
Abstract
Immune cells and their soluble factors have an important role in the bone healing process. Modulation of the immune response, therefore, offers a potential strategy to enhance bone formation. To investigate the influence of the immune system on osteogenesis, we developed and applied an in vitro model that incorporates both innate and adaptive immune cells. Human peripheral blood mononuclear cells (PBMCs) were isolated and cultured for 24 h and subsequently stimulated with immune-modulatory agents; C-class CpG oligodeoxynucleotide (CpG ODN C), polyinosinic acid–polycytidylic acid [Poly(I:C)], and lipopolysaccharide (LPS); all pathogen recognition receptor agonists, that target Toll-like receptors (TLRs) 9, 3, and 4, respectively. The conditioned medium (CM) obtained from PBMCs after 24 h was used to investigate its effects on the metabolic activity and osteogenic differentiation capacity of human bone marrow–derived mesenchymal stromal cells (MSCs). Conditioned media from unstimulated PBMCs did not affect the metabolic activity and osteogenic differentiation capacity of MSCs. The CM from CpG ODN C and LPS-stimulated PBMCs increased alkaline phosphatase activity (ALP) of MSCs by approximately threefold as compared with the unstimulated control, whereas Poly(I:C) CM did not enhance ALP activity of MSCs. Moreover, direct stimulation of MSCs with the immune-modulatory stimuli did not result in increased ALP. These results demonstrate that soluble factors present in CM from PBMCs stimulated with immune-modulatory factors enhance osteogenesis of MSCs. This in vitro model can serve as a tool in screening immune-modulatory stimulants from a broad variety of immune cells for (indirect) effects on osteogenesis and also to identify soluble factors from multiple immune cell types that may modulate bone healing.
Impact statement
In this study, an in vitro model is introduced that consists of multiple immune cell types from peripheral blood mononuclear cells and evaluates the effect of their secretome on mesenchymal stromal cell (MSC) osteogenesis after stimulation with pathogen recognition receptors ligands. This model can be used to develop a better understanding of the crosstalk between immune cells and bone-forming MSCs, paving the way to improve strategies for bone healing.
Introduction
Bone tissue has an intrinsic capacity to regenerate itself upon injury. However, the healing process of bone can be impaired under conditions like severe trauma, cancer (treatment), metabolic diseases, or mechanical instability.1,2 Transplantation of autologous bone to the defect site is currently the gold standard despite its limitations like increased procedure time 3 and limited availability. 4 Various synthetic grafts, particularly ceramics, hold the potential to be used as bone substitutes owing to their biocompatibility and osteoconductive properties. However, they generally lack bone-inducing capacity limiting their use as stand-alone grafts to treat large bone defects. 5 Even if ceramics perform as well as autografts, there still is a need for improvement to address the most challenging conditions such as spinal fusion.6,7 Strategies involving the use of cells, biomaterials, and/or growth factors8,9 are currently explored as a way to enhance the performance of synthetic grafts.
Owing to their close relationship with osteoblasts and bone deposition in the body, human multipotent mesenchymal stromal cells (MSCs) are the most common cells used to study the in vitro osteogenic potential of bone grafts and therapeutics. 10 Studies have shown that MSCs are capable of differentiation into osteoblasts, the effectors of bone formation in vivo, hence many strategies to augment bone regeneration focus on increasing the osteogenic potential of MSCs. 11 However, therapeutic approaches targeting only MSCs usually fail when translating their application to in vivo settings, indicating that the mechanisms underlying bone formation are still poorly understood. 10 For this reason, several new strategies to achieve predictable and enhanced bone formation in vivo are under development, and osteo-immunomodulation is a promising candidate. Modulation of the local immune response as a strategy to enhance bone formation has gained a lot of interest, considering the emerging recognition of the pivotal role of a balanced early inflammatory phase in the bone healing process. 12
Research showed that administration of bacterial stimuli leads to increased bone formation in a rabbit tibia model. 13 The rationale behind this phenomenon is believed to be the release of cytokines upon the activation of pathogen recognition receptors (PRRs) in immune cells and bone cells. Recently, it was shown that specific targeting of PRRs can be harnessed to promote osteogenesis through immune-modulation. 14 PRRs are single-pass transmembrane proteins with an extracellular region (involved in recognition of the pathogens), and an intracellular region (involved in signal transduction). They are highly expressed in immune cells, and, upon stimulation, PRRs trigger the release of cytokines and soluble factors. 15 Hence, the stimulation of PRR ligands to trigger the release of pro-osteogenic cytokines represents a valid therapeutic strategy to promote augmented osteogenesis in MSCs through the immune system.
