
Abstract
The state of host cells is reflected in the cargo carried by their extracellular vesicles (EVs). This makes EV a potential source of biomarkers for human diseases. Piwi-interacting RNA (piRNA) regulates gene expression through epigenetic regulation and post-transcriptional gene silencing. Thus, piRNA profiling in EVs derived from human clinical samples could identify markers that characterize disease stages and unveil their roles in disease pathology. This review aimed to report the expression profiles of EV-derived piRNA (EV-piRNA) in various human samples, as well as their role in each pathology. A systematic review was conducted to collate the findings of human EV-piRNA from original research articles published in indexed scientific journals up to February 16, 2022. Article searches were performed in PubMed, Web of Science, and Scopus databases, using a combination of keywords, including “EV” and “piRNA.” A total of 775 nonredundant original articles were identified. After subjecting articles to inclusion and exclusion criteria, 34 articles were accepted for this review. The piRNA expression levels among the small RNA profiles of human-derived EVs range from 0.09% to 43.84%, with the lowest expression level reported in urine-derived EVs and the highest percentage in plasma-derived EVs. Differentially expressed EV-piRNAs have been identified in patients with specific disease conditions compared to their counterparts (healthy control), suggesting an association between piRNA and progression in various diseases. Seven articles identified piRNA putative target genes and/or the pathway enrichment of piRNA target genes, and one study demonstrated a direct role of piRNA candidates in disease pathology. In conclusion, EV-piRNA has been isolated successfully from various human body fluids. EV-piRNA is a new research niche in human disease pathology. The expression profiles of EV-piRNA in various tissue types and disease conditions remain largely unexplored. Furthermore, there is currently a lack of guidelines on piRNA bioinformatics analysis, which could lead to inconsistent results and thus hinder the progression of piRNA discoveries. Finally, the lack of published scientific evidence on the role of EV-piRNA supports the need for future research to focus on the functional analysis of EV-piRNA as part of the route in piRNA discoveries.
Impact statement
This systematic review provides a detailed analysis of the extracellular vesicle-derived Piwi-interacting RNA (EV-piRNA) in various human samples that include the methods used for isolation and characterization of the EVs, sequencing platforms used, and bioinformatics analysis findings of the EV-piRNA expression profiles, as well as the distinct EV-piRNAs identified in various body fluids/clinical conditions. A summary of the EV-piRNAs derived from different samples is listed in this review, which can be used as a reference for future studies on the role of EV-piRNA in human disease.
Introduction
Extracellular vesicles (EVs) are membrane-bound vesicles (30–10,000 nm) that are released from cells into body fluid, such as blood, urine, and synovial fluid. They can be grouped into three subtypes, including exosomes (30–200 nm), microvesicles (100–1000 nm), and apoptotic bodies (>1000 nm), based on their size ranges and biogenesis pathways. Despite numerous studies on exosomes, distinguishing exosomes from other EV subtypes, such as microvesicles, due to overlapping size ranges and protein markers, remains challenging.
Exosomes arise by clathrin-dependent endocytosis, in which the cytosolic components of the donor cell are enclosed in a membrane-bound multivesicular body of late endosomes, and eventually fuse with the plasma membrane. 1 The vesicles are then released into the extracellular compartment through a process called budding. Identification of exosomes/EVs can utilize their biogenesis pathway. Membrane proteins and endosomal sorting cells required for transport (ESCRT)-related proteins are typically found on their surface or enclosed within exosomes, including tetraspanin family proteins (CD63, CD9, and CD81), Flotillin-1, and TSG101. 2 According to the International Society for Extracellular Vesicles (ISEV) guideline, EV will be used as the generic term in this review. 3
The in vitro and in vivo preclinical analyses of EVs' ability to transport cellular contents and regulate gene expression in their targeted cells have generated great interest in human disease studies. EVs carry various types of cargo, including proteins and nucleic acids, that reflect the nature/phenotype of donor cells, as well as their physiological and pathological state. Many RNA subtypes have been identified in EVs, including messenger RNA (mRNA), microRNA (miRNA), long noncoding RNA (lncRNA), transfer RNA (tRNA), small nuclear RNA (snRNA), as well as Piwi-interacting RNA (piRNA). 4 This underpins their ability to serve as a biomarker source for human disease. EVs can also mediate intercellular communication as they activate the signaling cascades in target cells upon binding to the transmembrane receptor. Moreover, EVs can fuse into recipient cells and release their cargo contents into the cytoplasmic compartment. 5
piRNA is a small noncoding RNA that can mediate gene silencing. It is 26–31 nucleotides in length, with 2′-O-methylation at the 3′ end. piRNA acts through interaction with Piwi protein subfamily of the ARGONAUTE protein family. 6 The vast majority of piRNAs are produced by specific genomic loci called piRNA clusters. piRNA clusters can be divided into two main groups: the uni-strand clusters, which are a widely distributed type, or dual-strand clusters, which are mainly detected in Drosophila germ cells.
Although mRNA has been the RNA subtype most frequently interrogated in studies of cellular behavior and pathophysiological conditions in vitro 7 or in vivo, 8 piRNA has also been investigated in cancer and other human diseases. Indeed, differentially expressed (DE) piRNAs have been detected between patients and controls, 9 suggesting that piRNA might play a role in disease progression. Early data indicated the functions of piRNAs to include transposon silencing and gene integrity maintenance in germline cells. More recently, piRNAs have been detected in both germline and somatic cells in many animal phyla, from insects and fish to mammals. 10 It is becoming apparent that piRNAs not only serve as transposon silencers but also they play many crucial roles in regulating gene expression. piRNA can mediate gene silencing in both the transcriptional and post-transcriptional levels, by interacting with DNA methyltransferases or histone-modifying enzymes or directly binding to complementary sequences in mRNAs, respectively. 11
To investigate if piRNA is expressed and stable in human body fluid-derived EVs, we have conducted a systematic review using original articles that characterized and analyzed the EV-derived piRNA (EV-piRNA) extracted from clinical body fluid samples. This systematic review aimed to interrogate the expression profiles of EV-piRNA in various human body fluids, as well as their potential role in each clinical condition.
