
Abstract
The classic bone tissue engineering model for bone regeneration combines three elements: scaffolds, biomaterials, and mesenchymal stem cells (MSCs). Incorporation of MSCs and growth factors into a scaffold implanted into the area of bone injury is a proven strategy to achieve successful bone regeneration as demonstrated in the literature. However, a major limitation of using bone grafts or scaffolds is oxygen (O2) deprivation in the inner sections of the construct, due to lack of adequate vascularization. To address this limitation, we proposed two treatment strategies for MSC-seeded constructs or adipose tissue scaffolds before implantation: (1) O2 enrichment and (2) acclimation to hypoxia. Based on previous studies, the significance of the different O2 concentrations on MSC biological characteristics remains controversial. Therefore, the optimal O2 condition for engineered bone tissues should be determined. Thus, we designed an innovative multichamber gas system aimed to simultaneously assess the effects of different O2 levels on cell culture. This system was assembled using three isolated chambers integrated into a single incubator. To explore the efficacy of our method, we investigated the effect of hyperoxia, normoxia, and hypoxia, (50–60%, 21%, and 5–7.5% O2, respectively) on the biological characteristics of human adipose-derived MSCs: immunophenotyping, adhesion, proliferation, and osteogenic, and angiogenic differentiation. Our findings demonstrated that hypoxic adipose-derived mesenchymal stem cells (ASCs) conditions exhibited significantly lower levels of CD34 (p = 0.014), with significantly higher osteogenic and angiogenic differentiation capacities (p = 0.023 and p = 0.0042, respectively) than normoxia. Conversely, hyperoxia-cultured ASCs demonstrated significantly higher levels of CD73 and CD90 expression than both normoxic ASCs (p = 0.006 and p = 0.025, respectively) and hypoxic ASCs (p = 0.003 and p = 0.003, respectively). In addition, hyperoxic ASCs showed significantly reduced proliferation capacity by day 11 (p = 0.032) and significantly enhanced migration rates after 48 h (p = 0.044). The newly developed controllable multichamber gas system was cost-effective and easy to use. Different assays can be performed concurrently while preserving all other conditions identical, and the use of other ranges of O2 concentrations is feasible and also necessary to determine the ideal O2 concentration. Furthermore, the multichamber gas system has the potential for wide application, including other cell cultures, grafts, or scaffolds for in vitro and in vivo experimentation. This study was approved by the Galilee Medical Center Helsinki Committee (No. 0009-19-NHR).
Impact statement
The introduced multichamber gas system provides a custom-made setup for simultaneous control of three oxygen (O2) levels in a single incubator. The use of our innovative multichamber gas system is essential to determine the ideal O2 levels for engineered tissues by examining multiple O2 concentrations on cells in vitro. The determined ideal O2 concentration will then be used through this system to investigate the engrafted cell survival ex vivo, to ensure successful integration of the engineered tissues and tissue regeneration in vivo. Use of this method may promote a therapeutic tool for a major limitation in tissue engineering due to the problematic O2 insufficiency in tissue scaffolds.
Introduction
Vascularization is most critical for the survival, integration, and functionality of engineered bone tissue, as it assures efficient gas and nutrition exchange within the regenerated tissue.1,2 Clinically, both the disruption in the vasculature and the subsequent hypoxia within the wound microenvironment are the primary risk factors for reduced bone regeneration.3,4 Subsequently, graft necrosis and failure to integrate with the host tissue due to ischemia and inadequate vascularization are the main causes of premature failure of implanted constructs or scaffolds.5,6 Recently, many attempts have been made to overcome the insufficient vascularization in engineered tissues, by engineering blood vessel systems, known as vascularized tissues, using both scaffold-based and cell-based techniques including coculture strategies.5,7,8
According to preclinical studies, cell-seeded constructs are proven to enhance more bone regeneration in comparison with acellular constructs.8,9 Yet, clinical translation of these tissues is still limited. 10 Therefore, cultivating mesenchymal stem cell (MSC)-seeded constructs under ideal oxygen (O2) levels during the engineering process might enhance cell proliferation and osteogenic and angiogenic differentiation, crucial for successful bone regeneration.11–13 To this end, we proposed two treatments to ensure adequate O2 supply to cell-seeded constructs or grafts before implantation: (1) O2 enrichment and (2) acclimation to hypoxic conditions. O2 enrichment is a strategy applied in hyperbaric oxygen (HBO) therapy, and involves repetitive exposure to hyperoxia. HBO is approved for multiple pathologies, and has been shown to promote neovascularization and stem cell recruitment, leading to accelerated tissue regeneration. 14 Hypoxia both mimics the physiological niche of stem cells and simulates the inner part of the ischemic construct,3,15 and has been suggested to present suitable conditions for cells to adapt to the hypoxic state.
The different tissues in our bodies experience different O2 levels, depending on the physiological O2 demands and environment.12,15–20 Remarkably, despite low O2 concentrations in the stem cell niche in the human body (1–8%), MSCs and many other tissues are cultured in conventional incubators at relatively high O2 concentrations (18–20% O2), which are considered “normoxic,” corresponding to atmospheric O2.15,21 However, the impact of O2 concentrations on the characteristic properties of MSCs in vitro remains controversial. Extreme hypoxia (2% O2) reduced osteogenic differentiation of murine adipose-derived mesenchymal stem cells (ASCs) and human ASCs (hASCs).11,22 In contrast, moderate hypoxia (5% O2) lowered proliferation rates, yet enhanced the chondrogenic potential of hASCs. 23 Severe hypoxia (1% O2) decreased proliferation and differentiation of human MSCs (hMSCs), whereas elevation of O2 levels to 3% restored osteogenic differentiation, and 21% O2 induced high proliferation rates. 21 In contrast, other studies found that 2% O2 significantly improved expansion characteristics, and 5% O2 enhanced the proliferation and osteoblast differentiation rates of hASCs.24,25
This remarkable lack of consensus in the field may pose confusion when comparing results across studies. Consequently, a novel means of preserving consistency of the experimental setup is critical. The few existing models used to investigate the effect of O2 on cells suffer from many drawbacks and some have not been properly described. For example, a hypoxia model as a cell culture incubator in which O2 is displaced by an infusion of nitrogen (N2) to mimic hypoxic conditions is a quite expensive and large model, which may be infeasible or inconvenient for small laboratories that do not perform hypoxia experiments on a daily basis.26,27 Similarly, use of a special incubator for each O2 condition under assessment is also expensive and space consuming. Another commercially available model is the hypoxia incubator chamber that fits inside the existing incubator. This chamber is expensive to purchase that is inapplicable especially if multiple units are required for simultaneous testing of different O2 levels. As an effective low-cost solution, we established a custom-made multichamber gas system, assembled of three separate chambers, as an alternative experimental model, allowing for simultaneous control of three O2 levels in a single incubator. This system was designed to enable concurrent investigation of the effect of three different O2 levels on cell culture. This study harnessed the multichamber incubator to assess the effect of hyperoxia, normoxia, and hypoxia on hASCs.