Taking cues from the fracture healing process as an example of efficient bone regeneration, the crosstalk between the immune cells and bone cells or their precursors (MSCs) is a prerequisite for optimal bone healing. 16 Both innate and adaptive immune cells can be found at the fracture site during the early stage after fracture, which indicates that inflammation is the first action of repair.17,18 After the injury, proinflammatory cytokines such as tumor necrosis factor (TNF-α), interleukin-1 (IL-1), IL-6, IL-17, and anti-inflammatory cytokines such as IL-10, IL-4 act in synergy with bone morphogenetic proteins (BMPs) and other transforming growth factors to facilitate infiltration of immune cells, regulate angiogenesis, and induce migration and differentiation of bone progenitor cells (MSCs) at the site of injury.19,20 Studies using knockout mice have demonstrated the functional role of these immune cells and related inflammatory factors during fracture healing. For example, the selective depletion of macrophages, dendritic cells, and T, and B cells at different time points after the fracture has confirmed their contribution to the fracture healing process.21–23
The innate and adaptive immune cells secrete cytokines and growth factors that favor osteogenesis and angiogenesis. For example, it was demonstrated that administrating the conditioned medium (CM) obtained by stimulating human macrophages with lipopolysaccharide (LPS) leads to increased alkaline phosphatase activity (ALP) in MSCs, indicating the presence of soluble pro-osteogenic factors in the medium. 24 Moreover, cytokines secreted by macrophages have been shown to induce signal transducer and activator of transcription 3 (STAT3) signaling in MSCs leading to upregulation of osteoblast-related genes, such as ALP and Runt-related transcription factor 2 (Runx2). 25 Similarly, we have shown that soluble factors present in antibody-CD3/CD28-activated T cell-derived CM led to an increase in MSC osteogenic differentiation. 26 These examples underline that both the innate and adaptive immune cells actively participate in the processes underlying bone regeneration, but the precise mechanisms are still poorly understood. The optimal inflammatory milieu needed to promote osteogenesis in MSCs is still elusive.
In this study, we introduce a model for the in vitro evaluation of all blood-resident immune cell types simultaneously to discover the inflammatory milieu needed to promote bone formation. To this end, investigating peripheral blood mononuclear cells (PBMCs), which include circulating immune cells, represents a valid approach. 27 PBMCs are mononuclear cells consisting of lymphocytes (T cells, B cells, and natural killer cells) and monocytes, the precursors of macrophages. Because PBMCs can be easily collected, they are currently an attractive source of immune cells for molecular and immunologic studies. However, studies utilizing whole PBMCs in in vitro models to screen potential osteo-immunomodulators and their effects on the secretome of immune cells for bone regeneration are still lacking. The in vitro model that we herewith introduced consists of both innate and adaptive immune cells from PBMCs and we investigate the effects of their indirect secretory factors on MSC differentiation into the osteogenic lineage. This model may be used as a screening tool for osteo-immunomodulation to improve bone healing.
Methods
In vitro model set up
Human PBMCs were isolated from the blood of five healthy donors using the density gradient centrifugation method. The PBMCs were seeded in 24-well tissue culture plates and cultured in PBMC culture medium in the presence and/or absence of immunomodulatory factors (PRR ligands). After 24 h, the supernatant was collected and centrifuged at 1500 g for 5 min to pellet the cells. The cell-deprived supernatant, referred to as the CM, was then stored at −80°C. The CM was analyzed using a cytokine array kit for quantifying the secreted soluble factors. Furthermore, the effect of the CM was investigated on MSC osteogenic differentiation (Fig. 1).

