
Abstract
Since it was first introduced in 2011, three-dimensional (3D) “Sasai” method for retinal differentiation became a strategy of choice for retinal tissue and neuron production. It is based on the recapitulation of retinal development and requires several stages: aggregate formation, neuroectoderm induction, and eye field induction, followed by retinal maturation. To achieve the consistency of retinal differentiation needed for drug discovery and cell transplantation, we have attempted to improve spheroid formation as well as approach xeno-free conditions. In this study we compared the effect of cell culture plate shape and material, medium viscosity, lipid, and bovine serum albumin concentrations on aggregate formation from mouse embryonic stem (mES) cells. We have also assessed the possibility of substituting Matrigel with the synthetic vitronectin-mimicking oligopeptide. RX-GFP mES cell line was used for experiments. The dose–response of synthetic extracellular matrix (ECM) has been assessed and quantified by live fluorescence microscopy, immunohistochemistry, flow cytometry, and quantitative polymerase-chain reaction (qPCR) for early retinal development genes (Rx, Pax6, Lhx2, Sox2, Six6). The comparison of seeding conditions at 24 h postseeding showed the dose-dependent effects of lipids (lipid concentration of 2% resulted in 100% efficiency of aggregate formation and significant increase in size to 532.8 ± 31.87 μm, p < 0.05); and viscosity (methylcellulose concentration of 0.06% in optic vesicle medium showed 100% efficiency and increase in aggregate size 532 ± 19.23 μm, p < 0.01). The addition of synthetic matrix resulted in retinal differentiation (34.47% of RX as detected by flow cytometry compared with 33.8%, observed with Matrigel). The early retinal genes expression at day 7 was confirmed by qPCR. We present the optimized conditions for 3D retinal differentiation, including the option of xeno-free ECM. These defined medium conditions significantly decrease the variability within and between batches and allow substantial scale up of retinal tissue and cell production for drug discovery, disease modeling, and transplantation purposes.
Impact Statement
The development of retinal regenerative therapies relies on the reproducible and renewable source of retinal neurons for drug discovery and cell transplantation. Three-dimensional approach for retinal differentiation from pluripotent cells recently emerged as the robust strategy for retinal tissue differentiation. In this work, we present the combination of optimized conditions and techniques for three-dimensional retinal differentiation from mouse embryonic cells that improves reproducibility and efficiency of retinal differentiation in organoid cultures. We also show that the retinal induction can be achieved with the synthetic oligopeptide instead of Matrigel that allows to approach xeno-free conditions for cell production.
Introduction
Retinal neuron loss of various etiology remains the leading cause of blindness in developed world with an immense socioeconomic impact.1,2 The death of photoreceptors in age-related macular degeneration and retinal ganglion cells in glaucoma is complicated by the lack of regenerative capacity in the adult mammalian retina. Several approaches, including gene therapy, neuroprotection with growth factors, small molecules and cell transplantation, as well as direct cell replacement are currently being explored. The primary cells and retinal explants, isolated from adult or developing animal and/or human eye have been used for disease modeling, in proof-of-concept studies of cell transplantation, to identify neuroprotective drugs and to test the specificity and efficiency of gene therapy.
Since the cultures of mouse and human embryonic stem cells (hESCs) have been established in vitro, the idea of using this reproducible source for retinal neuron differentiation has been widely investigated. For two decades, monolayer (two-dimensional, 2D) culture method remained the major approach for pluripotent cell differentiation toward retinal fate. Adherent cultures, although, mainly yield single-cell-type populations (retinal pigment epithelium [RPE] or retinal ganglion cell) by its selective expansion and passaging.3,4 Evolving 2D method, intermediate 2D/three-dimensional (3D) protocol takes in spontaneous spheroid formation from stem cell colonies and subsequent spheroid culture on adhesive substrate.5–7 The reported efficiency of these techniques is 20% Crx+ photoreceptor precursors in mouse embryonic stem (mES) cell cultures, along with 20+% and ∼10% of RPE and ganglion cell progenitors correspondingly. 5 The markers of other retinal neurons, such as horizontal, amacrine, and bipolar cells had been as well detected. While inhibition of Notch and Wnt signaling pathways promotes neural differentiation and rostral specification of neuronal tissues, the addition of the Notch and Wnt antagonists became an essential part for majority of monolayer protocols to improve the efficiency of retinal differentiation.5–9
Three-dimensional retinal differentiation method from ESCs first pioneered by Sasai group in 2011 revolutionized the field and became a promising strategy for retinal neuron production. 10 It is based on the recapitulation of retinal development and requires several stages: aggregate formation, neuroectoderm induction, and eye field induction/retinal maturation. The method relies on default developmental pathway and cultures are differentiated with no chemical, genetic, or cellular signaling pathway manipulation within scalable suspension culture.10,11 When grown in 3D culture systems, cells form multilayer cell aggregates composed of cells with different phenotypes, such as proliferating, nonproliferating, and necrotic. 12 In cell aggregates cell–cell and cell–extracellular matrix (ECM) interactions, gene and protein expression, and cell proliferation more closely mimic the natural environment found in vivo. It is expected that the morphology and architecture of the derived tissue will closely resemble its natural shape in the body opposing to the monolayer culture, where cells are stretched to the flat surface.
Three-dimensional retinal differentiation method results in the formation of multilayered retina with the efficiency of more than 70%.5,13 The neuroretinal tissue derived from mES cells has an aligned multilayer structure containing six major types of retinal neurons (photoreceptors, bipolar cells, ganglion cells, horizontal cells, amacrine calls, and Muller cells).10,11 Not only does it yield a high number of retinal cell populations for drug discovery and cell replacement therapies, but it also becomes a good model in vitro to study differentiation and maturation of retinal tissue.
With this advantage to mimic tissue microenvironment, 3D culture model has a broad application and is widely used as the testing platform in drug discovery. Cellular responses to drug treatment in 3D cultures have been shown to elicit cellular responses similar to the ones occurring in vivo. 12 For instance, cancer research widely adopted 3D culture technologies for drug development and screening, considering those as the gold standard for in vitro studies.14–18 This can reduce the number of animal studies and their associated costs, if targets are preliminarily screened in a more biologically relevant culture.