Methods
The multichamber gas system was constructed from three transparent polypropylene storage boxes (30.1 × 18.8 × 9.4 cm; Lock & Lock, Hanacobi, Korea), with two holes (6 mm diameter) drilled in their sides. Two push-in bulkhead connectors (6 mm; Metal Work Pneumatic) were inserted into the holes and sealed using epoxy glue. The connectors were connected to CO-PU tubes (Castello), which served as inlet and outlet ports for gas exchange (Fig. 1A–C). Each box served as a growth chamber, with all three boxes maintained in the same incubator (New Brunswick Galaxy 170S) to conserve the same temperature (37°C) and relative humidity (95%). O2 concentration was independently set for each chamber. A premixed gas cylinder was connected to the inlet port of each chamber through a CO-PU tube, along with a flow meter to maintain adequate and low air pressure. Each gas cylinder was filled with the desired O2 concentration, while preserving 5% carbon dioxide (CO2) (Fig. 1D, E). This research examined the following mixtures: hyperoxia (60% O2), normoxia (21% O2), and hypoxia (5% O2), with 5% CO2 and N2 for the remaining gas content (35%, 74%, and 90%, respectively). We intended to examine these O2 levels since 21% is the atmospheric O2 concentration used in conventional incubators, whereas 60% O2 is a mean range between extreme hyperoxia 85–100% O2 and the atmospheric 21% O2. According to our preliminary study using two different incubators, short-term hyperoxia (85% O2) exhibited higher adhesive and proliferative capacities than normoxia, yet long-term hyperoxia reduced ASC proliferation. Regarding hypoxia, 5% O2 is considered a moderate hypoxic concentration according to the literature.23–25 However, any gas mixture can be used in this setup.
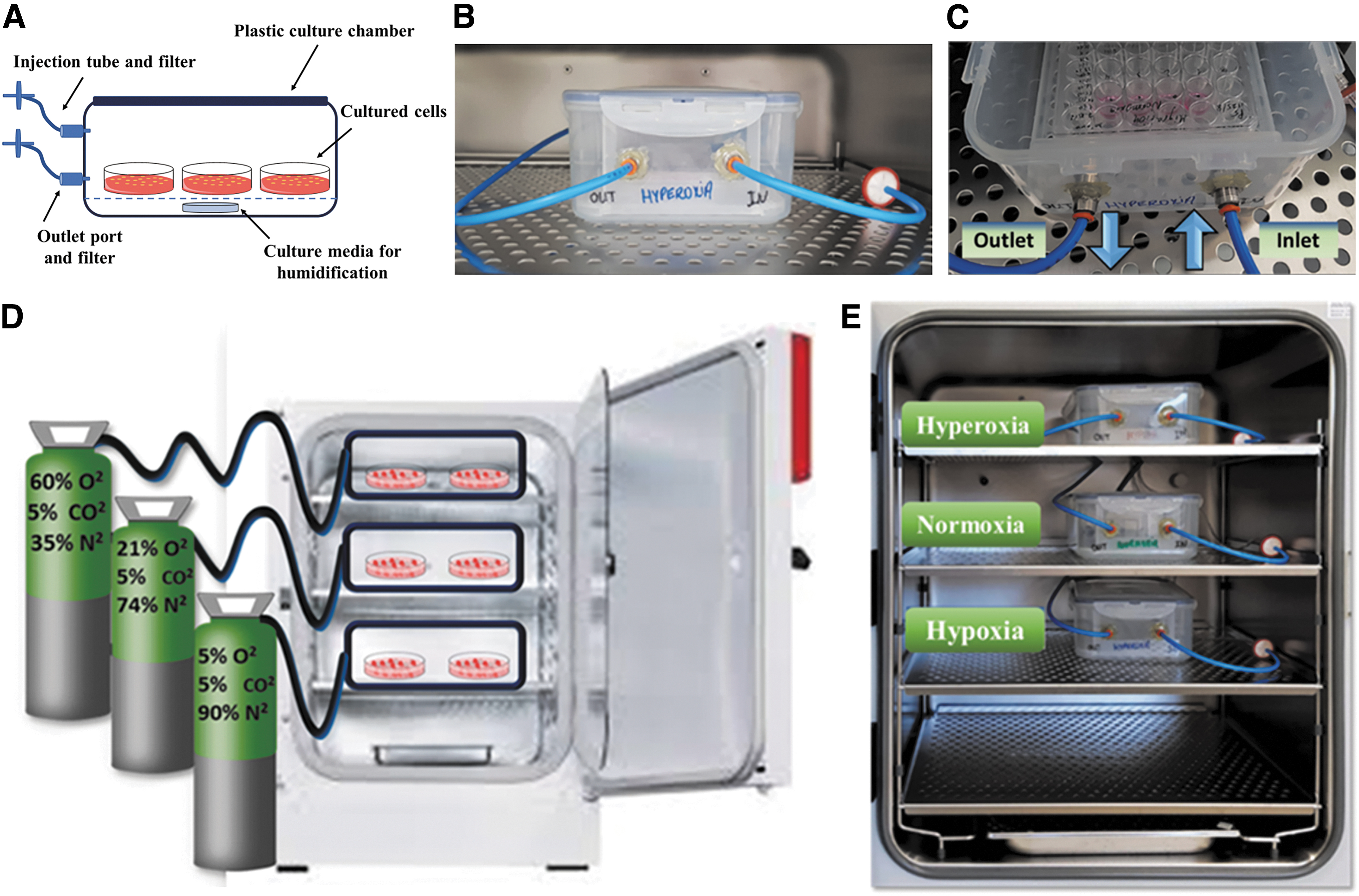
Controllable multichamber gas system in a cell culture incubator. Three transparent polypropylene chambers (30.1 × 18.8 × 9.4 cm) attached to inlet and outlet gas ports were filled with prespecified gas mixtures cultivated in a single incubator.