In vitro model set up for investigating the response of immune-modulatory PRR ligands on osteogenesis in vitro. PBMCs were isolated from human blood and stimulated with immunomodulatory PRR ligands for 24 h. Following incubation, the CM was used to investigate its effects on MSCs. The soluble immune cell products present in the CM were analysed using a cytokine array. Image has been created using
Reagents
C-class CpG oligodeoxynucleotide (CpG ODN C) (M362) and high molecular weight polyinosinic acid–polycytidylic acid [Poly(I:C)] were purchased from InvivoGen (San Diego, CA). LPS (O55:B5, produced from E. coli) was purchased from Sigma Aldrich (St. Louis, MO). The concentration range was based on the manufacturer's data sheet and literature. 14
Isolation of mononuclear cells from human peripheral blood and generation of CM
Twenty milliliters of blood from healthy donors (n = 5) was obtained with informed consent from the blood bank (Sanquin, Utrecht, the Netherlands) because ∼15 million cells were required for the experiments. The heparinized blood from individual donors was diluted 1:1 with phosphate-buffered saline (PBS) solution and layered on top of a 15 mL density gradient medium (Ficoll-plaque plus, Cytiva, MA) in SepMate PBMC tubes (Stem Cell Technologies, Cologne, Germany). These tubes were then centrifuged at 1200 g for 10 min with the brake on at room temperature. Following centrifugation, the supernatant containing serum and cells was quickly poured off into a new centrifuge tube and centrifuged at 300 g for 8 min to obtain a cell pellet. PBMCs were then suspended in a culture medium consisting of RPMI-1640 glutamax (Thermo Fisher Scientific, Waltham, MA) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS) and 100 U/mL penicillin and 100 μg/mL streptomycin.
To produce 1 mL of the CM, PBMCs obtained from individual donors were seeded in 24-well tissue culture plate at a density of 500,000 cells/cm2 and cultured for 24 h at 37°C in a humidified atmosphere containing 5% CO2. Subsequently, PBMCs were stimulated with different PRR ligands (Table 1). Control experiments were performed in the absence of PRR ligands. After 24 h of culture, the CM was collected, centrifuged at 1500 g for 5 min, and stored at −80°C until use.
Overview of the Tested Pathogen Recognition Receptor Ligands and the Toll-Like Receptors That They Target
CpG ODN C, C-class CpG oligodeoxynucleotide; LPS, lipopolysaccharide; Poly(I:C), polyinosinic acid-polycytidylic acid; PRR, pathogen recognition receptor; TLR, toll-like receptor.
Isolation and culturing of MSCs
Human MSCs were isolated from bone marrow aspirates obtained from four subjects 50–80 years old that underwent orthopedic surgery for knee or hip reconstruction and provided written consent for this procedure, for which approval was provided by the UMC Utrecht medical ethical committee under the protocols METC 08-001/K and METC 07-125/C. MSCs were isolated and cryopreserved after passage 2 as previously described in detail. 28 This standardized method of isolation yields multipotent cells as confirmed with assays into osteogenic, adipogenic, and chondrogenic lineages.
For the experiment, MSCs were thawed and expanded in an expansion medium (EM) consisting of minimum essential medium (α-MEM [Gibco]) supplemented with 10% (v/v) heat-inactivated FBS and 100 U/mL penicillin and 100 μg/mL streptomycin and 0.2 mM
Metabolic activity assay of MSCs
The metabolic activity of the MSCs was determined using the Alamar blue assay, a nondestructive assay that allows quantification of metabolism products that reflect expansion and/or activity. Alamar blue solution was prepared by dissolving 5.54 mg of Resazurin sodium salt (R7017; Sigma Aldrich) in 50 mL PBS (10 × ) and sterile filtered. On days 1, 3, and 7 of MSC culture with CM and controls, the media of all the wells were replaced by the Alamar blue solution that was diluted in the MSC EM. After 2 h of incubation at 37°C in a humidified atmosphere containing 5% CO2, the color change in the media was measured using an absorbance plate reader (Clariostar plus; BMG Labtech, Germany) at 570 nm and corrected at 600 nm.
Cell count and osteogenic differentiation assay of MSCs
Following 7 days of culture, cells were lysed with 0.2% (v/v) Triton-X 100/PBS for 30 min at room temperature. ALP was determined by using the hydrolysis reaction of p-nitrophenyl phosphate (pH = 9.6) (SigmaFast Pnpp tablets; Sigma Aldrich). The absorbance was measured at 405 nm using an absorbance plate reader and corrected at 655 nm. ALP was normalized to the DNA content in the wells. DNA quantification was performed using Quanti-kit (Invitrogen, MA) according to the manufacturer's instructions.
Experiments
Dose determination of CM for MSC culture
To determine an appropriate dose of the CM that should be added to the MSCs, the CM from unstimulated PBMCs (n = 3) was added in different ratios to the MSC culture. The metabolic activity, cell count, and osteogenic differentiation assays were used as readout parameters.
To determine the effect of the CM on metabolic activity, MSCs (n = 3) were seeded and cultured in an EM at a density of 15000 cells/cm2 in 96-well tissue culture-treated plates. Upon confluency, the CM obtained from unstimulated PBMCs was added to the wells of the EM (CM: EM) in the ratios 1:10, 1:4, and 1:1 for 7 days. Wells with EM were used as control. The medium was refreshed every 3 days.