Despite all the advantages, the full potential of organoids in research and therapy has remained unrealized due to the existing limitations. Among those are issues of high variability and heterogeneity between cell lines and differentiation batches, extended time (time line for mES is around 30 days, whereas for human ES is up to 6 months), shortage of robust quantitative assays, and lack of automation. Nearly complete dependence of current organoid cultures on animal-derived matrices restricts their downstream clinical application. The ECM is a major component of the cellular environment and any in vitro expansion or differentiation of stem cells is built upon cell–ECM interactions.19,20 Mouse sarcoma-derived ECM Matrigel remains the most popular substrate for stem cell culture and is used for our retinal differentiation protocol. Matrigel and all other animal-derived matrices are not conductive to controlled modifications and pose the risk of immunogenicity and pathogen transfer.21–23 Current good manufacturing practice requirements demand strict characterization of all media, its components and additives,15,18 and animal-derived components should be avoided where possible.21,24 Furthermore, xenogeneic ingredients, including but not limited to Matrigel, do not have a defined chemical composition, contain numerous unquantified components, which contribute to variability observed between differentiation batches. 25
In this study, we approached the limitations of current 3D retinal differentiation from progenitor cell protocol. As heterogeneity in aggregate size and consequent differentiation efficiency depend in part on initial reaggregation, we studied the aggregation efficiency by comparing the effect of cell culture plate shape and material, medium viscosity, lipid, and bovine concentration on aggregate formation from mES cells. We also hypothesized that substituting Matrigel with chemically defined animal-free compound should yield more consistent results of differentiation. Hence, we studied the effect of vitronectin-mimicking oligopeptide (Synthemax II SC; Corning) as a signaling component for eye field induction.
In addition, we introduced semiautomated live imaging aggregates in 96-well plates using Image FL II imaging system. Overall, the experiments described below pursued two major goals: (1) study and improve aggregate formation efficiency; (2) investigate the effect of synthetic substitute of Matrigel Synthemax SC II on early retinal differentiation.
Methods
Retinal differentiation
Mouse ESC line EB5, in which GFP gene is knocked in under Rx promoter (rxGFP), was used for the experiments. 26 The baseline differentiation protocol we have is outlined below (Fig. 1). For induction of differentiation we dissociated the mES colonies at 70–80% confluence (maintained and passaged in mES medium with 10% FBS, 1 mM NEAA, 1 mM sodium pyruvate, 1% antibiotic–antimycotic, 0.1 mM 2-mercapthoethanol, and LIF 1000 μ/mL) (Supplementary Table S1). Cells were dissociated to single cells with Trypsin and seeded in cell suspension in the differentiation medium (optic vesicle [OV] medium, containing 1.5% FBS, 1 mM sodium pyruvate, 1% antibiotic–antimycotic, 1% chemically defined lipids, 0.1 mM 2-mercaptoethanol) (Supplementary Table S1) on 96-well V bottom polypropylene plates (Falcon), with seeding density of 3000 cells in 50 μL of OV medium per well.5,27 Matrigel (Corning) or Synthemax II SC was added at day 2 and 5 in the differentiation medium; at day 9, the medium got changed to optic cup medium (supplemented with NS2128), with subsequent changing every 2 days.
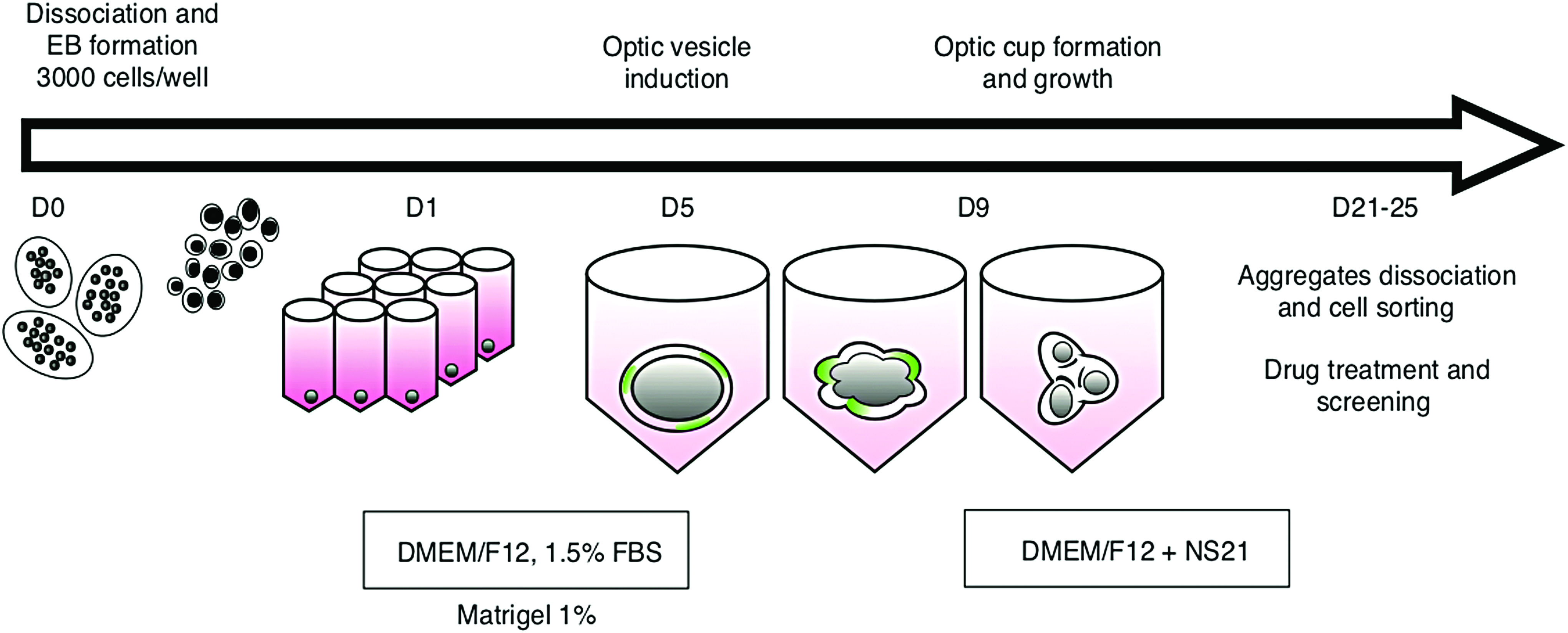
Schematic timeline and overview of the retina differentiation (SFEB) protocol. mES cells for all conditions are seeded at the same time in low-serum differentiation media. Matrigel (in concentration range from 0.5% to 4%) or Synthemax (concentration range from 0.005 to 0.08 mg/mL) is supplemented to the media at D2 and D5. At D9, the media are changed to the OC supplemented with NS 21. 28 DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum; mES, mouse embryonic stem; NS 21, neuronal supplement 21; OC, optic cup; SFEB, self-forming embryoid bodies. Color images are available online.