Before utilizing the chambers for cell culture, they were first sterilized using common sterilization methods, including 70% ethanol and ultraviolet radiation for at least 2 h. In addition, a small sterile gauze pad soaked with autoclaved water containing 1% Aquaguard-1 (Biological Industries) was placed inside each chamber to provide proper moisture, similar to a common humidified CO2 incubator (Fig. 1A).
The chambers were infused with the prespecified gas mixtures for 15 min every day, to ensure proper displacement of atmospheric air. The flow rate of gas mixtures was set to ∼5 L/min using adjustable flow meter connected to the inlet tube. O2 concentration in the chamber was measured using an O2 analyzer (Servomex) connected to the outlet tube. The outlet port was sealed immediately thereafter until the next gas infusion. To confirm achievement of the desired O2 concentrations, O2 concentrations in each chamber were measured daily using a Servomex O2 analyzer, immediately after gas infusion and on the following day (before the subsequent infusion).
Experiment
Experimental Design
ASC isolation and culture
Adipose-derived stem cells (ASCs) were isolated from human subcutaneous adipose tissue using the rapid isolation method established by Zeng et al.
28
The procedure and subsequent research were approved by the Galilee Medical Center Helsinki Committee (No. 0009-19-NHR). In brief, human adipose tissue was obtained from patients undergoing lipectomy, after receiving their signed informed consent. After surgery, adipose tissue samples were cut and washed twice with phosphate-buffered saline (PBS), supplemented with penicillin–streptomycin–amphotericin (200 U/mL, 200 μg/mL, and 0.5 μg/mL, respectively; Biological Industries). The small tissue explants were placed in cell culture dishes in basic complete growth medium (CGM), comprising Dulbecco's modified Eagle medium (DMEM), supplemented with 10% fetal bovine serum (FBS),
Flow cytometry analysis
ASCs cultured in a conventional CO2 incubator were characterized by flow cytometry to validate MSC phenotype. Flow cytometry was also used to characterize ASCs cultured under either hyperoxia, normoxia, or hypoxia for 1 week in the multichamber gas system. For ASCs immunophenotyping in all culture conditions, ∼1 × 106 cells were harvested and centrifuged. Next, they were incubated for 15 min at 4°C, in the dark, with phycoerythrin (PE)-conjugated antibodies against human CD45, CD73, CD34, CD11b, CD19 (BD Biosciences), CD90 (Merck), CD105 (Biolegend), or HLA-DR (R&D Systems). Analysis was performed using a Navios flow cytometer and Kaluza software (Beckman Coulter).
Adhesion and proliferation analyses
ASCs were seeded in 24-well plates (7.5 × 103 cells/cm2) in basic CGM, under different O2 concentrations, and cultured in the multichamber gas system for 2 weeks. Cell adhesion was determined 1 day postseeding by washing the cells with PBS and incubating them in CGM containing 10% AlamarBlue dye (Bio-Rad) at 37°C for 2 h. Fluorescence was recorded using a SpectraMax M2 ELISA (enzyme-linked immunosorbent assay) reader (Molecular Devices). The proliferation rate was determined by performing the same procedure on days 4, 8, 11, and 15 postseeding. Results are presented in relative fluorescence units, which correlate with the number of live cells. For detailed information about the effect of different oxygen concentrations on ASC morphology, see the Supplementary Data.
Wound healing—scratch assay
ASCs were seeded in 24-well plates (15 × 103 cells/cm2) until ∼90% confluence. Then, a scratch was made across each well using a 100 μL pipette tip. Plates were washed twice with PBS and then cultured, in basic CGM, under different O2 levels in the multichamber gas system. Cell migration was assessed by observing the scratch area and photographing it with an inverted Olympus microscope and Nikon camera, immediately and 24, 48, and 72 h after making the scratch. The wound extent (the percentage of the area not occupied by cells, relative to scratch area at 0 h) was determined using ImageJ software.
Assessment of osteogenic differentiation
To induce osteogenic differentiation, ASCs were cultured in osteogenic medium (DMEM, 1 × 10−8 M dexamethasone, 100 μg/mL
In vitro ASC angiogenic differentiation
To assess angiogenic differentiation of ASCs, cells were cultured under various O2 concentrations in complete endothelial cell growth basal medium-2 (EBM-2). This medium was supplemented with growth factors: vascular endothelial growth factor (VEGF), hydrocortisone, hFGF-B, R3-IGF-1, ascorbic acid, hEGF, GA-1000, and heparin according to the manufacturer's instructions (Lonza), and 20% FBS,
Tube (capillary) formation assay
ASCs were seeded in six-well plates (7.8 × 103 cells/cm2) in complete EBM-2 and then cultured under three different O2 conditions for 1 week. Capillary formation was analyzed by coating three 24-well plates with Matrigel (250 μL/well; Corning) for 25 min at 37°C. Then, ASCs (5 × 104 cells/cm2) were seeded in 200 μL complete EBM-2 onto the Matrigel-coated plates and incubated under three different O2 levels. Tube formation was assessed after 2 and 4 h of incubation, as lumen-containing tubules begin to appear usually within 2 h. Using an inverted Nikon microscope, cell cultures were photographed, so the number of tubes formed can be easily assessed as an evaluation for in vitro angiogenic differentiation. Quantitative analysis of tube formation was determined through the number of capillary-like structures after 4 h using ImageJ software.