Methods
This systematic review queried three literature databases, namely PubMed, Web of Science (WoS), and Scopus, for original articles published up to February 16, 2022 (protocol summarized in Fig. 1). The search terms used for exhaustive searches against the three databases were: “exosome OR exosomes OR extracellular vesicle OR extracellular vesicles OR exosomal OR EVs OR EV OR small EVs OR small EV OR small extracellular vesicle OR small extracellular vesicles” AND “piRNA OR piRNAs OR piwi-interacting RNA OR piwi-interacting RNAs OR piwiRNA OR piwiRNAs OR piwi RNA OR piwi RNAs OR PIWI-interacting RNA OR PIWI-interacting RNAs OR piwi interacting RNA OR piwi interacting RNAs.” The articles identified by the searches were compiled and subjected to an initial screening based on their title and/or abstract. A total of 775 nonredundant original articles were identified.

Flow chart for the literature search. PRISMA workflow for the systemic selection process for articles included in this systematic review. PRISMA, Preferred Reporting Items for Reviews and Meta-Analyses.
Inclusion criteria were original research articles related to EV-piRNA derived from human clinical samples and published in English, whereas exclusion criteria were nonoriginal articles (i.e., review articles) and other document types, such as book chapters and meeting reports. In addition, articles that were not published in the English language, non-EV-piRNA relevant, reported on nonhuman species, reported on piRNAs isolated from in vitro cell culture supernatant-derived EVs, and reported solely on in silico analysis of EV-piRNA, were excluded.
Based on these inclusion and exclusion criteria, a total of 34 articles were eligible for this systematic review. A full list of the selected articles is shown in Table 1. The trend for research in this area is increasing trend (Fig. 2); no EV-piRNA publications related to human clinical samples were reported in 2015 and 2017.
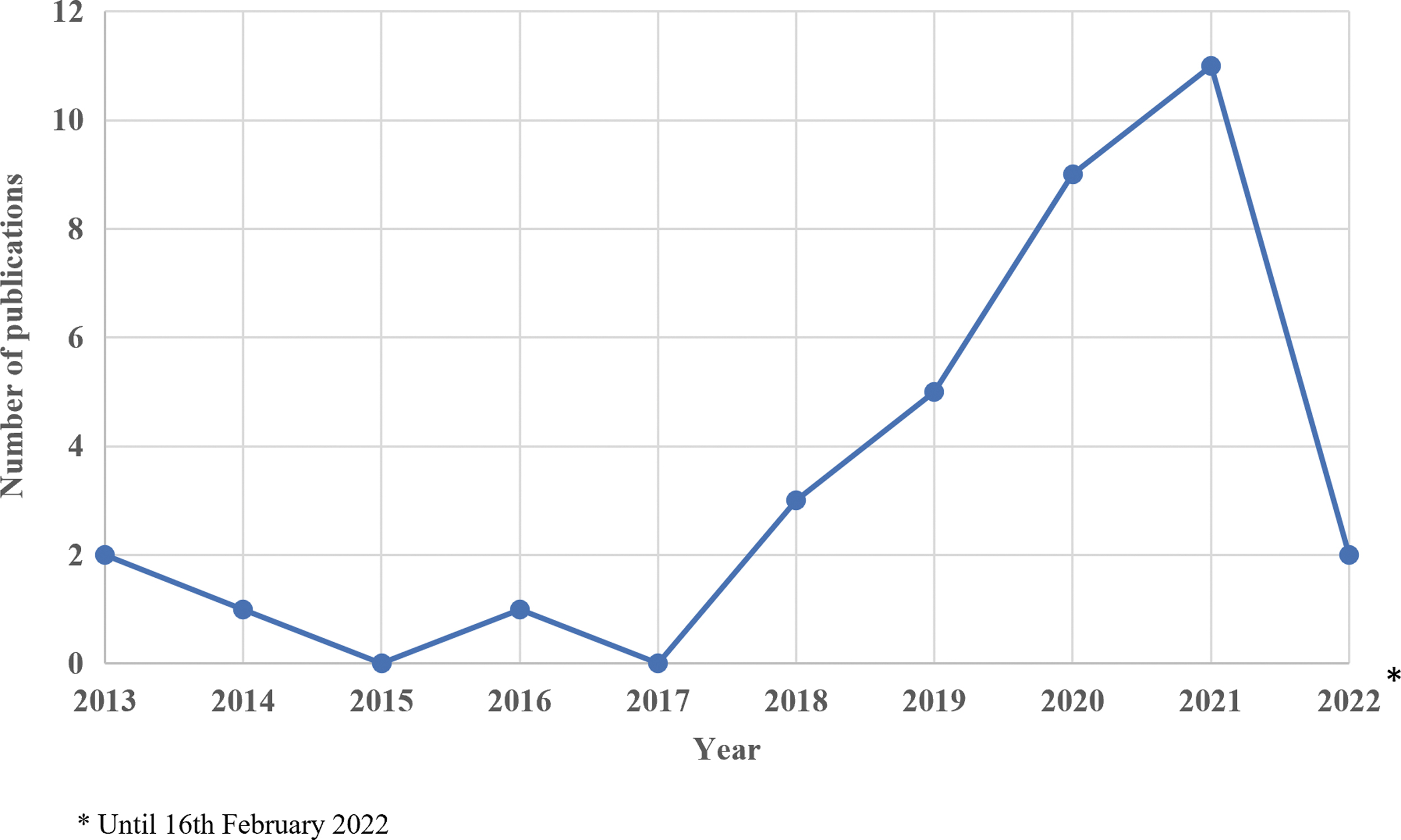
Trend of publication on human biofluid-derived EV-piRNA from the year 2013 to 2022. Review queried conducted up to February 16, 2022, for the year 2022. EV, extracellular vesicle; EV-piRNA, EV-derived piRNA; piRNA, Piwi-interacting RNA.