In an attempt to minimize the differentiation timeline, resuspending and seeding of cells, immediately from thawing the vial, was performed as well. Cell suspension in OV medium was seeded on 96-well V bottom polypropylene plates (Falcon), with seeding density of 3000 cells in 50 μL of OV medium per well.5,27
To study the aggregate formation and the factors that affect it, the following experimental design had been used. Twenty aggregates per condition were seeded (day 0) with seeding density of 10k cells/well, readout after 24 h (day 1). The factors chosen for testing were: well shape (V-bottom well, U-bottom well, flat-bottom well), lipid concentration in OV medium, viscosity (achieved by methylcellulose) of OV medium, and bovine serum albumin (BSA) concentration in OV medium. In 24 h, photos of plates were taken by G: Box Chemi XRQ system (Syngene), fluorescence achieved by adding 15 μL of diluted SYBR safe DNA stain (Invitrogen) to each well. Diameter of each aggregate was measured manually, and images were processed with ImageJ.
To study the effect of synthetic ECM Systemax SC II, the latter was added in place of Matrigel at day 1 and 5 of differentiation protocol. Concentration range from 0.005 to 0.08 mg/mL had been tested (1/200, 1/100, 1/50, 1/25, and 1/25 dilutions of initially resuspended Synthemax powder, respectively).
Live imaging
For live imaging, 10–12 embryoid bodies (EBs) from each group (each group represents different concentrations of Synthemax or Matrigel concentration added) were collected at D5 and D7 in culture and transferred to a Petri dish and imaged with inverted microscope Olympus CRX41, as well as with EVOS FL cell imaging system (Thermo Fisher). Notably, EVOS FL imaging system allows scanning of 96-well plates in automatic manner, creating separate image for each well. On separate run, two groups of organoids were started on a different type of 96-well plate (polystyrene transparent bottom with lid; Corning), and the scanning routine was created with the Celleste software. The latter plates were imaged automatically, both bright field and fluorescence, at several time points (D5, D7, D9, D15). Batch image processing was done with Fiji (ImageJ) (Fig. 2C, D).

Effect of lipid, methylcellulose addition to OV medium and 96-well shape on aggregate formation efficiency.
Aggregate immunohistochemistry
Twenty to 24 aggregates from each group were collected for cryosectioning, washed with PBS, and incubated in PFA 4% overnight at +4°C, after which PFA was replaced with 30% sucrose and aggregates were incubated at +4°C overnight. The following day EBs were moved to cryosection molds, excess fluid was removed, and molds were filled with embedding medium optimal cutting temperature medium (OCT) (Tissue Tek). Filled OCT molds were immediately frozen on dry ice and then transferred to −80°C for storage before sectioning. Prepared samples were sectioned using Leica CM 1850 Cryostat (14–16 μm thickness) and stored at −20°C before immunohistochemistry. Several aggregates from each group were separately processed for immunohistochemistry as whole organoids by mounting onto triple concavity slides (Lab Scientific).
The sections and whole organoids were washed with PBS for 5 min and placed in a blocking buffer (Triton × 0.1%, Tween 20 0.1%, 10% goat serum) for 1 h at RT. After blocking samples were incubated overnight with primary antibodies (Retinal RX Santa Cruz sc-79031 1/200, LHX2 Chemicon Abe1402 1/200, SIX6 Thermo Fisher A5-49779 1/200, PAX6 Hybridoma Bank 1/200, GFP Abcam 13970 1/2000, Ki67 Chemicon AB9260 1/200, Recoverin Chemicon AB5585 1/2000, Cone arrestin Chemicon AB15282 1/200, PKCa Santa Cruz sc8393 1/200, GFAP Zymed 18-0063 1/200, and pan-CD90 BioLegend 105202) (Supplementary Table S2). The following day, samples were washed twice with washing buffer and left in secondary antibody solution for 2 h at RT on orbital shaker or overnight for whole organoid stain (anti Rbt IgG Jackson Alexa 647 1/500, anti-Ms Invitrogen Alexa555 1/500 and Jackson Cy2, anti-Chicken Jackson Alexa 488 1/500, anti-rat Jackson Alexa 488). Secondary antibodies were washed and 1 μg/mL solution of DAPI (4, 6-diamidino-2-phenylindole) was applied for 1 min for cell nuclei counterstaining. After another two 5-min washes with PBS, sections and whole aggregates were mounted for further imaging. For imaging Nikon Eclipse E800, upright fluorescent microscope and Leica TCS SP5 confocal microscope were used.
Quantitative polymerase-chain reaction
Twelve aggregates at D7 were randomly collected and further processed for RNA extraction using the PureLink RNA Mini Kit (Ambion). Obtained RNA was reversed transcribed with SuperScript quantitative polymerase-chain reaction (qPCR) IV First-Strand Synthesis System (Invitrogen). RNA and DNA concentrations were measured with NanoDrop 2000 spectrophotometer (Thermo Scientific). qPCR plates were prepared using SyberGreen master mix and ran on StepOne Real-Time qPCR System (Applied Biosystems). The amount of cDNA normalized to LDH A housekeeping gene; gene expression at D0 of differentiation served as a reference group. The primers used are shown in Supplementary Table S3.