Human VEGF quantification assay
ASCs were cultured in 24-well plates (15 × 103 cells/cm2) in EBM-2, under three different O2 conditions, for 1 week. Samples of culture supernatant were collected after 3 and 6 days of culture, and frozen at −20°C. To measure the amount of human VEGF (hVEGF) secreted from ASCs, an ELISA was performed using the Quantikine Human VEGF Immunoassay kit (R&D Systems) according to the manufacturer's instructions. hVEGF concentration was determined through its optical density at 450 nm, as measured using a Varioskan Lux spectrophotometer.
Statistical analysis
Prism 8.0.1 Software (GraphPad) was used for statistical analyses. Descriptive statistics include means ± standard deviation. Comparisons between samples cultured under different O2 conditions, and between different time points, were performed using the one-way and two-way analyses of variance. p-Value <0.05 was considered statistically significant.
Experimental Results
Creation of the desired O2 environment through the multichamber gas system
The described multichamber gas system was successfully developed. O2 content measured immediately after gas infusion into the hyperoxia, normoxia, and hypoxia chambers was 60%, 21%, and 5% O2 levels, respectively. After 24 h, O2 concentration in hyperoxia chamber has changed from 60% to 53.4% ± 0.44% O2 and that in the hypoxic chamber changed from 5% to 7.4% ± 0.36% O2, while it remained stable in the normoxic chamber (21% to 20.98 ± 0.04) (Fig. 2). Thus, characterization of ASC was examined under different O2 concentration ranges: hyperoxia (50–60% O2) and hypoxia (5–7.5% O2). Accordingly, the chambers maintained the required range of hyperoxia, normoxia, and hypoxia and were able to sustain the ratio of gas components for short-term and long-term experiments.
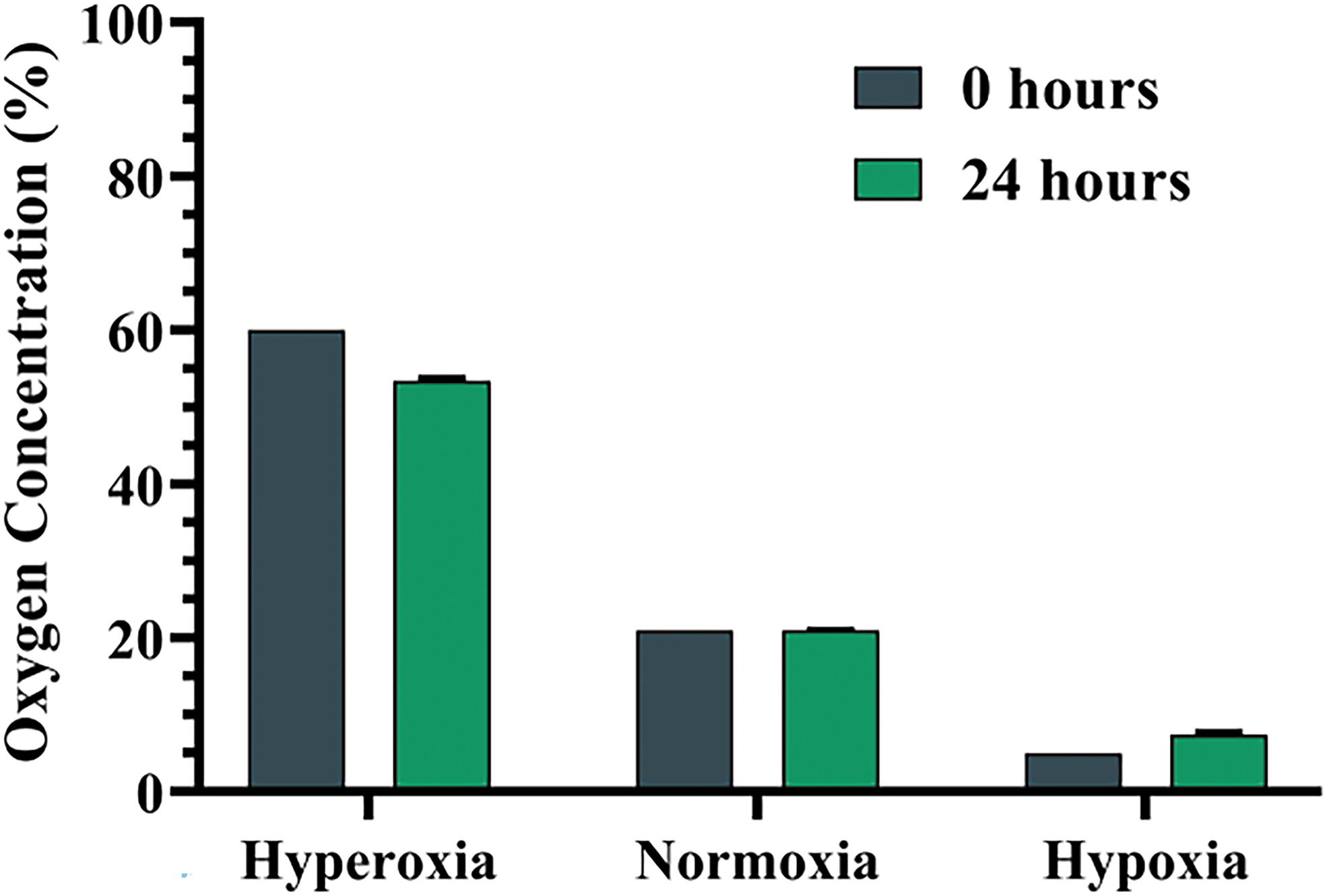
Oxygen (O2) concentrations inside the multichamber gas system chambers. Measurements of O2 concentrations (as a percentage of the gas O2) in the three chambers immediately (0 h) and 24 h after each gas infusion. Results are presented as mean ± SD (n = 9). SD, standard deviation. Color images are available online.