Full List of Original Articles Reviewed (n = 34) and the Pipeline/Software and Piwi-Interacting RNA Database Used for Piwi-Interacting RNA Identification
GRCh Genome Reference Consortium Human genome build; piRNABank (
piRNA, Piwi-interacting RNA.
In addition, piRNA sequence and the putative target genes of the reported EV-piRNAs, either highly expressed in the biofluids or significantly DE in a particular clinical condition, were retrieved from piRNABank (
Results
Clinical conditions and sources of EV-piRNA
Subjects diagnosed with defined clinical conditions were recruited together with their matched controls. piRNA identification and analysis were performed on the DE small RNA profiles that could potentially characterize disease pathological conditions (Supplementary Data S1). The frequency of each clinical condition category investigated is summarized in Figure 3a. From this review, cancer is the most studied human disease for EV-piRNA identification, which accounts for 41% (14/34) of the articles reviewed. Pulmonary/cardiovascular disease (n = 5), reproductive disorder (n = 4), and neurological disorder (n = 3) have been widely studied for their EV-piRNA. The aim of these studies was to identify potential piRNA candidates to serve as diagnostic biomarkers or therapeutic targets. No studies have yet reported on aging-related degenerative noncommunicable diseases such as osteoarthritis (OA) and other musculoskeletal disorders.
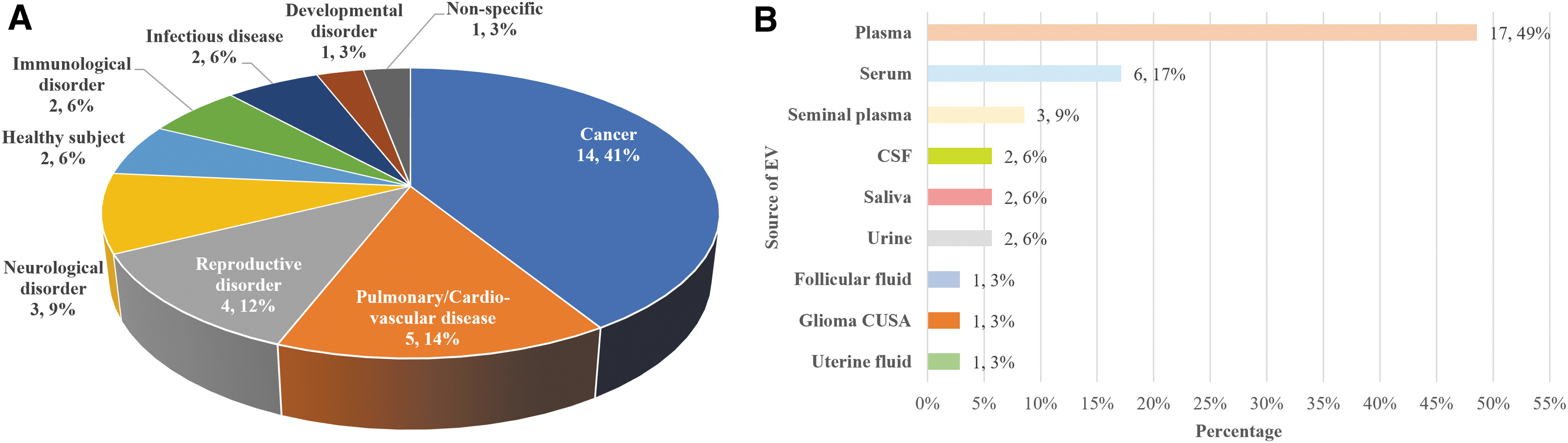
Clinical conditions and source of EVs where EV-piRNA was identified or analyzed.
EV-piRNAs have been identified from various human body fluids, including plasma, serum, seminal plasma, cerebrospinal fluid, saliva, urine, follicular fluid, glioma Cavitron Ultrasonic Surgical Aspirator (CUSA), and uterine fluid. Upon EV extraction, total RNA was further extracted for genomic analysis followed by bioinformatics analysis (Fig. 3b and Supplementary Data S1). Plasma is the most common source of EV-piRNA for piRNA discovery and identification, accounting for 49% of all sample types (17/35), followed by serum, reported in a total of six articles.
EV isolation method
EVs can be isolated using different methods as summarized in Figure 4 and Table 2 (Supplementary Data S2). The most commonly used approach in the articles reviewed for this study and performed by 40% were centrifugation-based methods (16/40). These include ultracentrifugation, differential ultracentrifugation, and density gradient ultracentrifugation. A further 35% of studies (14/40) performed polymer-based precipitation.
