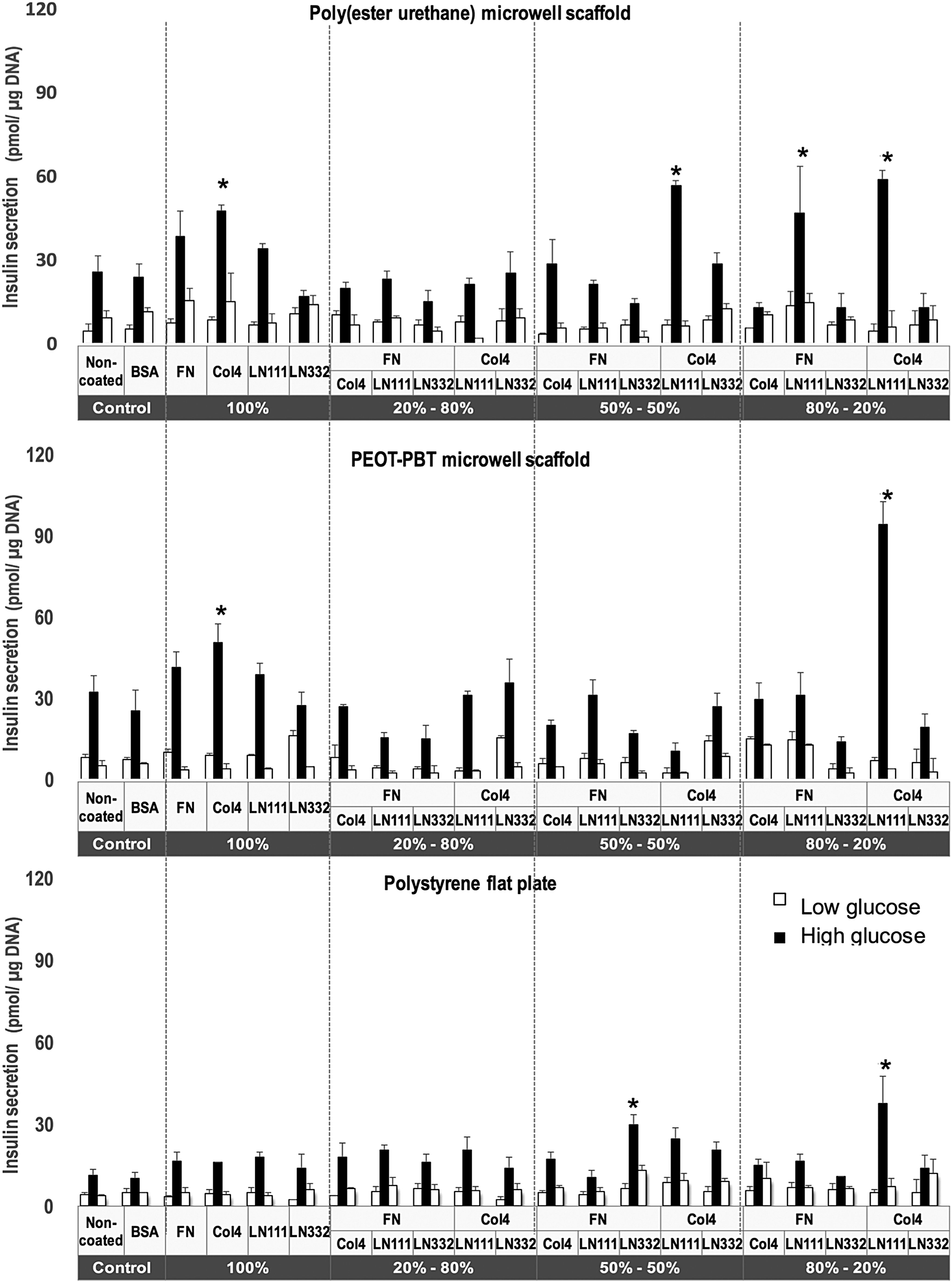
Glucose-stimulated insulin secretion test of human islets on monolayer-coated ECM proteins on poly(ester-urethane) microwell films (top panel), PEOT-PBT microwell films (middle panel), and polystyrene tissue culture plastic (bottom panel) with different ratios of ECM proteins at day 7. Values represent mean ± SEM. *Indicates statistically significant differences (p < 0.05) for the glucose responsiveness of islets in high glucose when compared with the controls (nontreated and BSA-coated controls) at day 7.