ASC characterization
ASCs (P3) cultured in a conventional CO2 incubator demonstrated the characteristic MSC phenotype. ASCs were positive for the stromal surface markers CD73 and CD90, and negative for the hematopoietic lineage surface markers CD45, CD34, CD19, CD11b, and HLA-DR. When cultured in the multichamber gas system for 7 days, there were no significant differences in the percentages of ASCs positive for each surface marker among the different O2 levels (hyperoxia, normoxia, and hypoxia) (Fig. 3A, B). However, analysis of the mean fluorescence intensity of each marker demonstrated that hyperoxic ASCs had significantly higher levels of CD73 than ASC grown under normoxic or hypoxic conditions (p = 0.006, p = 0.003, respectively). Similarly, CD90 levels were significantly higher in hyperoxic ASCs than in normoxic and hypoxic ASCs (p = 0.025 and p = 0.003, respectively) (Fig. 3C). The expression of CD34 was significantly lower in hypoxic ASCs than in normoxic conditions (p = 0.014) (Fig. 3D).
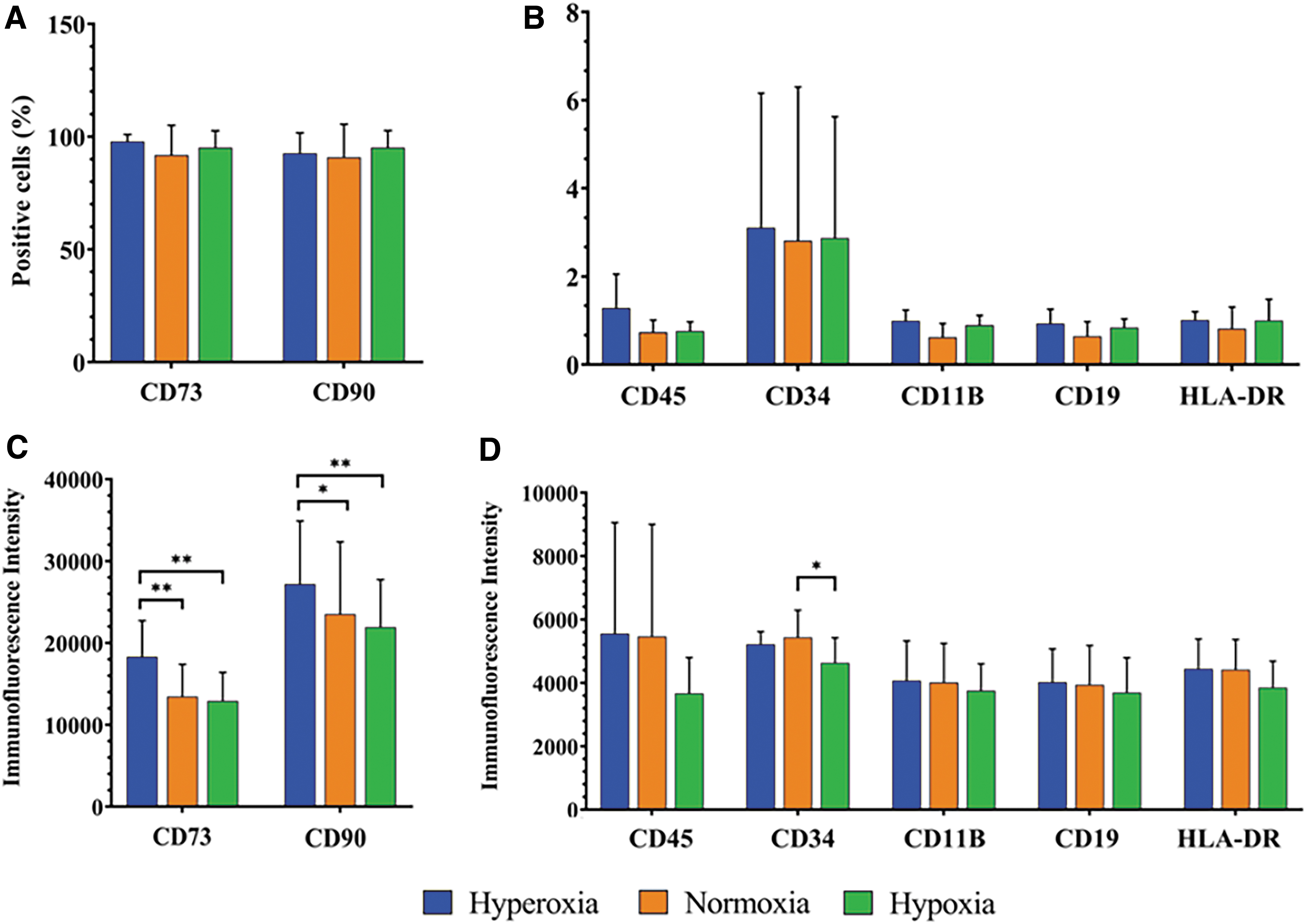
Immunophenotyping of ASCs after exposure to three O2 levels. Flow cytometric analysis of ASCs after exposure to hyperoxia (50–60% O2), normoxia (21% O2), or hypoxia (5–7.5% O2). Percentage of ASCs expressing
Effect of different O2 concentrations on ASC proliferation
No major differences in ASC adhesion were noted across the three O2 conditions (day 1). An increase in cell proliferation was observed in all three groups during the first week of culture, with hyperoxic and normoxic ASCs showing higher expansion rates than in hypoxic ASCs. Thereafter, a substantial decrease in proliferation rate was observed in hyperoxic cultures, which was significant on day 11 (p = 0.032) (Fig. 4). Morphological observations exhibited two different behaviors according to oxygen abundance in repeated experiments (Supplementary Fig. S1).