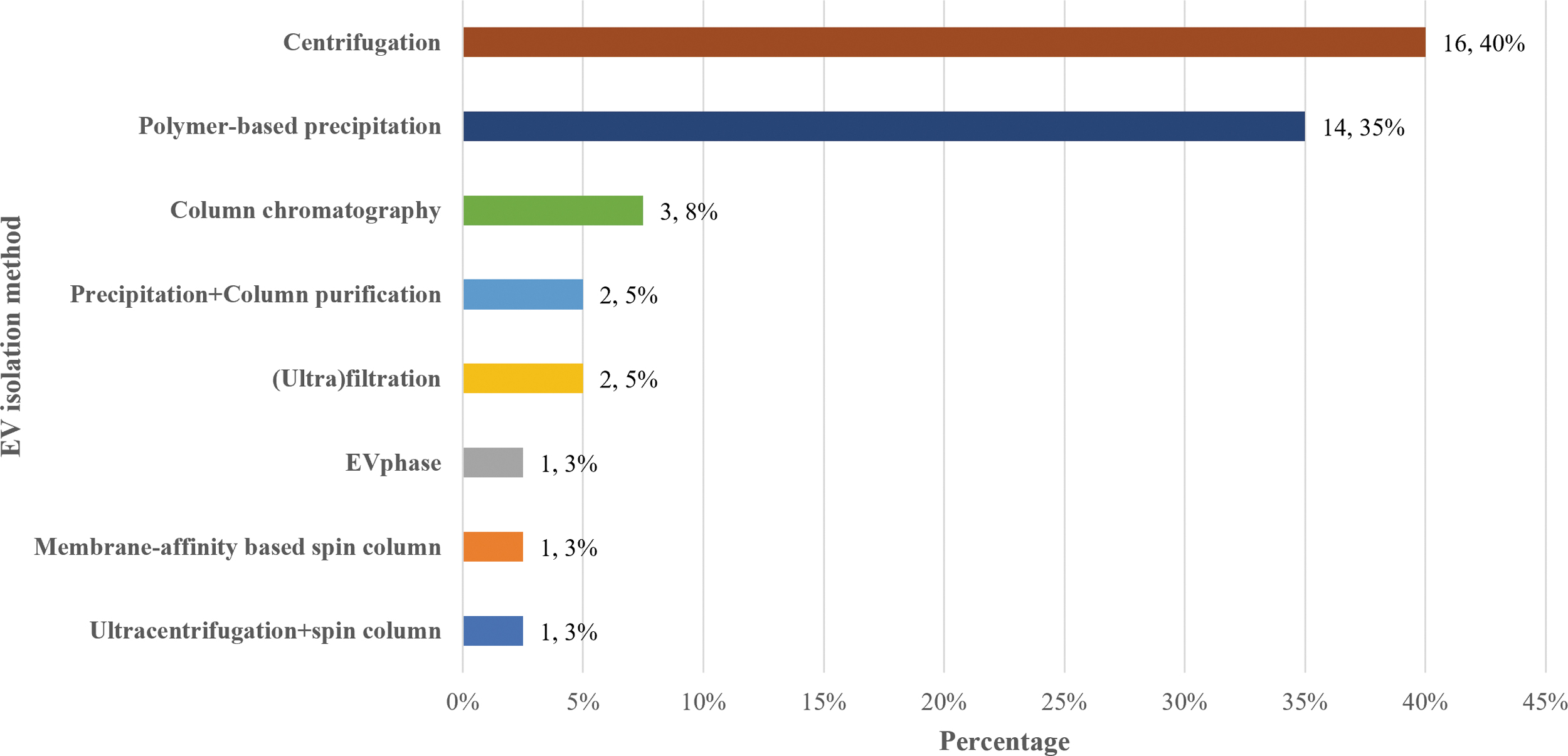
EV isolation methods categorized based on the principle of each methoda: ultracentrifugation, polymer-precipitation-based, column chromatography, ultrafiltration, EV phase, membrane affinity-based spin column, and combination of two methods. aA total of three articles used more than one method to isolate EV, and one article did not report on the isolation method used.
Extracellular Vesicle Isolation Methods Reported in the Reviewed Articles (n = 34)
One article did not report on the isolation method used, and a total of three articles used more than one method to isolate EV.
CSF, cerebrospinal fluid; EV, extracellular vesicle; PEG, polyethylene glycol.
Ultracentrifugation is a commonly used method for EV isolation, as it can separate the vesicles from other materials, such as cellular debris and macromolecular proteins while retaining the vesicular RNA components. However, ultracentrifugation is a time-consuming and heavily instrument-dependent technique. In addition, it cannot separate vesicle subtypes with overlapping sizes, which could result in low purity. In contrast, density gradient ultracentrifugation has a greater separation efficiency and thus results in higher purity as different EVs present different densities regardless of their size, reflecting their cellular origin. 12
Another widely used method for isolating EVs is the polymer-based precipitation method. In this method, the fluidic samples are first mixed with a polymeric reagent, which reacts and forms a polymeric web to capture the vesicles according to size. 13 A precipitating reagent is then applied to pellet the EVs at low centrifugal speed. As this method largely depends on the size, it can isolate vesicles with great uniformity in size; however, it can also include contaminants, such as lipoprotein that are within the same size range. An example of this method is the ExoQuick™ Ultra Kit which combines the precipitation-based method with a purification column that increases the yield and purity of EV isolated.