Flow cytometry
Ninety-six aggregates at day 7 of development were collected and dissociated with 1% Trypsin. After washing and spinning, the cells were strained (40 μm, Danville), fixed with 4% PFA on ice for 30 min. After another washing and spinning, cells were suspended in staining buffer and left overnight in primary antibodies. After 24 h, cells were spun, washed, and resuspended, secondary antibodies were added, and left at RT for 1 h of incubation. Following the last wash and spin, cells were resuspended in BD flow buffer and analyzed. Both BD LSR II and MACSQuant (Miltenyi Biotec) were used for flow cytometry sample processing. Obtained data were analyzed in FlowJo software. For pluripotency markers, analysis flow cytometry was performed, cell colonies were dissociated from culture flasks, washed, and fixed with 4% PFA for 30 min on ice. Conjugated antibodies, Oct3/429 and Nanog, 30 were added (PE; Miltenyi) (Supplementary Table S2) and left for 1 h of incubation. Following incubation, cells were washed, resuspended in BD flow buffer, and analyzed on BD LSR II flow cytometer.
Statistical analysis
Data analysis was performed in Prism Software, using t-test and ANOVA. p Values were two-tailed, and p < 0.05 was considered to be significantly different. All data are presented as mean ± SEM. Each experiment was replicated at a minimum of three times.
Results
Aggregate formation efficiency
Comparison of seeding conditions 24 h after seeding demonstrated the dose-dependent effect of lipids and methylcellulose on aggregate formation efficiency. Lipid concentration of 2% resulted in 100% efficiency of aggregate formation and significant increase in size to 532.8 ± 31.87 μm, p < 0.05); viscosity (methylcellulose concentration of 0.06% in OV medium showed 100% efficiency and increase in aggregate size 532 ± 19.23 μm, p < 0.01) (Fig. 2). However, well shape (U bottom vs. V bottom vs. 384w V bottom), have not shown any difference in formation efficiency and size of embryoid bodies, with the exception of flat-bottom plate, where they were not formed at all. Seeding cells immediately after thawing prevents aggregate formation. So far there is no cryopreservation method or medium that allows seeding right after thawing and the protocol requires at least one expansion passage.
The effect of xeno-free culture matrix on retinal differentiation
Our second goal was to determine if synthetic chemically defined ECM can induce early retinal differentiation and whether that effect is comparable to the one produced by Matrigel. Before starting the differentiation experiments, mES rxGFP cell line was tested for pluripotency markers. Flow cytometry for Nanog 29 and Oct 3/430 transcription factors showed more than 60% positive populations ensuring the pluripotency state of the cell line (Supplementary Fig. S1A). Immunohistochemistry for pluripotency genes demonstrated expression of Oct 3/4 and SSEA1 (Supplementary Fig. S1B). For Matrigel substitution, the effect of animal-free, synthetic vitronectin-based peptide Synthemax SC II was studied. We tested the range of Synthemax SC II (S) concentration range (0.005, 0.01, 0.02, 0.04, 0.08 mg/mL) along with Matrigel (M) concentrations (0.5%, 1%, 2%, 4%) in OV medium on induction of retinal differentiation in the 3D culture system.
Live imaging
We looked in this study at neuroectoderm induction and OVs evagination stages happening at DD5-9 of embryoid bodies' development.10,13,27 After aggregation, organoids from Synthemax groups formed neuroepithelial structures with rx-GFP+portions by D5 (Fig. 3A) and rx-GFP+OVs by D7 (Fig. 3B). By live imaging we observed comparable organoid morphology with rx-GFP+structures between Synthemax and Matrigel groups. During this experiment, we utilized new semiautomative imaging approach using EVOS AUTO FL imaging system. It allowed us to image 96-well plate of aggregates in a semiautomative fashion, taking both bright fields and fluorescent images for each well (Fig. 4A–C). The same scanning routine is applied to the plates; thus, the system gives an option to track each aggregate over time. Furthermore, it substantially reduces the time investment for imaging and can serve as perspective tool for future scale up.
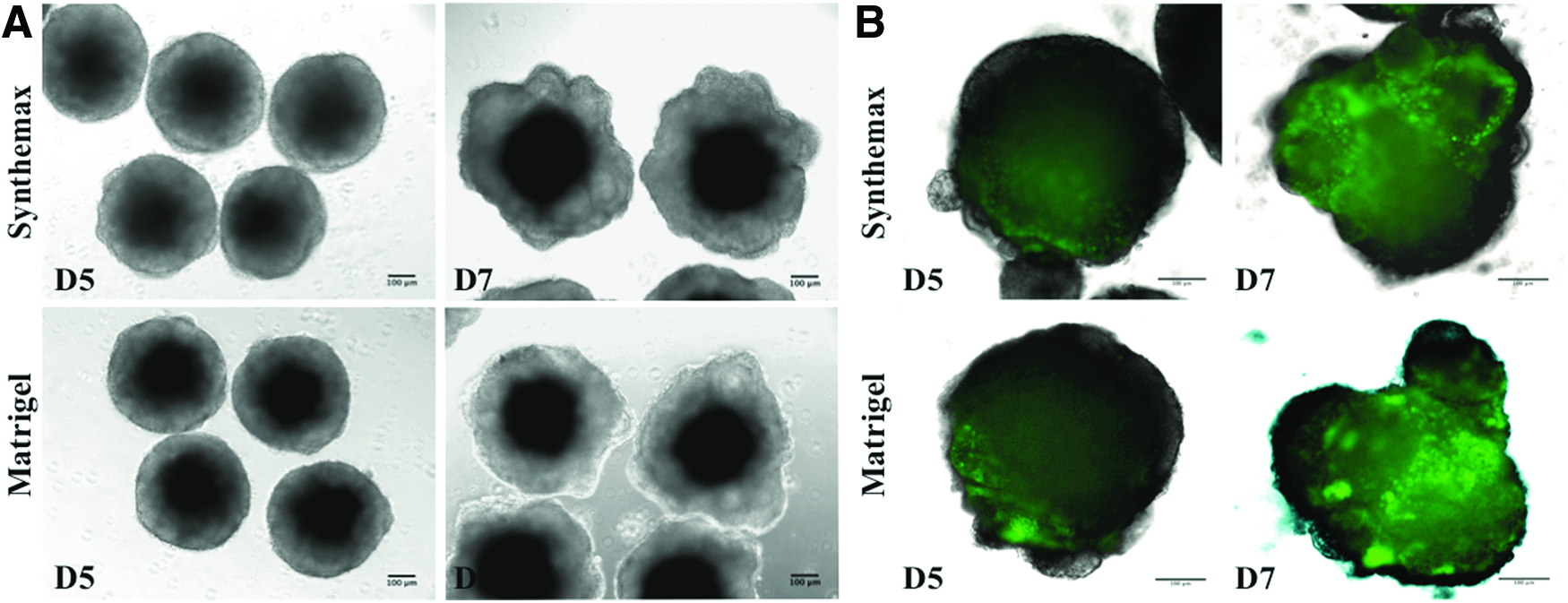
Aggregates of RX-GFP mES cell line at D5 and D7 in culture.