Adhesion and proliferation of ASCs grown under different O2 concentrations. Human ASCs derived from five donors were incubated under hyperoxia (50–60% O2), normoxia (21% O2), or hypoxia (5–7.5% O2) and tested for adhesion (day 1) and proliferation (days 4, 8, 11, and 15) using the AlamarBlue. Results are presented as mean ± SD, *p < 0.05. Color images are available online.
ASC migration under different O2 levels
ASC migration, as measured using a scratch test, was accelerated in hyperoxia cultures, particularly after 48 h as compared with the other test conditions. Importantly, 72 h after inflicting the scratch, hyperoxic plates were fully occupied by ASCs (Fig. 5A). Quantitative analysis of the scratch area confirmed that ASC migration was significantly highest under hyperoxic as compared with normoxic conditions 48 h after the injury (p = 0.044) (Fig. 5B).

ASC migration under different O2 levels.
ASC osteogenic differentiation under hyperoxia, normoxia, and hypoxia
ASCs cultured in osteogenic medium were analyzed for osteogenic differentiation by ARS staining of calcium precipitates. No deposition of a calcified matrix was detected by day 7 in any of the cultures. Early mineralization was first observed after 14 days in osteogenic medium, with the most intense mineralization observed in the hypoxic group. By day 21, extensive matrix calcification was observed in all osteogenic cultures (Fig. 6A). Quantitative analysis of ARS staining demonstrated that hypoxic ASCs cultured in the osteogenic medium showed higher rates of cell differentiation throughout the entire experiment than the other tested O2 environments, with significantly higher differentiation capacity than normoxic ASCs measured on day 21 (p = 0.023) (Fig. 6B).
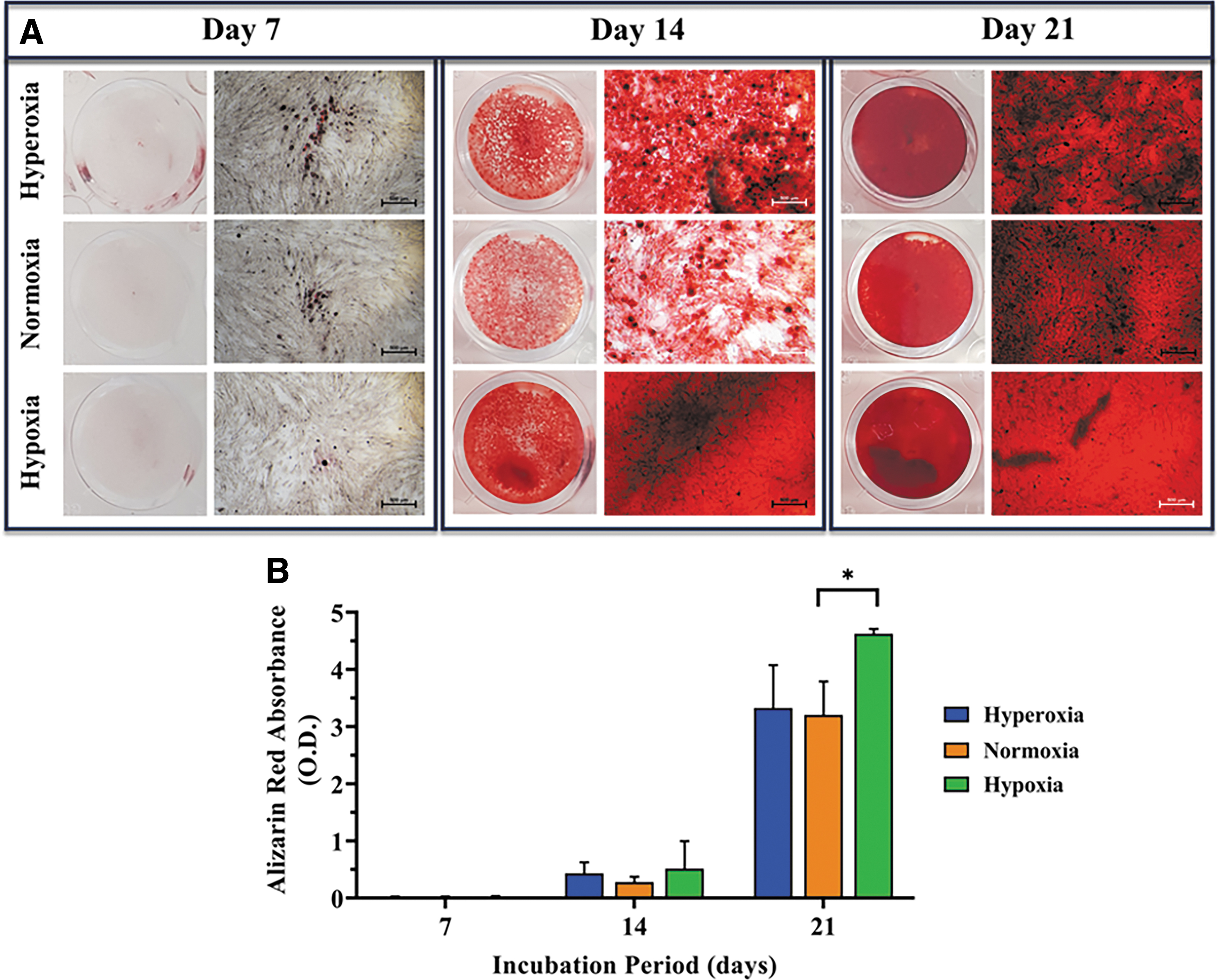
Effect of hyperoxia, normoxia, and hypoxia on ASC osteogenic differentiation. Human ASCs were cultured in osteogenic medium in the multichamber gas system and stained with ARS for detection of calcium precipitates.