To test the effect of the CM on osteogenic differentiation, MSCs (n = 3) were similarly seeded and cultured. Upon confluency, the medium was replaced with an osteogenic differentiation medium (ODM) (EM supplemented with 10 mM β-glycerophosphate and 10 nM dexamethasone) alone or combined with the CM obtained from unstimulated PBMCs. The CM to ODM (CM: ODM) in ratios 1:10, 1:4, and 1:1 was added every 3 days.
MSCs osteogenic differentiation following indirect stimulation of PRR ligands
MSCs (n = 4) were stimulated continuously with the CM obtained by stimulating the PBMCs with the PRR ligands (Table 1) for 24 h. The PRR ligand concentration that yielded high ALP activity in MSCs for osteogenic differentiation, upon indirect stimulation was chosen for further experiments (Supplementary Fig. S1). The final concentrations used were CpG ODN C (1 μg/mL), Poly(I:C) (10 μg/mL), and LPS (1 μg/mL). Because the effect of the CM obtained after the stimulation with PRR ligands from five PBMC donors was similar in terms of osteogenic differentiation of MSCs (Supplementary Fig. S2), CM from one PBMC donor was used. The MSCs cultured in standard osteogenic medium alone or combined with CM from unstimulated PBMCs were used as controls. The CM obtained from PRR stimulation was not supplemented with any additional amounts of PRR ligands for further analysis. All MSC experiments were performed with the ODM described previously for 7 days, with the medium refreshed every 3 days. After 7 days, the ALP activity and DNA content were measured as described previously.
Direct stimulation of PRR ligands on MSCs osteogenic differentiation
Because the PRR ligands was shown to enhance osteogenic differentiation of MSCs upon direct stimulation, 14 this experiment was performed as a control for indirect stimulation experiments with effects of PRR ligands using the CM from PBMCs. The MSCs (n = 4) were also stimulated with CpG ODN C (1 μg/mL), Poly(I:C) (10 μg/mL), and LPS (1 μg/mL) directly and cultured in ODM for 7 days. After 7 days, the ALP activity and DNA content were measured as described previously.
CM characterisation using cytokine array
To identify cytokines, chemokines, and growth factors present in the CM after stimulation with PRR ligands in one PBMC donor, a cytokine array assay (GSH-BMA-1000; RayBiotech, GA) was performed to determine a panel of inflammatory mediators. The array included 41 proteins. Pro-inflammatory cytokines and chemokines TNF-α, interleukins (IL-1α, IL-1β, IL-6, IL-8, IL-11, IL-17A), monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-1a (MIP-1α) along with bone-related proteins like BMPs (BMP-2, BMP-4, BMP-6, BMP-7, BMP-9), osteoactivin, osteopontin, osteoprotegerin DKK-1, activin A, FGF-1, amphiregulin, bFGF, E-selectin, CD54, IGF-1, matrix metalloproteinase 2 (MMP-2), MMP-9, MMP-13, P-cadherin, RANK, SDF-1alpha, sonic hedgehog N-terminal, TGF-β1, TGF-β2, CD106, CDH5, DKK-1, MMP-3, TRANCE, PDGF-BB, and TGF-β3 were analyzed according to manufacturer instructions.
In brief, the glass slide was first blocked with the sample diluent for 30 min to prevent unspecific binding. Later, 100 μL of the CM obtained from unstimulated PBMCs and PRR ligand-stimulated PBMCs was added to different wells in the array slide and incubated for 2 h at room temperature. Following incubation, the array was washed three times for 5 min each. Subsequently, a biotinylated antibody cocktail was added to the wells of the array and incubated for 2 h at room temperature. Following washing, the array was incubated with Cy3 equivalent conjugated streptavidin for 1 h. The signals were then visualized using a fluorescence laser scanner at a PMT gain of 50%. Data analysis was performed using R-data software.
To show the results, a ratio average (RA) plot was used. Essentially, the RA plot is a scatter plot that shows the differential expression of a cytokine in group “a” versus group “b” according to their ratio (R = log2(a/b)), and average (A = 1/2log2(a*b)). In particular, “a” is the positive measure in the group of interest (here the medium of the PRR stimulated PBMCs), whereas “b” is the positive measure in the control group (here the medium of unstimulated PBMCs). Thus, overexpressed cytokines have R > 0, whereas downregulated cytokines have R < 0. A threshold of a twofold increase was applied to identify differences.