Interestingly, a significant increase in insulin secretion was observed for Col4- and LN111-coated (mass ratios used 5:5 and 8:2) scaffolds at day 3. Although at 7 days a slight decrease in insulin secretion was observed in most conditions, islets cultured on Col4:LN111-coated (8:2) microwell scaffolds showed a clear increase of insulin secretion (Figs. 3 and 4). The outcomes from islets cultured on a combination of Col4- and LN111-functionalized scaffolds show consistently that their endocrine function has significantly improved in the presence of these specific ECM proteins in comparison to all other conditions.
By comparing the islet function (see Supplementary Figures S1 and S2) in nonfunctionalized microwell scaffolds with tissue culture polystyrene, the positive influence of the 3D microwell shape on the islet's endocrine function can be analyzed. In both poly(ester-urethane) and PEOT-PBT microwell scaffolds, glucose responsiveness of islets increased between day 3 and day 7, whereas on tissue culture polystyrene, a slight decrease of insulin release was observed. In short, islets on flat polystyrene plate as well as microwell groups functionalized with FN, Col4, LN332, and a combination of Col4 and LN111 (5:5, 2:8, and 8:2), FN and LN111 (8:2), and FN and Col4 (8:2) indicated significantly increase of glucose secretion at day 3 in comparison to their control groups (nontreated and BSA coated) at day 3. Comparing the insulin release at day 3 and day 7 indicated the reduction in the rate of insulin release over time (from day 3 to day 7) in the presence of aforementioned ECM molecules. However, stimulation index for the flat polystyrene and PEOT-PBT and poly(ester-urethane) microwells coated with a combination of Col4:LN111 (8:2) was significantly higher than nonfunctionalized groups at both day 3 and day 7. It indicated the importance of this combination of Col4:LN111 (8:2) for the function of islets over a longer time span.
Islet structure on the ECM-coated surfaces
To assess whether the presence of ECM molecules on biomaterials can affect islet morphology, human islets were seeded on flat surface polystyrene plates functionalized with ECM molecules. The ECM proteins include FN, Col4, LN111, LN332, a mixture of LN111, 211, 121, 311, 411, and 511, as well as pairwise combinations of FN, Col4, and LNs with a total protein concentration of 10 μg/mL at mass ratios of 5:5, 2:8, and 8:2. Additionally, ultra-low adhesive plates were used as free-floating controls. Islets were stained for insulin and glucagon using immunofluorescent labeling and imaged using phase-contrast transmission light and confocal fluorescent microscopy.
We observed that the islets in the free-floating control group did not adhere to the surface of nontreated plates and thus they kept their native round shape until day 7 (99% ± 0.8% of islets at day 3 and 90% ± 2.1% at day 7) (Fig. 5a, g). However, histological observations revealed that in these islets it seems that there are relatively less insulin-positive β-cells compared with the other conditions (Fig. 5m–r).

Morphology of islets on polystyrene that was coated with FN, Col4, LN111, LN332, and a combination of Col4 and LN111 at a ratio of 8:2 at day 3 and day 7. The islets cultured on ultra-low adhesive plates were used as a control group.
Human islets cultured on FN- and Col4-functionalized substrates adhered to the surface and showed cellular outgrowth after 7 days resulting in loss of their rounded morphology (100% of islets) (Fig. 5h, i). In contrast, human islets cultured on LN332-functionalized substrates had only partially lost their round morphology (Fig. 5e, k). The same islets cultured on substrates with coatings of Col4 and LN111 at 5:5 and 8:2 mass ratios showed some adherence to the underlying substrate, thereby fixing the islets in place, while their original rounded morphology was preserved (91% ± 1.7% of islets) (Fig. 5f, i). Moreover, histology revealed that these islets seemed to contain a higher numbers of functional β-cells after 7 days in comparison to the other conditions (Fig. 5m–r).