Effect of hyperoxia, normoxia, and hypoxia on ASC angiogenic differentiation
Cells cultured in EBM-2 for 1 week and imaged immediately (0 h) after seeding on Matrigel demonstrated similar features under all tested O2 environments; all cells appeared rounded, and lacked the characteristic spindle-like triangular morphology. During the subsequent 2 h, hypoxic ASCs spread randomly and began to form small and interconnected clusters. Normoxic ASCs also formed less prominent interconnections. Four hours after plating, prominent and highly distinguished tube formation was observed in hypoxic ASC cultures, which manifested as highly connected clusters, which gave rise to cell cords interconnected by nodes (Fig. 7A). Notably, this phenomenon was not detected in any of the other ASC populations. ASCs kept under hyperoxic conditions maintained a round morphology throughout the entire experiment, and expressed none of the morphological changes already mentioned (Fig. 7A). Quantitative analysis of the tube formation after 4 h demonstrated that the amount of capillary-like structures formed under hypoxia was significantly higher than that formed under both normoxic and hyperoxic conditions (p = 0.0042 and p = 0.0007) (Fig. 7B). Evaluating hVEGF secretion levels suggested no significant differences among tested populations on both days 3 and 6 of culture (Fig. 7C).
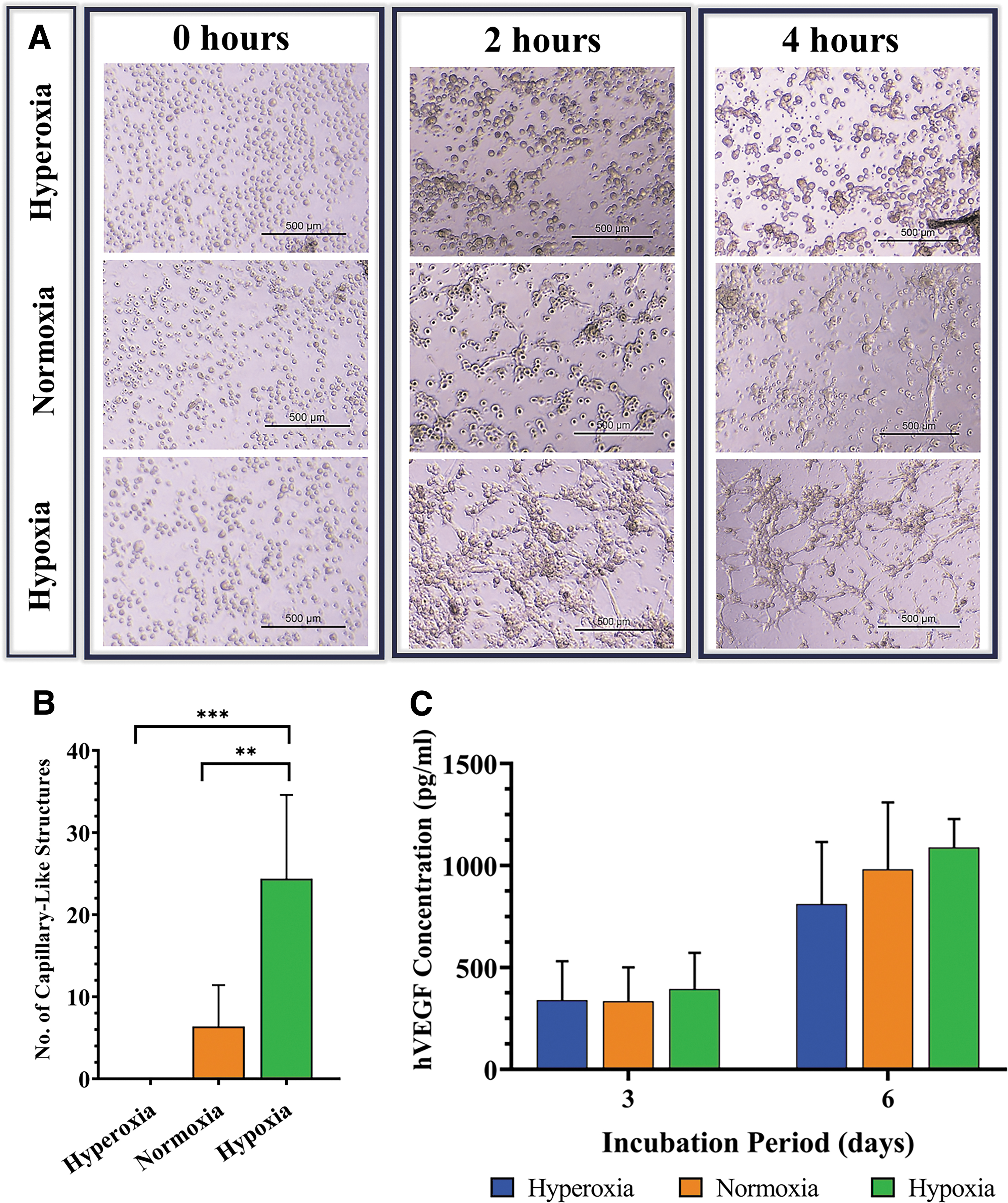
In vitro angiogenic evaluation of ASCs cultured under three O2 conditions.
Discussion
One of the major challenges of application of engineered bone tissue is to ensure successful integration of the implanted scaffolds and prevent graft necrosis, subsequent to insufficient vascularization and ischemia. This can be achieved by ensuring an appropriate O2 level in the cell niche, which, in turn, maintains the cell proliferation and osteogenic and angiogenic differentiation necessary for successful bone regeneration. In this study, two main strategies were suggested to overcome O2 deprivation in the implanted construct, namely, exposure of cell cultures, graft, or scaffold to hyperoxic conditions before implantation, or acclimation of the cells to reduced O2 (hypoxic) conditions. Hyperoxia is used in HBO therapy, which is proved to promote neovascularization, angiogenesis, and tissue regeneration. 14 Also, in vivo hyperoxygenation studies have showed positive effects.29,30 In contrast, the hypoxic atmosphere mimics both the physiological niche of ASCs 15 and the inner part of the ischemic construct, therefore, might be a suitable niche for cells to adapt to the hypoxic state.