Statistical analysis
Statistical analysis was performed. The analysis was carried out using R studio software (version 1.3.1093) and Prism Graphpad (San Diego, CA). The normality of the data was checked using the Pearson normality test and Q-Q plots. If the data were normally distributed, ordinary one-way analysis of variance (ANOVA) was performed and if data were not normally distributed, a Wilcoxon signed-rank test was used to determine significant differences between the groups.
Results
Dose determination of the CM from unstimulated PBMCs for MSCs culture
The addition of the CM from unstimulated PBMCs in different dose ratios (CM: EM) did not affect the metabolism of the MSCs on days 1 and 7 for all three donors (Fig. 2a). On day 3, a 50% increase in metabolic activity was observed for one of the three donors in the groups treated with the CM. This increase may be owing to an increase in cell amount or because of high activity of cells, or a combination of both.
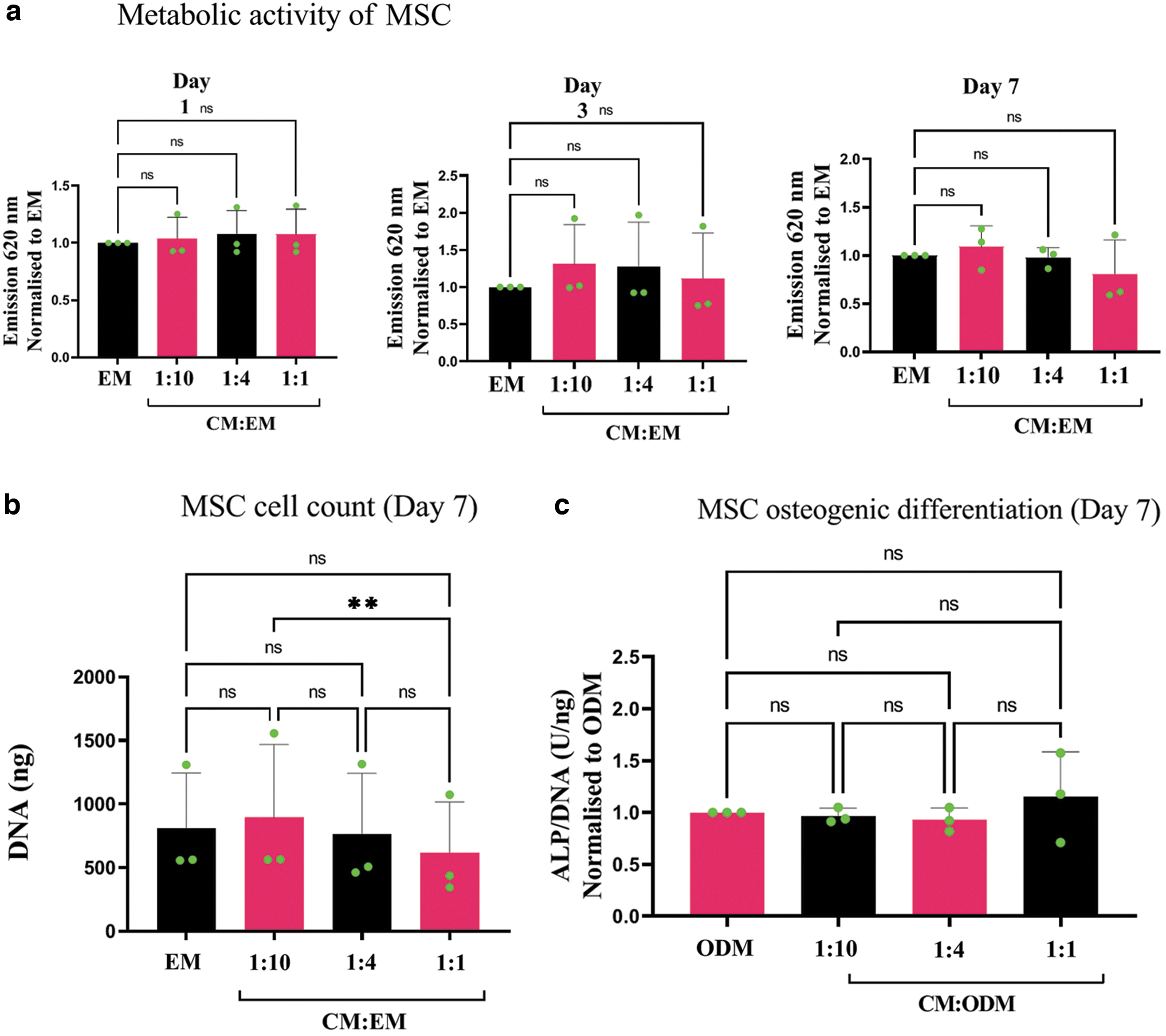
Effect of CM of unstimulated PBMCs (n = 3) on the metabolic activity, cell count, and osteogenic differentiation of MSCs (n = 3).