Column chromatography is an EV isolation principle applied in Norgen's Plasma/Serum Exosome and Free-Circulating RNA Isolation Mini Kit. In this technique, the sample is loaded onto a Mini Filter spin column with a silicon carbide resin separation matrix. This method can facilitate the subsequent isolation of vesicular total RNA, as no centrifugal force is applied that could potentially damage the EVs. Similarly, Qiagen's exoRNeasy Serum/Plasma Kit also allows the isolation of vesicular RNA in the subsequent steps, as it makes use of the membrane affinity-based spin column to capture EVs from sample fluid without subjecting to high centrifugation speed.
Ultrafiltration membranes are also used for EV isolation, in which the isolation is dependant on the pore size of the membrane. As the EVs are concentrated or loaded onto the membrane, this can lead to EV loss because of poor recovery from the membrane. In addition, the need to apply additional force to push the sample fluid through the filter membrane could potentially damage the EVs. 13 Another less commonly used technique is Exo2D™, an EV isolation kit based on the surface property of EVs. In this technique, biofluid-containing EVs are mixed with an aqueous two-phase buffer, in which the EVs can be isolated to one of the phases through the interaction between EVs' surface and each phase. The principle of this method is called “EV phase” and has been suggested to yield EVs with higher purity than ultracentrifugation and ExoQuick. 14
EV characterization
The ISEV has established a guideline outlining the minimal information for studies of EVs (MISEV2018). 3 The recommendation is that three aspects of the isolated EV should be characterized: (1) quantification, for example, measuring the particle number and EV diameter using nanoparticle tracking analysis (NTA) or flow cytometry, (2) protein composition, for example, western blotting (WB) or flow cytometry can be performed to determine the presence of EVs and assess their purity, and (3) single vesicle analysis, for example, visualization techniques, including transmission electron microscopy (TEM), to observe EV morphology.
Regarding the protein markers, the ISEV recommended that researchers use at least one marker from each of the three main categories to characterize EVs. The three main categories are (1) Transmembrane protein, which can demonstrate the lipid-bilayer structure specific to each EV subtype, such as tetraspanins (CD9, CD63, and CD81); (2) Cytosolic protein, to determine that the isolated vesicles enclose cellular contents, including TSG101, ALIX, Flotillins, and heat shock proteins; and (3) Proteins that are major components of non-EV structures (negative marker), to assess the purity, such as lipoproteins APOA1/2. Other protein markers coexpressed in the intracellular compartments (e.g., mitochondria and endoplasmic reticulum) can also serve as negative markers for EV, such as calnexin and GM130.
Our systematic review noted that before the publication of MISEV2018 guideline, only one article reported the characterization of EVs which fulfilled the criteria recommended in MISEV2018 guidelines 15 ; most of the studies reported did not perform EV characterization in all three aspects. In most studies, the markers used to assess protein composition fit only one or two categories. After publication of MISEV2018 guidelines, there was an increase of 12% in published articles that followed the MISEV2018 characterization criteria (Fig. 5a). Most of the studies performed NTA, TEM, and WB to characterize the isolated EVs. Other methods include flow cytometry and mass spectrometry (Fig. 5b). Of note, most of the studies failed MISEV2018 guideline compliance from their lack of negative markers used in EV characterization.
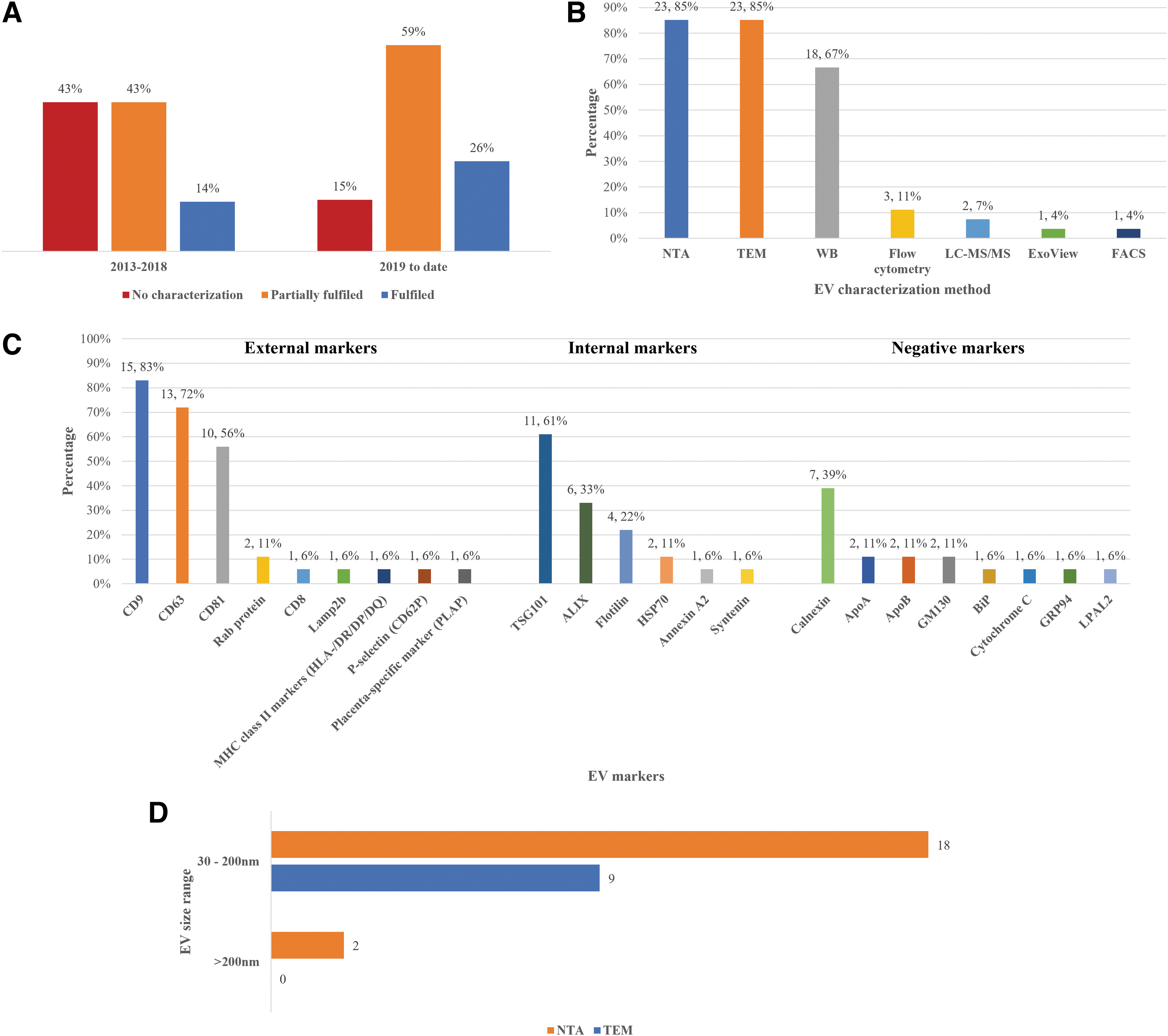
EVs were characterized based on their expression markers and/or size.