Discussion
Islet transplantation is an efficient treatment for type 1 diabetes. However, it faces many obstacles such as reduced function of islets or loss of 60% of the islets in a short period of time after transplantation.17,18 In clinical islet transplantation, the islet ECM is degraded by the enzymes used during the isolation procedure. 19 It was also reported that the β-cells are not the most important producers of ECM in islets, but instead capillary endothelial cells were found to be the dominant producers of basement membrane (BM), which forms a major part of the islet's ECM. 20 Therefore, it is critical to support transplanted islets with external ECM molecules, particularly in the first 2 weeks, until revascularization and deposition of new ECM can take place. 21 The aim of this study was to investigate the effect of specific islet ECM molecules on the endocrine function and cell behavior in human donor islets. Human islets from two different donors were used for performing the experiments because they have karyotype and function similar to in vivo situation. However, using different donors might result in donor-dependent variation in the level of insulin secretion. The selection of specific ECM molecules in this study was based on what is currently described in the literature describing their presence in the microenvironment and possible influence on the physiology of pancreatic islet cells. It is known that the signaling interaction between the ECM molecules and islet cells can regulate multiple aspects of islets physiology, such as insulin release, survival, and proliferation of β-cells. 22 Naba et al. by performing quantitative proteomic profile of the ECM of normal pancreatic islets detected 120 ECM and ECM-associated proteins, which are composed of 57 ECM glycoproteins, 37 collagens, 9 proteoglycans, 23 ECM-affiliated proteins, 42 ECM regulators, and 10 secreted factors. Among these molecules, fibrillar collagens, FN, LNs, and nidogens are the most abundant molecules of the islet ECM. 23 We selected FN, Col4, LN111, and LN332, which were all shown to support the function and viability of islets in previous publications.3,24,25
Not only the effect of individual proteins but also the combinatorial effect of these proteins on islets was studied. Furthermore, we compared the effect of these proteins when bound to the surface of poly(ester-urethane) or PEOT-PBT microwell scaffolds on the glucose responsiveness of human donor islets with their nonfunctionalized counterparts.
One of the drawbacks of most macroencapsulated islet delivery devices is the clustering and aggregation of islets inside these devices. Aggregation negatively affects islet morphology and function by limiting the oxygen and nutrients supply leading to necrosis in the core of islets.17,26,27 We showed that in this and previous studies that by seeding islets in individual microwells, the islet's rounded morphology can be preserved and aggregation prevented.14,16 In general, we observed in this study that islets cultured in microwell scaffolds show a higher insulin secretion in comparison to the same islets grown on flat polymer substrates. This change in islets function and morphology seems to be related to the design of the microwell scaffold's semispherical structure. In addition, each microwell acts as a physical barrier to avoid aggregation of neighboring islets. The mechanical properties of the polymer scaffolds also might have a positive influence on islets. The tissue culture polystyrene plates have a higher stiffness than both poly(ester-urethane) and PEOT-PBT, which are rather elastic polymers in part explaining the lower insulin secretion levels found in these controls. Seewaldt observed that the stiffness of ECM can induce physical changes in breast cancer cells and influence their behavior due to a phenomenon called mechanotransduction. 28 To the best of our knowledge, there has not been systematic study on the effect of ECM stiffness and elasticity in relation to β-cell function.
Regarding the influence of ECM molecules on islets, the GSIS test indicated significant increase of insulin secretion for the islets that were seeded on substrates functionalized with either FN or Col4. The positive impact of Col4 on the function of islets in in vivo 29 and in vitro 30 was reported previously. Moreover, it was reported that the cultivation of islets in the presence of FN increases insulin release and preserves the viability and mass of islets after transplantation. 24 Interestingly, although FN and Col4 molecules individually display a positive effect on islets, based on our findings the combinations of these two molecules have a negative effect on insulin secretion. In favor of this finding, Flaim et al. reported similar results regarding the function of hepatocyte cells. 31 They assessed the effect of Col4 and FN on cellular differentiation of primary rat hepatocytes by immunofluorescent staining of intracellular albumin (a marker of specific liver function). Kaido et al. have described that there is a strong interaction of islets with Col and FN, which can result in a transition from a rounded morphology into a monolayer-like appearance. 29 It is reported that the disruption of the spheroidal architecture of the islets can possibly interfere with integrin-mediated signaling and paracrine interactions between individual islet cells. 32 The integrin-mediated signaling in response to the pancreatic ECM plays a crucial role in β-cell survival and insulin secretion. For instance, integrin binding to Col including α1β1, α2β1, and α3β1 integrins regulate cell adhesion, insulin gene expression, and islet survival.