The CM from unstimulated PBMCs significantly affected the cell count, except in the case of the highest dose in which the CM equaled the osteogenic medium (1:1) (Fig. 2b). Furthermore, the CM did not interfere with the MSC's osteogenic differentiation capacity (Fig. 2c), according to the alkaline phosphatase assay. In particular based on the latter results (ALP/DNA), we choose to conduct all further experiments with the ratio of 1:4 for CM to ODM. In addition, the CM from unstimulated PBMCs did not promote the osteogenic differentiation of MSCs in the absence of dexamethasone (Supplementary Fig. S3).
Effect of direct versus indirect stimulation of PRR ligands on osteogenic differentiation
Upon direct stimulation (PRR ligands directly with the osteogenic medium of MSCs), both CpG ODN C and LPS enhanced the ALP activity in MSCs by approximately twofold as compared with CM from unstimulated PBMCs (control CM) (Fig. 3). Of interest, the CM obtained from CpG ODN C stimulated PBMCs (indirect stimulation) induced even higher ALP activity (increase in donors ranging from 10% to 145%, p = 0.06) as compared with direct stimulation with CpG ODN C.

Alkaline phosphatase levels in MSC cultures, corrected for DNA, after addition of PRR ligands to ODM. MSC stimulation with ligands was either direct (grey) or indirect (pink) via PBMC (n = 1) stimulation where unstimulated PBMC conditioned media (control CM) served as control situation for normalization. Immunomodulatory PRR ligands CpG ODN C (1 μg/mL), Poly(I:C) (10 μg/mL), and LPS (1 μg/mL) were given either directly to the MSCs (with ODM) or used to stimulate the PBMCs, from which the CM was subsequently provided to the MSCs. The CM was added in the ratio of 1: 4 (CM: ODM). The graph represents the mean ± SD of technical triplicates performed per donor for four individual MSC donors. Significance was tested using Wilcoxon signed-rank test, one-tailed. CpG ODN C, C-class CpG oligodeoxynucleotide; LPS, lipopolysaccharide; Poly(I:C), polyinosinic acid-polycytidylic acid.
CM characterization using cytokine array
The cytokine array analysis was only performed for CM obtained from CpG ODN C as high ALP activity was seen because of this form of indirect stimulation. CM from LPS-stimulated PBMCs was used as a positive control because LPS is known to activate the nuclear factor kappa B (NF-kB) signaling pathway leading to the release of proinflammatory cytokines. Stimulation with CpG ODN C resulted in the release of proinflammatory cytokines IL-8 (threefold) and chemokine MCP-1 (fourfold), when compared with unstimulated PBMCs (Fig. 4b). A twofold increase in MMP-9 and MMP-13 was observed in the CpG ODN C-stimulated CM as compared with control CM. Upon LPS stimulation, the release of proinflammatory cytokines IL-6, IL-8, TNF-α, and IL-1β were seen along with chemokines MCP-1 and MIP-1α (Fig. 4c).

Cytokine production by PBMCs upon stimulation with PRR ligands CpG ODN C and LPS after 24 h.
Discussion
Although the modulation of immune responses may be a way to enhance bone repair, an understanding of the optimum inflammatory environment to achieve this in vivo remains elusive. With the present work, we provide an in vitro model that employs PBMCs isolated from peripheral blood, which includes a variety of immune cells from both the innate and adaptive immune systems, to study their effects on MSC osteogenesis. Herewith, we show that using the CM obtained from unstimulated PBMCs does not affect MSCs in terms of metabolic activity or osteogenic differentiation.
We further applied this model to screen two immunomodulatory PRR ligands that may possess pro-osteogenic effects. Upon stimulation with CpG ODN C and LPS, directly or via a CM obtained by stimulating PBMCs, MSCs showed an increase in ALP activity as compared to controls with only osteogenic medium, whereas stimulation with Poly(I:C) directly or via the CM did not show an increase in ALP activity of MSCs. Moreover, the ALP activity was higher in the CpG ODN C stimulated CM group (indirect stimulation), indicating that the soluble factors secreted from PBMCs contributed to the pro-osteogenic effects of MSCs rather than the CpG ODN C itself (direct stimulation).