qPCR and flow cytometry
Eye field formation in embryo is characterized by expression of several transcription factors (Rx, PAX6, LHX2, SIX6, SIX3). The expression levels of Rx, Pax6, LHX2, and Six6 were tested for our groups at D7 by qPCR (N = 5, n = 4, where N is the number of biological replicates and n is the number of technical replicates). Upon our qPCR results, the expression of Rx, PAX6, LHX2, and SIX6 did not significantly differ between Synthemax and Matrigel groups at D7 in culture (Fig. 5A). However, no dose-dependent effect of Synthemax can be established. To assess whether cell proliferation is affected in groups supplemented with Synthemax, additional sample was added to flow cytometry experiment with stain for proliferation marker Ki67. Analysis shows 62.1% Ki67+events in Synthemax 0.005 mg/mL group compared with 65.5% Ki67+events in Matrigel 0.5% group.
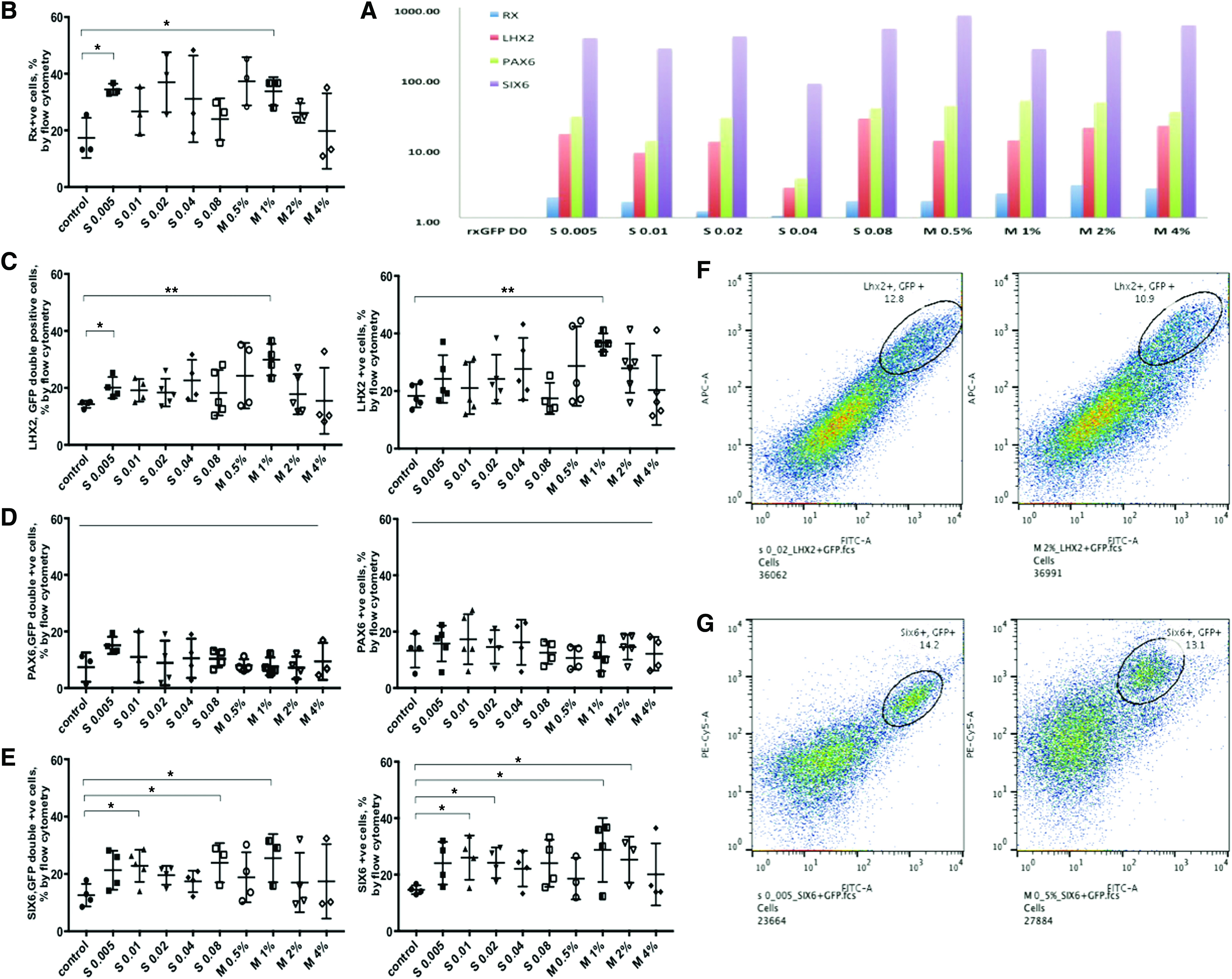
Effect of Synthemax and Matrigel concentration range on early eye field specification on day 7 of culture assessed by Flow Cytometry and qPCR.
Flow cytometry analysis results with staining for the genes Rx, LHX2, Pax6, Six6, and GFP are summarized in Figure 5B–E. Matrigel and Synthemax supplementation achieved comparable eye field induction rate across batches. Five biological repeats were analyzed. The average of 34.47% ± 1.15% Rx+events in Synthemax 0.005 mg/mL compared with average of 33.8% ± 2.9% Rx+events in Matrigel 1% group; the average 20.1% ± 1.86% LHX2, GFP double-positive events in Synthemax 0.005 mg/mL group compared with average of 29.9% ± 2.76% LHX2, GFP double-positive events; the average 22.82% ± 2.5% SIX6, GFP double-positive events in Synthemax 0.01 mg/mL group compared with 25.5% ± 4.85% SIX6, GFP double-positive events in Matrigel 1% group.