The multichamber gas system was developed to investigate these proposed approaches. The novel system enables simultaneous testing of three different O2 atmospheres in three isolated chambers within a single incubator. According to our knowledge, despite the many attempts made to provide different O2 concentrations for cell culture, no efficient approach simultaneously testing three different O2 levels has been reported. Moving forward, it was important to harness this method to our needs. Therefore, our system was used to simultaneously investigate the effect of hyperoxia, hypoxia, and normoxia on hASCs in vitro characteristics: immunophenotype, attachment, proliferation, and osteogenic and angiogenic differentiation.
Flow cytometry analyses found no significant immunophenotypic differences between ASCs cultured under different O2 levels. However, quantitative examination of the ASC surface markers revealed significantly higher levels of both CD73 and CD90 on hyperoxia-cultured ASCs, whereas lower levels of CD34 were found on hypoxic ASCs. CD73 is a surface marker described as a mediator of cell adhesion and migration, 31 which correlates with our finding that hyperoxia significantly enhanced ASC migration. Of note, the enhanced migration response observed under hyperoxic conditions did not appear to result from increased viability or proliferation of cells, as elevation in proliferation of ASCs was not seen under such conditions. Importantly, CD90 has also been associated with enhanced cell migration. 32 The reduction in CD90 expression in hypoxic ASCs was associated with increased differentiation of MSCs, which aligns with its suggested role as an obstacle in the pathway of differentiation commitment. 33 Importantly, CD34 is considered a progenitor marker that correlates with cell plasticity; hence, loss of CD34 was associated with differentiation. 34 In this study, hypoxic ASCs displayed a higher differentiation capacity, combined with a significant decrease in both CD34 and CD90 levels, which aligns with the known roles of these markers.
In parallel, ASC proliferation in long-term cultures was enhanced in a reduced O2 environment, whereas their proliferative capacity in a hyperoxic atmosphere was significantly reduced on day 11. The present findings that cell proliferation is inhibited by hyperoxia, yet enhanced by hypoxia, are in partial agreement with other studies. 25 Grayson et al. and Kwon et al. reported that hMSCs displayed significantly improved expansion rates under hypoxia (2% and 5% O2, respectively).24,35 Conversely, Wang et al. and Antebi et al. showed contrasting results.23,36 These contradictory results can be explained by the variations in the experimental setup, such as the use of MSCs from different species and sources, at different stages of maturity, different culture conditions, and, more importantly, subtle differences in the O2 concentrations. 21
This study demonstrated that the osteogenic differentiation of ASCs was significantly enhanced during the 21 days in osteogenic medium under hypoxic conditions as presented by Kwon et al. 35 However, the hypoxic environment used in various reports varied from extreme hypoxia (1% or 2% O2) to 3% or 5% O2, which has seemingly conflicting reports on the impact of O2 on osteogenic differentiation.11,22,25 Kwon et al. reported that 5% hypoxia significantly enhanced osteogenic differentiation of hMSC. 35 In contrast, severe hypoxia of 1% decreased hMSC differentiation, whereas elevation of O2 levels to 3% restored osteogenic differentiation. 21 According to these studies, the precise O2 concentration seems to be a critical determinant of cell quiescence. Regarding ASCs angiogenic differentiation, ASCs promote new vessel formation by differentiating into vascular endothelial cells and secreting the VEGF. 37 Accordingly, our analysis demonstrated that hypoxia exhibited significantly higher tube formation capacity and may be enhancing hVEGF secretion. These findings closely align with other studies that demonstrated that angiogenesis is controlled by the secretion of growth factors and chemokines secreted by the hypoxic tissue, 38 particularly VEGF, which is considered the most potent inducer of blood vessel growth. 39
However, the ASCs used in this study were extracted from human adipose tissue of patients undergoing plastic surgery, which considerably limited the number of available donors for each experiment. In addition, sparse data exist, if any, on the effect of direct hyperoxia (>21% O2) on cell culture in general, and on ASC characteristics, in particular. Some recent studies examining the effect of in vivo hyperoxygenation have showed positive effects29,30; thus, further investigation of the impact of high O2 concentrations on ASC may be challenging, yet essential.
Through the multichamber gas system, this study identified several favorable effects of hypoxic exposure on in vitro ASC characteristics, which may be of importance when such cells are to be used for bone repair applications. Furthermore, this custom-made system possesses several unique features, including tight control of the gas pressure during infusion and complete removal of atmospheric air before each gas infusion. In addition, it can guarantee the required range of O2 concentration in each chamber. Furthermore, all three chambers are incubated under the same temperature and humidity conditions, so different assays can be performed concurrently in the same incubator, being a critical advantage. Importantly, the fabricated chambers are low cost, easily sterilizable, unlikely to create a pressurized environment during their use, and highly compliant with all culture plates, dishes, and flask sizes. Therefore, future studies testing additional O2 concentrations would be required to evaluate the efficiency of this system and determine the ideal O2 concentration for bone regenerations.
Conclusion
The innovative multichamber gas system overcame the lack of consensus of previous studies by enabling performance of all assays under consistent conditions, which yielded reproducible and comparable results. The system is cost-effective, easy to use, and applicable with any O2 level required. Future efforts that harness this method upon other cell cultures, grafts, or scaffolds under both in vitro and in vivo experimentation may provide new perspectives in the field of tissue engineering and regenerative medicine.
Footnotes
Authors’ Contributions
S.K. planned and performed the experiments, processed the experimental data, designed the figures, and interpreted the results. Moreover, she took the lead in writing the article. T.H.K. contributed to the study design and helped in data analysis and interpretation. In addition, she drafted the article. E.P. contributed to the conception of the research and the design of the experimental system. Moreover, he reviewed the article. S.S. supervised the experiment plan and the findings of this study. Furthermore, he contributed to the interpretation of the results and revised the article.
Acknowledgment
The authors gratefully thank Dr. Leonid Kogan for providing the adipose tissue specimens used in this study.
Disclosure Statement
None of the authors of this article has any competing interests to declare; therefore, no competing financial interests exist for any of the authors.
Funding Information
The authors received no specific funding for this study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.