Guided by these observations, we investigated which cytokines and growth factors were present in the PBMC-derived CM upon stimulation with CpG ODN C. To do so, a cytokine array was performed on the conditioned media to quantify the release of multiple (most inflammatory) soluble factors simultaneously. Interestingly, we found that inflammatory mediators, mainly chemokines IL-8 and MCP-1 were upregulated upon stimulation with CpG ODN C. As previously described by others, MCP-1, also referred to as CCL-2, is an important chemokine involved in bone regeneration, since knock-out models for MCP-1 showed reduced bone formation.32,33 This may be attributed to MCP-1 attracting monocytes and dendritic cells to locations of injury; the bone defect zone. These monocytes can further develop into macrophages and produce more MCP-1, with subsequent more monocyte attraction. As such the monocyte-macrophage feedback loop can be considered the key player in bone regeneration. Along with the chemokines, an upregulation in MMP-9 and MMP-13 was detected in CpG ODN C stimulated CM. MMPs are known to degrade the extracellular matrix and play a vital role in bone remodelling and skeletal repair. 34 It was shown that MMP-9 plays a crucial role in regulating the chondrogenic and osteogenic differentiation in early phases of fracture healing. 35 It was also shown that fractured tibiae of MMP-9 knockout mice developed non-unions. 36 Growth factors BMP-2 and BMP-6 were also secreted by PBMCs upon stimulation with CpG ODN C. Hence, although this cytokine array experiment was performed for one donor only, we found similarities with previous in vivo observations, indicating the suitability of our model to screen for pro-osteogenic factors.
To confirm the reliability of our model, we screened for inflammatory mediators present in the conditioned media upon stimulation with LPS; a known stimulator of inflammatory cytokine production and for bone regeneration in vitro. 19 As expected, the analysis revealed high expression of pro-inflammatory cytokines including IL-6, IL-1β, and TNF-α 37 next to inducing TGF-β1, TGF-β2 and BMP-9, respectively. Further, studies recreating the composition of the CM in terms of cytokines/chemokines in vitro can help in determining the exact role of the secretory cytokines and chemokines in maintaining the balanced inflammatory microenvironment created in vivo during bone healing process.
In the past decade, the effects of individual immune cell subpopulations during the initial phase of fracture healing have been studied extensively. At the fracture site, macrophages secrete cytokines and chemokines that recruit not only monocytes (via MCP-1) but also direct MSCs to the site of injury from their local niches to start the rebuilding of new bone. Macrophages support osteoblast formation by release of growth factors like TGF-β1, BMP-2 and BMP-4 and thus stimulate MSCs toward the osteogenic lineage.18,25 Lymphocytes consisting of T- and B- cells, affect the differentiation and maturation of bone cells, other than their immune functionality and are present in fracture hematoma formed at the site of injury. Activated T-cells and their subsets can stimulate osteogenic differentiation of MSCs in vitro. 26 In contrast to that, fracture healing was enhanced in the mice which lacked functional B- and T- lymphocytes, implying that adaptive immune cells somehow have a negative impact.38,39 Further, it was reported that CD8+ T- cells, a subset of T lymphocytes impair the fracture healing process in humans and mice, 40 while CD4+ cells have been shown to enhance osteogenic differentiation. 41
Recently, it was shown that activated T cells promote B cells to produce osteoprotegerin, which has a promoting effect on bone tissue regeneration via the CD40/CD40L signalling pathway. 42 Though roles of individual immune cell subpopulations during fracture healing are characterized to a certain extent, there is a need for combined models of bone (progenitor) cells and various types of immune cells in vitro that recapitulate the cross talk, microenvironment, and key features of the bone healing process of the in vivo situation.
Our study identifies CpG ODN C as a potential osteo-immunomodulator. CpG ODN C is a synthetic bacterial DNA and currently investigated in clinical trials as an adjuvant for asthma and tumour therapies.43,44 Recently, we showed that CpG ODN C enhances MSCs osteogenic differentiation in presence of dexamethasone. 14 CpG ODN C is recognized by intracellular receptor toll-like receptor (TLR)9, which upon activation, initiates the NF-kB and TIR domain-containing adapter-inducing interferon β (TRIF) signalling pathway, which results in the release of pro-inflammatory cytokines like interferons, IL-8, IL-6.45,46 In agreement, we found upregulation in IL-6 and IL-8 secretion in the CpG ODN C-stimulated PBMC group suggesting the activation of the NF-kB signalling cascade.