Apart from quantification with NTA and single EV imaging with TEM, most studies performed WB to characterize the isolated EV (Supplementary Data S3). The protein markers widely used in WB for EV characterization are (1) external markers: CD9, CD63, and CD81, (2) internal markers: TSG101, ALIX, and Flotillins, and (3) negative markers: Calnexin, GM130, and Apolipoproteins (Fig. 5c). Moreover, the NTA and TEM results suggested that the diameter of EVs mostly ranged from 30 to 200 nm, corresponding to the exosome subtype, with a lower number of EVs larger than 200 nm; indicating the isolation of various EV subtypes (Fig. 5d and Supplementary Data S3).
Next-generation sequencing platforms
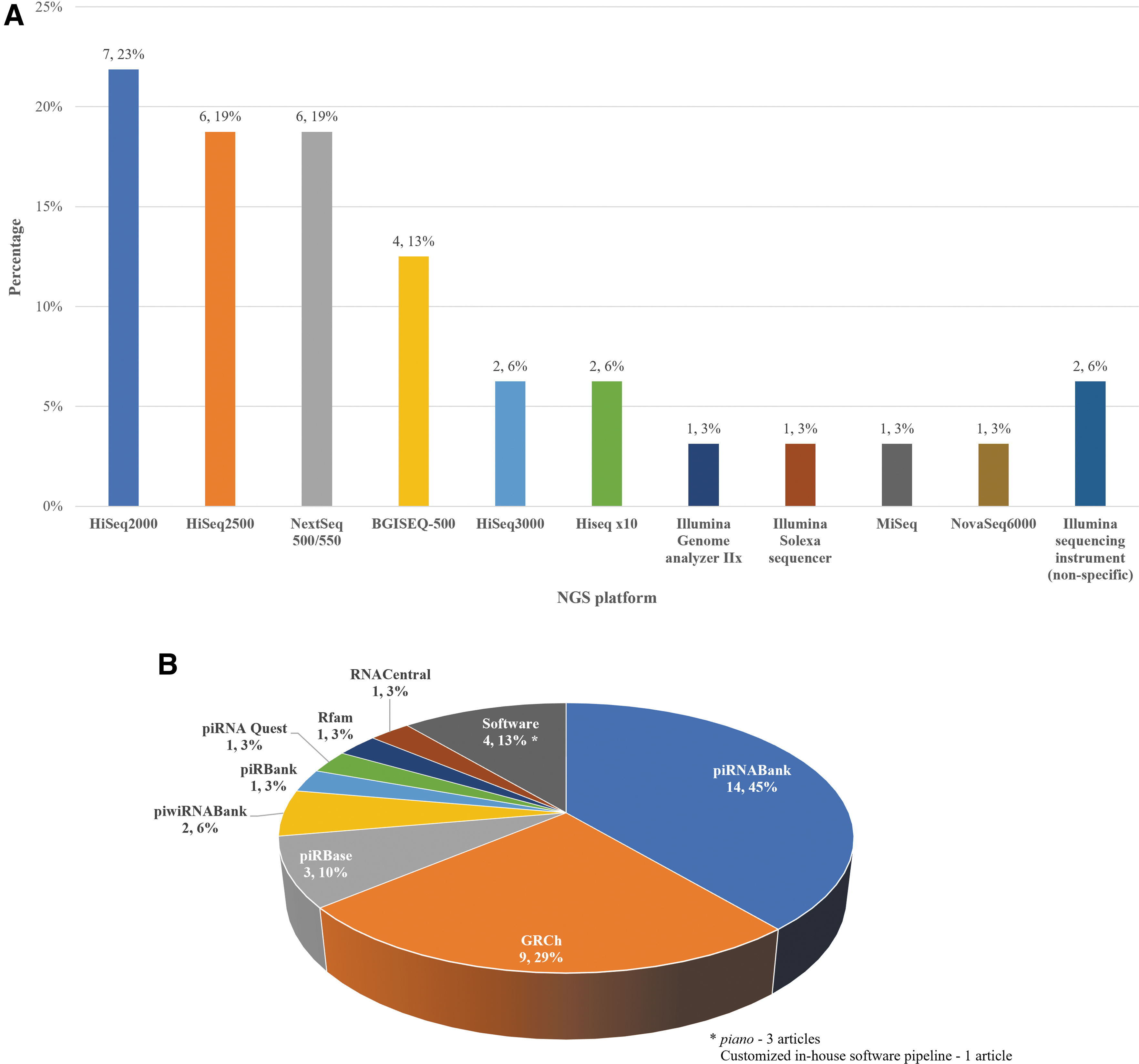
Most of the studies performed transcriptome sequencing using the Illumina next-generation sequencing (NGS) platform, with distinct systems, such as HiSeq2500 and NextSeq500, apart from four studies which used the BGISEQ-500 platform (Fig. 6a and Supplementary Data S4.1). The distinct features of commonly used Illumina sequencing platforms are summarized in Table 3. Two studies did not either use or document any sequencing platform; these studies used the uniCHA system and/or performed quantitative real-time polymerase chain reaction to assess the expression levels of piRNA of interest.