Although, the dose–response effect of Synthemax was not observed between groups by flow cytometry analysis, the low concentrations of latter (S 0.005 mg/mL and S 0.01 mg/mL) have resulted in the highest percentage of Rx +ve cells along with LHX2/GFP and SIX6/GFP double-positive cells. In addition, flow cytometry data, obtained during this experiment demonstrates the consistency between biological replicates and robustness of the adopted 3D differentiation protocol.
Immunohistochemistry and flow cytometry of aggregates
We performed immunostaining of organoids at D7 to confirm retinal induction. We detected early morphological features and expression of early eye field transcription factors in Synthemax groups, indicating retinal differentiation commitment. LHX2 and GFP coexpression, as well as SIX6 and GFP coexpression were observed in Synthemax group (Fig. 6A), and comparable to Matrigel group (Fig. 6B). Pronounced RX and PAX6 expression was detected on whole organoid stain (Supplementary Fig. S2). This pattern was observed at later stages as well: at D17 and D21 whole aggregates were collected for staining, and at D21 for flow cytometry analysis for late retinal neuron markers. Flow cytometry results confirm retinal differentiation of Synthemax-treated aggregates. Synthemax 0.01 mg/mL group has shown 6.77% of RNA-binding protein with multiple splicing (RBPMS)+events compared with 4.9% of RBPMS+events in Matrigel 0.5% group; 11.2% Recoverin+events in Synthemax 0.01 mg/mL group compared with 9.24% of Recoverin+events in Matrigel 0.5% group; 9.56% Cone arrestin+events in Synthemax 0.01 mg/mL group compared with 7.13% Cone arrestin+event in Matrigel 0.5% group; and 7.59% PKC A+events in Synthemax 0.01 mg/mL compared with 7.04% PKC A+events in Matrigel 0.5% group (Fig. 7B). Immunohistochemistry of organoids at D21 in culture validated development of retinal neurons in Synthemax groups, as confirmed by positive staining for pan-retinal ganglion cell marker RBPMS (Fig. 7A).
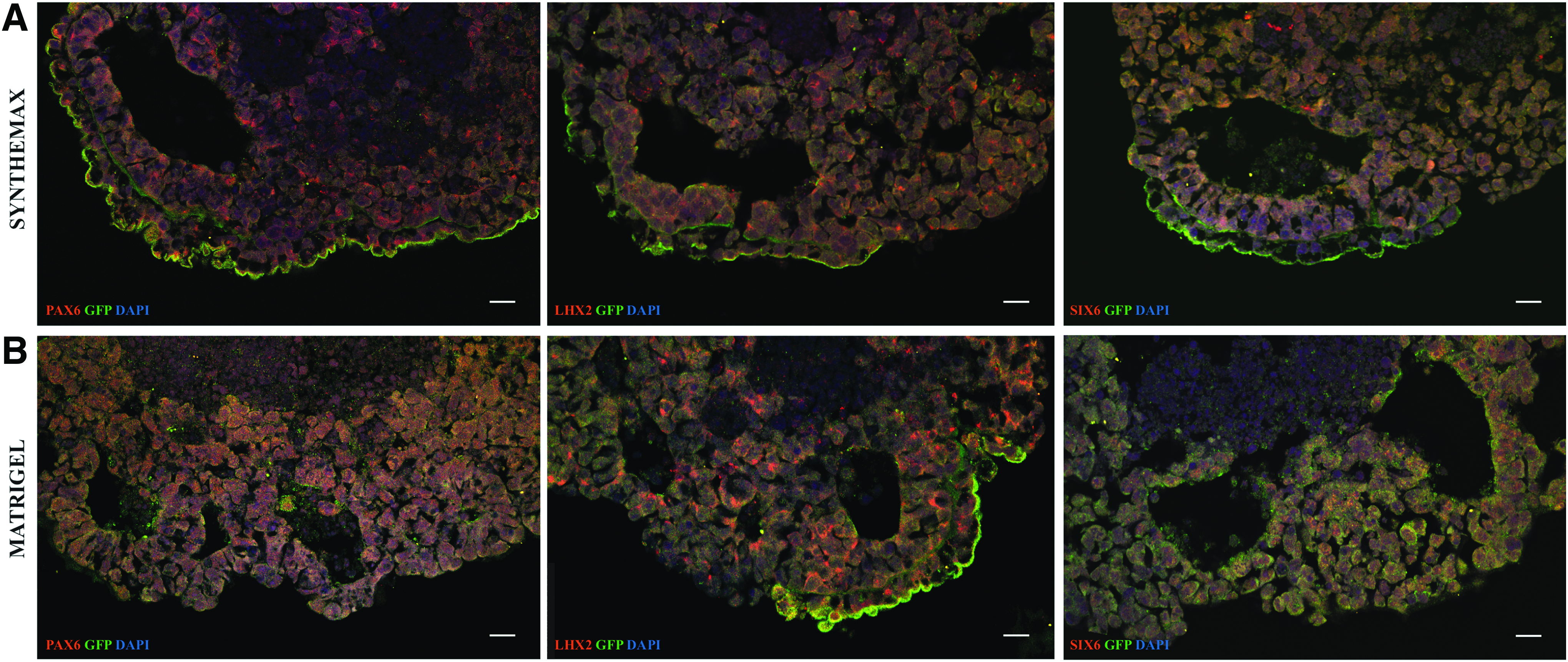
Immunohistochemistry of aggregates at D7 in culture. Synthemax 0.005 mg/mL
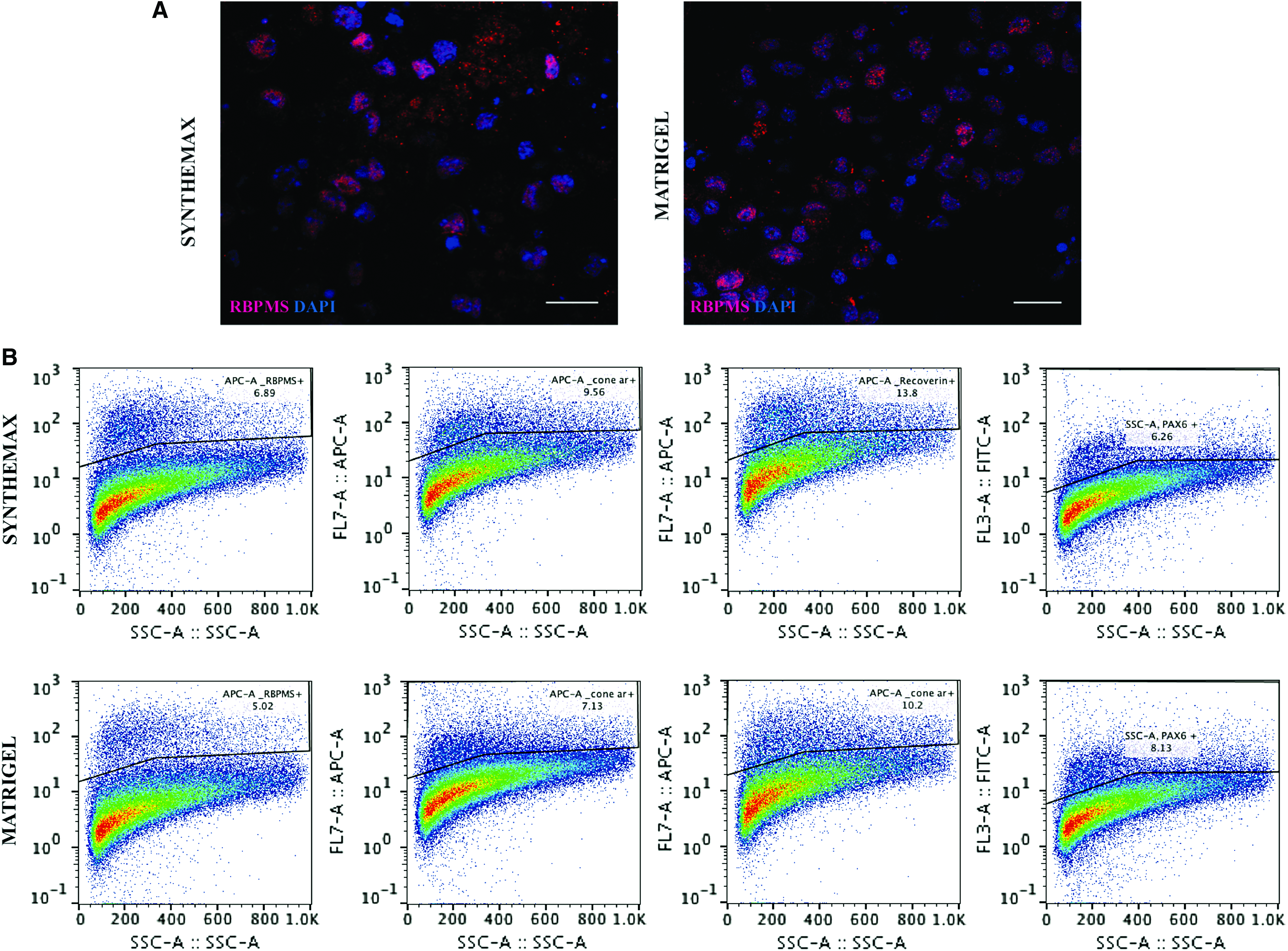
Immunohistochemistry and flow cytometry of aggregates at D17 and D21 in culture.
Few randomly collected organoids at D15 were as well stained with proliferation marker Ki67. The observed staining patterns obtained in Matrigel 0.5% and Synthemax 0.005 mg/mL groups confirm that synthetic matrix Synthemax supports the stable proliferation even in mature organoids. Flow cytometry and immunohistochemistry with proliferation marker stain are summarized in Supplementary Figure 3A, B.
Discussion
With retinal degenerative diseases remaining largely untreatable, the need for cell therapy grows. The animal studies show the efficiency of cell replacement for retinal pigment epithelium and photoreceptors. The pioneering work of Sasai group10,31 and others demonstrated that it is possible to drive differentiation of both mouse and human ES toward the retina by recapitulating the development program in 3D culture, making this approach the major tool for retinal neuron differentiation for cell manufacture for transplantation and drug discovery.