Korpos et al. reported that LNs of peri-islet BM (consisting of LN α2, α4, β1, β2, γ1, γ2, and γ3 chains) are distinct from the LNs found in close vicinity of surrounding blood vessels (consisting LN α5, β1, and γ1 chains). 33 Since, LN β1 and γ1 chains are common between peri-islet ECM and the BM of surrounding blood vessels, we expected to see significant influence of LN111 on islets. Interestingly, the interaction of LN111 with islets did not positively affect insulin secretion, whereas LN111 in combination with Col4 showed a significant increase in the insulin secretion even higher than Col4 alone. It is reported that Col4 tend to bind via the intermediary glycoproteins nidogen, or entactin, or via direct interaction with heparin sulfate proteoglycans. 34 Our results suggested that a combination of important ECM proteins can play a synergistic role to enhance insulin secretion of human islets. Comparing all different combinations used in this study, we found the highest level of insulin secretion in human islets cultured on a combination of Col4 and LN111 at a mass ratio of 8:2 with a concentration of >5 μg/cm2 after 7 days. Histological observations revealed that islets seeded on both microwell scaffolds and flat substrates coated with Col4:LN111 (8:2) maintain their rounded shape and thus improve the function of islets even further. In addition, the semispherical structure of the microwells in the scaffold exposes the islets to an ECM-functionalized substrate almost all around their exterior, whereas in the flat substrate, the islets are only exposed to ECM at the bottom. Therefore, based on our data, we postulate that the combination of two ECM molecules (Col4 and LN111 with 8:2 mass ratio) along with specific design of microwell scaffolds could have an additional positive effect on the endocrine function possibly by a more efficient exposure to the biofunctionalized surface. But one important question still remains: Why does the insulin secretion increase so significantly if only a few cells on the exterior of islets are exposed to a biofunctionalized substrate? We think that there might be a few plausible explanations for our findings.
It is commonly accepted that insulin secretion is mediated via Ca2+; when β-cell secretes insulin, this is accompanied with a sudden raise in intracellular Ca2+ levels via an influx through specific voltage-gated channels in the cell membrane or via the release of endoplasmic reticulum-stored Ca2+ into the cytosol. Although the activation of specific signal transduction pathways triggered by integrin binding is not very well studied in β-cells, there is quite some evidence from other fields that show that integrin binding can lead to the activation of several important pathways such as MAPKs and Rho via integrin-regulated FAK/Src signaling. 35 Both pathways involve an increase in cytosolic Ca2+, perhaps this increase could lead to enhanced glucose responsiveness. It is known that Ca2+ is involved in many different mechanisms in β-cells, varying from mitochondrial function to β-cell electrical activity. 36 No relationship between specific integrin binding and cytosolic Ca2+ has been reported so far; therefore, we can only speculate about its role in improved insulin secretion and this only in part explains our findings. Another possible explanation could be that β-cells can act as β-cell hubs, which can drive insulin secretion in pancreatic islets. Work performed by Rutter and colleagues suggests that certain β-cells can act as pacemakers for insulin secretion in islets. They found that a small group of β-cells are leading rhythmic pulsatile insulin secretion and connect to many other β-cells in islets. 37 Their finding can perhaps explain why even though only a handful of β-cells located on the outside, which are in direct contact with ECM proteins, can enhance insulin secretion so significantly in our scaffolds.
In conclusion, we indicated that islet–ECM interactions can influence the insulin secretion of human donor islets. Understanding the role of each particular ECM molecule on insulin secretion, viability and morphology of the islets are key factors to engineer biomimetic microenvironments of β-cell replacement devices to treat type 1 diabetes. We showed that exposure of islets to LN111 (>5 μg/mL) in combination with other ECM proteins (<5 μg/mL) results in a decline in insulin secretion over time. In contrast, lower ratios of LN (<5 μg/mL) in combination with other ECM molecules led to a sustained release of insulin over a 7-day period. Both PEOT-PBT and poly(ester-urethane) microwell scaffolds functionalized with the combination of Col4 and LN111 with a mass ratio of 8:2 led to significantly enhanced insulin secretion in human donor islets.
Footnotes
Acknowledgments
This research was financially supported by the Diabetes Cell Therapy Initiative (DCTI) FES 2009 program LSH-DCTI including the Dutch Diabetes Research Foundation (DF). Dr. J.L. acknowledges financial support from the Dutch NWO innovative research incentives scheme Veni award (#14328) and the European Research Council (ERC, Starting Grant #759425).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.