NGS and bioinformatics analysis on EV-piRNA.
Comparison Between Four Types of Illumina Next-Generation Sequencing Platforms
WGS, whole genome sequencing.
Illumina NGS platform uses a fluorescence-based paradigm for reading the bases in a nucleotide sequence, and all sequence reads generated during a single experiment have the same lengths. Moreover, it allows paired-end sequencing (read from both ends of a fragment). In contrast, the BGISEQ-500 sequencing platform is featured with combinatorial Probe-Anchor Synthesis (cPAS) and DNA Nanoballs (DNB) technology. The DNA sample is anchored on the DNB, and amplification is initiated once the fluorescent probe is incorporated onto the DNA sample by cPAS. BGISEQ-500 has been demonstrated to provide a comparative performance in gene quantification with the Illumina HiSeq4000 sequencing platform. 16
Bioinformatics analysis of piRNA expression profiles
Upon sequencing, the raw data have to be filtered and aligned to the reference genome and database to annotate for the corresponding small RNA type. For piRNA identification, the most widely reported database is piRNABank (Fig. 6b and Table 1). Other databases, including piRBase, human genome reference database (GRCh), and piRNA Quest, are also available (Supplementary Data S4.1).
A total of 14 articles have reported on the expression levels of piRNA among EV small RNA profile (Supplementary Data S4.1). The results showed that EV-piRNA expression level ranges from 0.09% to 43.84% of the EV small RNAs, with the lowest expression levels reported in urine-derived EVs and the highest percentage in plasma-derived EVs.
Among all sample types, seminal plasma has been reported with the highest quantity of EV-piRNAs, with 2344 piRNAs detected; the next most abundant source were serum samples which reported 533 piRNAs. A total of 195 piRNAs commonly expressed among at least two sample types (Supplementary Data S4.2). Serum-derived EV and glioma CUSA-derived EV reported the highest number of commonly expressed EV-piRNA, with 43 commonly expressed EV-piRNAs, followed by plasma-derived EV and seminal plasma-derived EV, with 23 commonly expressed EV-piRNAs.
Furthermore, DE EV-piRNA profiles in subjects diagnosed with clinical conditions of interest compared to their matched control were revealed (Supplementary Data S4.1). This suggests that EV-piRNA can potentially serve as a diagnostic biomarker and might play a role in disease progression. However, not all studies identifying EV-piRNA conducted a downstream analysis to explore the potential role of piRNA in each disease pathology (Supplementary Data S4.1). Among these, five studies investigated the putative target genes of piRNA candidates by utilizing software or databases, such as miRanda database and RNAhybrid algorithm. In addition, several studies performed Gene Ontology (GO) enrichment, Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis, or Ingenuity Pathway Analysis (IPA) to explore the piRNA involving pathways (based on the predicted piRNA target genes) (Supplementary Data S4.1).
Only one study reported a direct role of piRNA candidates in promoting disease pathology. Li et al explored the potential of multiple myeloma (MM) cell-derived EVs in delivering piRNA-823 to endothelial cells. 17 The treatment of MM-derived EVs carrying piRNA-823 has increased cell proliferation and attenuated apoptosis of endothelial cells, suggesting a role of piRNA-823 in promoting tumorigenesis.
In addition, to providing a reference for future studies on the role of EV-piRNA in human diseases, putative target genes of the reported EV-piRNAs, either highly expressed EV-piRNAs or significant DE EV-piRNAs in each article, were retrieved from piRNA databases and summarized (Table 4).
Putative Target Genes(s) of the Reported Extracellular Vesicle-Derived Piwi-Interacting RNAs in each Article (n = 31).
Not found in piRNA database.
CUSA, Cavitron Ultrasonic Surgical Aspirator; DE, differentially expressed; lncRNA, long noncoding RNA; tRNA, transfer RNA; SINE, short interspersed nuclear element.
In Table 4, five piRNA candidates were reported to be highly expressed in healthy subjects, of which four piRNAs were from saliva-derived EV and one piRNA from plasma-derived EV. These piRNAs were not detected in other EV samples derived from any particular disease condition. Furthermore, putative target genes of these piRNA were analyzed, in which PRPH2 and HMCN1 were reported to be involved in vision, METTL21A and DHX15 were involved in mRNA transcription and splicing, and CREB1 as a transcription factor involved in various signaling pathways, such as inflammation. Hence, these piRNAs can potentially serve as a negative biomarker or a therapeutic target in human disease.
Of note, some of the novel piRNAs reported through piano are undetectable in current piRNA databases, including piRNABank and piRBase.