However, the full potential of the organoid system remains unrealized, in part, due to the presence of Matrigel animal-derived matrix, used to induce neuroepithelia formation and support tissue-like structure. Synthetic substrates are a promising alterative for the stem cell cultures. Numerous studies within and outside the retinal field initiated to design synthetic matrices for stem cell culture and differentiation.32–34
As typical extracellular matrices, including Matrigel, consist of different proteins, including collagen, vitronectin, laminin, heparin sulfate, fibronectin, proteoglycans, entactin, and some undefined factors,19,25 using ECM-mimicking peptides could be the potential solution. The term ECM-mimicking peptide refers to the peptides that promote, induce, or maintain cell attachment, proliferation, spreading, viability, and/or differentiation. For instance, the use of fibronectin and laminin-mimicking peptides has been explored.34,35 Synthemax II SC is a short oligopeptide, KGGPQVTRGDVFTMP, based on Arg-Gly-Asp (RGD) motif, designed after vitronectin protein. 36 Synthemax was originally developed for hESC culture and cardiomyocyte differentiation of hESC in different media, although later it was also used for iPSC derivation and maintenance and retinal pigment epithelium differentiation.37,38 Synthemax acts as integrin receptor ligand, mainly ανβ5, and integrin signaling has been shown to be critical for retinal development.37–41
In this study, we have studied the effect of Synthemax on retinal differentiation from mouse ESCs. Our data demonstrated that this chemically defined oligopeptide induces early retinal differentiation at the level no different from Matrigel, and can replace the latter in widely used 3D “Sasai” protocol for retinal differentiation. Concentration range of Synthemax used for our experiments (0.005–0.08 mg/mL), did not show any marked dose–response. It is probably related to the stochastic nature of eye field induction, where Integrin receptors need to be activated. Yet, supplementation with low concentration of Synthemax SC (0.005 and 0.01 mg/mL) resulted in statistically significant increase in early eye field gene expression assessed by flow cytometry and more pronounced morphological development compared with a group with no ECM added, assessed by live imaging. Our results propose low concentrations of Synthemax (0.005 and 0.01 mg/mL) being optimal for induction of retinal differentiation. Aggregates formed in Synthemax groups demonstrated similar morphology and timeline of their development to those grown with Matrigel supplementation. qPCR assessment of Rx, LHX2, and SIX6 expression in aggregates of D7 in culture did not reveal any significant difference between Synthemax and Matrigel groups. Although, we have not observed significant increase in retinal yield after the switch to synthetic matrix from Matrigel, we believe that Synthemax at used concentrations provides sufficient stimuli for eye field induction and retinal maintenance.