In this study, antibodies targeting interferons were not included in the cytokine array. It has been shown that CpG ODN C stimulation in macrophages leads to the release of MCP-1 via the c-Jun N-terminal kinase signalling pathway, mainly due to the activation of TLR9. 47 Since CpG ODN C is reported to be a potent stimulator of B-cells, NK cells, and dendritic cells that induce secretion of immunoglobulins and interferon type 1 cytokines,48,49 it will be interesting to examine the presence of interferons in CpG ODN C stimulated CM. Thus, evaluating the contribution of the immune cells upon stimulation with CpG ODN C on MSC osteogenic differentiation represents an important step toward the use of CpG ODN C in enhancing bone repair. In the future, studies investigating the contribution of immune cell-secreted IL-8 and MCP-1 to bone formation should be carried out to provide a deeper insight into the mechanism benefitting osteogenesis.
As a limitation of this study, the evaluation of bone marker expression was limited to the investigation of ALP activity in MSCs, an early marker of osteogenesis. ALP activity is considered to be a superior predictor of bone formation in vivo 50 as compared to later osteogenic markers like osteocalcin, and osteonectin. The effect of the CM, stimulated with PRR ligands, should also be investigated on matrix mineralization, which is considered a functional endpoint assay for osteogenesis. Furthermore, characterization of the conditioned media obtained from stimuli like LPS, which are already investigated in detail, 37 in terms of its cytokine content by using a larger cytokine array or enzyme-linked immunosorbent assays could provide invaluable insights into the microenvironment that might be beneficial for developing future therapies.
Use of limited number of donors proved to be another limitation of this study. In our study the four donors used in the indirect stimulation experiments did show large differences (Fig. 3). However, donor variation may have an important effect on our outcomes and further investigation with multiple donors should be performed before a next step to in vivo experiments can start. Another limitation of this study lies in the fact that we use CM from the immune cells to stimulate MSCs rather than co-culturing of PBMCs and MSCs. The current model does not account for the effects of cell-to-cell contact on cytokine secretion. For instance, MSCs may have immunomodulatory effects on PBMCs that may suppress or stimulate the production of soluble mediators. 51
Nevertheless, the established in vitro model resembles the physiological inflammatory state created in vivo, and proves to be simplified practical tool that can be applied for various applications including the identification of new osteo-immunomodulators. Since it has been elucidated that both adaptive and innate immune cells are key players in the immune-mediated bone deposition, developing new platforms that allow screening of potential bone enhancing immune modulators is of key importance to accelerate their employment in the clinic. Herein, we described a model that allows such tests and may as well help to identify immunomodulatory biomaterials and thereby supports in vivo experiments that are still required to clinically test bone regenerative compounds and materials.
Conclusion
In conclusion, in this study, we provide evidence of the suitability of a new in vitro model based on PBMC-derived conditioned media produced upon stimulation with PRR ligands to study osteogenesis in MSCs. By culturing PBMCs in the presence of PRR ligands for 24 h, the production of cytokines and growth factors was induced as shown by the cytokine array. In particular, stimulating PBMCs with CpG ODN C, a well-known adjuvant in clinical trials, and administrating the CM from the stimulated PBMCs to MSCs led to increased alkaline phosphatase levels as an indicator of osteogenic differentiation. No effects were observed when MSCs were cultured in the presence of conditioned media from unstimulated PBMCs, confirming the viability of this in vitro model. Hence, the model described allows the screening of PRR ligands for osteoimmunomodulation and represents a promising strategy to better mimic the in vivo situation where multiple immune cell types interact with osteo-immunomodulators.
Footnotes
Acknowledgment
The authors thank Dr. Margreet Wolfert, Department of Chemical biology and drug discovery, University Utrecht for her valuable help in reading the cytokine array using a fluorescent laser scanner.
Authors' Contributions
P.K., R.B., and M.C. designed and conducted the experiments, interpreted, and analyzed the data, and wrote the article. M.C.K. and H.W. secured the funding and contributed to the experiment design, data interpretation, and article review. D.G. contributed to data interpretation and article review. All authors have read and approved the final submitted article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This article is supported by PPS allowance from the Health∼Holland LSH-TKI (grant no.: LSHM18011) and the EU's H2020 research and innovation program under Marie S. Curie Cofund RESCUE (grant agreement no. 801540).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.