Discussion
This systematic review examined 34 articles that reported on human body fluid-derived EV-piRNA. A detailed analysis of the EV-piRNA was conducted, in which the isolation and characterization methods of the EV, sequencing platform, and bioinformatics analysis findings of the EV-piRNA in each article were reported here. This review can serve as a reference for future studies on the analysis workflow of EV-piRNA.
Due to the ability to transport cellular contents and regulate gene transcripts in the targeted cells, EVs have emerged as a source for biomarker discovery, as well as an important element in cell-free therapy. This review revealed that piRNA can be detected in EVs, with the expression levels ranging from 0.09% to 43.84% of the EV small RNAs. Moreover, most studies reported high percentages of unannotated sequencing reads. This suggests that there might be still unidentified piRNAs contributing to the low number of piRNA detected. To address this problem, three studies reported on the use of piano software, in revealing novel piRNA. However, some of the novel piRNAs reported through piano are undetectable in current piRNA databases, including piRNABank and piRBase. Hence, the identity of these piRNAs remains putative.
In addition, by analyzing EV-piRNAs reported in each study, a total of 195 EV-piRNAs were found to be commonly expressed among different sample types. However, it is important to note that different piRNA databases were used for small RNA annotation among each article. As the nomenclature of the piRNA is not consistent among databases, this could lead to redundancy in piRNA reported, and thus, it cannot be concluded yet that the piRNA candidates identified from each EV source are tissue specific.
Furthermore, it is noted that only one study has demonstrated a direct correlation between EV-piRNAs and cellular behaviours, such as cell proliferation and cell death, in MM. As such, the exact role of EV-piRNA in other disease conditions is still unclear. In addition, the EV-piRNA expression profile also remains unexplored in various sample types of different disease conditions, that is, osteoarthritic synovial fluid.
OA is a noncommunicable degenerative disease that occurs in the knee joint and is characterized by low-grade systemic inflammation and cartilage damage. Mesenchymal stem cells (MSCs) have been proposed as a feasible cell source for cartilage repair, as they can differentiate into chondrocytes 18 and promote the synthesis of cartilage matrix components, such as type II collagen and proteoglycans.
The role of noncoding RNAs, such as miRNA and lncRNA, has been investigated in OA, with a large number of candidates being implicated in the pathological changes of OA, including chondrocyte apoptosis and extracellular matrix degradation. 19 In addition, a few piRNA candidates have also been reported to be involved in inflammation in OA, 20 as well as chondrogenesis of MSCs. 21 However, there is a scarcity of downstream analysis of the functional role of these EV-piRNAs. Thus, together with the results of the systematic review, this confirms the need for future studies to unravel the expression profile and mechanism of EV-piRNA in different sample types, to fully explore the potential of EV-piRNA as a novel therapeutic intervention in human diseases.
In vivo and in vitro studies are useful approaches to demonstrate the functional role of EV-piRNA. piRNA functions together with PIWI proteins to mediate gene silencing, through complementary base pairing with its mRNA target, or through epigenetic regulation. Hence, to study the functional role of EV-piRNA in animal models, homologous gene and protein function, as well as the mechanism of function of piRNA/PIWI complex among species, need to be understood. Previous studies have reported on the highly conserved structure of PIWI proteins among zebrafish, mice, and humans, 22 as well as similar piRNA biogenesis pathway and mechanism of function of piRNA/PIWI complex in gene regulation among Drosophila, murine, and human. 23 In addition, comparative genomic studies have revealed gene homology between humans and mice. 24 Hence, these findings suggest the potential application of animal models in understanding the potential of EV-piRNA as an RNA-based drug before translation into clinical studies.
piRNA is still an emerging field in human disease; however, it is apparent that the current bioinformatics analysis database and software for piRNA and the experimental analysis of piRNA functional role are limited and often lacking entirely. Of particular concern is that the piRNA nomenclature in different piRNA databases is inconsistent, suggesting an urgent need for a harmonized piRNA nomenclature to be established. Alternatively, researchers should annotate the sequencing data on more than one piRNA database, to acquire a more complete piRNA identification and expression profile analysis, as well as to screen for redundancy across the different databases. It is important to improve the bioinformatics analysis platform, not only for a harmonized annotated database but also for a standardized guidelines for putative piRNA target gene prediction software/pipeline, to accelerate the study of EV-piRNA functional role in human disease.
Despite this systematic review reporting the current limitations in EV-piRNA studies regarding the bioinformatics analysis platform of piRNA, most of the studies partially or completely fulfilled the ISEV guideline in EV characterization. In addition, the sequencing platforms used are mostly Illumina sequencers, which provide consistency and quality in the sequencing reads. Thus, this review regards it as confirmed that EV-piRNA can be derived from various biofluid samples with confidence.
Conclusion
Our systematic review has demonstrated that piRNA is stably expressed in EV derived from various human body fluids, and DE piRNAs are also reported between subjects with specific clinical conditions and healthy controls. It is crucial to investigate the role of EV-piRNAs in human disease pathology, which could lead to the discovery of a diagnostic biomarker or the development of a novel therapeutic intervention.
Footnotes
Authors' Contributions
T.-X.G. has conducted article search, data collection, and article writing. S.-L.T., M.M.R., S.-H.T., and T.K. supervised and contributed to the article writing.
Disclosure Statement
Authors declare no conflict of interest.
Funding Information
This study is part of the Fundamental Research Grant Scheme (FRGS) project funded by the Ministry of Higher Education (reference number: FP039-2017A and FRGS/1/2017/STG05/UM/02/9).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.