The role of this synthetic ECM on mouse and human pluripotent cell proliferation and apoptosis in expansion conditions have been studied before with no significant difference observed between Matrigel and Synthetic ECM groups. 42 Matrigel and Synthemax have also been compared as substrate in multiple differentiation conditions, including the differentiation of neural retina, retinal pigment epithelium, and cardiomyocytes. No significant difference was found. However, the comparison of synthetic ECM and Matrigel in 3D differentiation of retina has not been performed, although previous work suggests that the differences (if any) will be subtle. To evaluate the effect of Synthemax on cell proliferation in organoid culture, we performed flow cytometry and immunostaining or organoids at D7 and D15, respectively, for proliferation marker Ki67. No significant difference between organoids of Matrigel and Synthemax groups has been detected.
Optimization of retinal differentiation is an essential step to bring cell manufacture closer to clinical applications. Various different research groups explore the potentials for 3D retinal differentiation protocol improvement,13,43 reporting chemical and physical factors affecting yield, efficiency, and reproducibility of differentiation. In this work, we studied aggregate formation efficiency as one of the ways to reduce organoid heterogeneity within and between the batches.
The formation of aggregates is a crucial initial step of any organoid culture and can be defined as 3D assembly of certain number of singularized cells. Cells that did not incorporate into embryoid body are less viable, typically undergo apoptosis, and contribute to inefficiencies of aggregation process. 44 In this study, we measured the size of formed aggregates to assess their formation efficiency. As previously shown, the size of formed embryoid body is important, yet simple physical parameter affects further cell differentiation toward different lineages.45,46 Besides, Sharma et al. with their work have shown that the size of formed embryoid bodies can influence retinal differentiation, confirming increased expression of early eye field transcription factors, such as Rx, SIX3, and Otx2, in larger aggregates. 44 Moreover, better control and increased homogeneity of aggregate size will add to their more synchronized development toward retinal fate.
Curtis et al. in their study of plasmalemmal lipids' effect on cell adhesion verified that adhesion increases as fatty acid content increases after incubation of neural retinal cells in lipid-enriched media. 47 Newer approaches, such as liposome delivery systems, allow cell surface modification by incorporation of lipids that drive cell–cell adhesion. 48 On the other hand, serum albumin, widely used for cell culture, is known for its unique capability to bind a variety of ligands and interact with the cell surface. 49 Hence, we hypothesized and tested the effect of chemically defined lipid concentrate and BSA concentration in OV medium, as well as viscosity of the medium, on aggregates' formation efficiency. Viscosity is a physical property of any liquid and can be modified by the addition of various macromolecules. 50 Recent work of Yoshida et al. have demonstrated that natural polymer dextran can precipitate cell adhesion and facilitate 3D cellular assembly through depletion effect. 50 In our experiments, we used methylcellulose in 0% to 1% concentration range to modify the OV medium viscosity. Aggregate diameter was measured 24 h postseeding with size being an estimate of proper cell assembly.
Our data demonstrated that both lipid concentration and medium viscosity affect aggregate formation efficiency and size. Addition of either 2% lipid concentrate or 0.06% methylcellulose to OV medium resulted in most pronounced aggregate size increase. Interestingly, the sizes of the aggregates seeded in 10,000 cells per well under these conditions are close to identical, being 532 and 532.8 μm, respectively, for the 2% lipids and 0.06% methylcellulose groups.
Being a simple physical parameter, the size of formed aggregates is influenced by many factors, including but not limited to seeding density, medium composition, well shape, as well as culture handling, aggregation method, and initial status of pluripotent cell colonies.44,45 The size of reported embryoid bodies' size in 24 h after seeding varies prominently between and within laboratories. The variability range can be as high as 300 μm.11,13,27,43–45 In our laboratory, we observe the aggregate size ranging from 300 to 600 μm within tested physical parameters. Without any medium manipulation, the vast majority of formed aggregates fall in the 380–550 μm-size bracket, when seeded in 10,000 cells per well. Assuming other factors are evened by short seeding experiment time and one person handling, the obtained matching values in aggregate size in lipids 2% and methylcellulose 0.06% groups may be explained by creating optimal physical environment for cell adhesion.
Better control of organoid size in the first place will enhance the homogeneity of 3D cultures and will help to achieve more reproducible retinal differentiation. To further validate the benefits of described conditions on retinal differentiation in 3D cultures, additional in vitro and in vivo experiments are to be performed. The yield and viability of isolated retinal neurons could be compared between discussed conditions as well as between differentiation batches. Given that the cell replacement is an ultimate goal, various animal cell transplantation studies can be completed. Therefore, with applied protocol to retinal organoid system, research on donor retinal cell survival, integration, and functionality are among our further steps.
Timing and lack of automation for cell culture and analysis are other limitations we observed during this study. We implemented a novel semiautomatic imaging manner of aggregate plate using EVOS FL autoimaging system, which will allow substantial scale up of aggregate expansion. Moreover, it will enable long-term tracking of single aggregate and become a valuable tool to further study retinal differentiation and aggregate heterogeneity between wells.
Conclusions
In this study, we present a set of optimized conditions for 3D retinal differentiation from pluripotent cells. We suggest that 2% lipid concentrate or 0.06% methylcellulose to OV medium will improve reaggregation of cells after seeding. We confirm with our data that use of synthetic vitronectin-mimicking peptide, Synthemax SC, is efficient to induce early retinal differentiation. The efficiency of retinal differentiation induction has no dose–response effect and is comparable to the one observed with Matrigel. Nevertheless, our results propose low concentrations of Synthemax being optimal for induction of retinal differentiation. We suggest these defined medium conditions will improve reproducibility, and decrease the risk of contamination while allowing substantial scale up of retinal tissue and cell production for drug discovery, disease modeling, and transplantation purposes.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.