
Abstract
The synthesis and assembly of mature, organized elastic fibers remains a limitation to the clinical use of many engineered tissue replacements. There is a critical need for a more in-depth understanding of elastogenesis regulation for the advancement of methods to induce and guide production of elastic matrix structures in engineered tissues that meet the structural and functional requirements of native tissue. The dramatic increase in elastic fibers through normal pregnancy has led us to explore the potential role of mechanical stretch in combination with pregnancy levels of the steroid hormones 17β-estradiol and progesterone on elastic fiber production by human uterine myometrial smooth muscle cells in a three-dimensional (3D) culture model. Opposed to a single strain regimen, we sought to better understand how the amplitude and frequency parameters of cyclic strain influence elastic fiber production in these myometrial tissue constructs (MTC). Mechanical stretch was applied to MTC at a range of strain amplitudes (5%, 10%, and 15% at 0.5 Hz frequency) and frequencies (0.1 Hz, 0.5 Hz, 1 Hz, and constant 0 Hz at 10% amplitude), with and without pregnancy-level hormones, for 6 days. MTC were assessed for cell proliferation, matrix elastin protein content, and expression of the main elastic fiber genes, tropoelastin (ELN) and fibrillin-1 (FBN1). Significant increases in elastin protein and ELN and FBN1 mRNA were produced from samples subjected to a 0.5 Hz, 10% strain regimen, as well as samples stretched at higher amplitude (15%, 0.5 Hz) and higher frequency (1 Hz, 10%); however, no significant effects because of third-trimester mimetic hormone treatment were determined. These results establish that a minimum level of strain is required to stimulate the synthesis of elastic fiber components in our culture model and show this response can be similarly enhanced by increasing either the amplitude or frequency parameter of applied strain. Further, our results demonstrate strain alone is sufficient to stimulate elastic fiber production and suggest hormones may not be a significant factor in regulating elastin synthesis. This 3D culture model will provide a useful tool to further investigate mechanisms underlying pregnancy-induced de novo elastic fiber synthesis and assembly by uterine smooth muscle cells.
Impact Statement
The results of this study add to the growing body of literature exploring the important effects of mechanical forces on elastin production by adult cells in a three-dimensional culture environment, critical for the advancement of methods for the regeneration or replacement of engineered elastic tissues with tailored biomechanical properties.
Introduction
Elastic fibers are macromolecular structures of the extracellular matrix (ECM) that allow tissues such as blood vessels, lungs, and skin to stretch. They are made up of an amorphous elastin core surrounded by a microfibril scaffold. Although the overall mechanical properties of a tissue ECM result from a composite of structural proteins, its mechanical integrity and elasticity are predominantly attributed to elastin. 1 The primary function of elastic fibers is to enable the repetitive and reversible deformation of a tissue in response to stretch. In addition to their role in the mechanical support system, elastic fibers also play an essential role in maintaining normal tissue function by modulating signaling events that regulate cell phenotype, adhesion, proliferation, migration, and matrix synthesis. 2
As adult cells do not spontaneously regenerate elastic fibers in vivo or in vitro, finding the necessary conditions to stimulate and guide the formation of new elastic fibers is a central challenge to achieving functional tissue regeneration. Based on previous research summarized by Yeo et al., we know that elastin (ELN) gene expression is a limiting factor in tissue engineering and regeneration. 3 Although a variety of strategies have been investigated, the limited success of bioengineering approaches to generate elastin structures similar to native tissue highlights the need to better understand the dynamic machinery regulating elastogenesis.4,5
Although dogma states that elastic fiber synthesis effectively ceases after adolescence, the transient reactivation of elastogenesis in the uterus during pregnancy is a noted exception.6,7 Associated with a 250-fold increase in size and 11-fold increase in tissue weight, elastin in the human uterus increases five-fold by term pregnancy.6,8–10 The myometrium is the smooth muscle layer of the uterine wall and main component in the enlargement of the gravid uterus. As the only occurrence of physiological elastogenesis in adult life, the study of the gravid uterus presents a valuable and unique opportunity to elucidate mechanisms regulating functional de novo elastic fiber synthesis and organization by adult cells. Previous research has demonstrated mechanical stretch and endocrine signals synergistically drive uterine growth and matrix remodeling events required for successful pregnancy.11,12 However, how these signals may regulate elastic fiber synthesis specifically is largely unexplored.
Cyclic strain is well established to stimulate elastin synthesis in engineered tissues; however, the amplitude, frequency, and duration of strain are all important factors which influence the mechanotransductive response by cells and thus cell phenotype and matrix synthesis.13–16 The effects of cyclic strain amplitude on smooth muscle cell (SMC) phenotypic state and matrix remodeling have been reported in several studies, while others have reported on the effects of strain frequency on SMC responses.17–24 To date, the impact of these two strain parameters has not been investigated concurrently. To obtain optimal strategies to control SMC behavior, investigation of the role of cyclic strain amplitude and frequency would provide valuable and needed insight.
The uterus experiences a wide range of cyclic strains during different stages of the menstrual cycle and pregnancy with amplitudes of 5 − 20% and frequencies of 0.1–0.5 Hz.25–27 The uterine myometrium produces continuous motion, with low amplitude and higher frequency that follows a sine-wave pattern during much of the menstrual cycle, to higher amplitude, lower frequency contractions during menses, as discussed in Bulletti et al. 28 The nonpregnant uterus experiences amplitudes of 5 − 12% and frequencies not exceeding 0.1–0.5 Hz during the menstrual cycle. 28 The exact stresses and strains in the pregnant uterus through gestation and labor are difficult to completely elucidate as there is a combination of normal uterine motility, intrauterine pressure from the growing fetus, and fetal movements. 27 In the pregnant uterus of the rhesus macaque, the contractions change over time and in the third trimester average approximately 6 mmHg with a frequency of 6/h. 29
Despite constant motion, elastin synthesis in the uterus only occurs during pregnancy, and the excess elastin is degraded postpartum. That suggests that there are factors other than mechanical forces that influence elastin synthesis. Moreover, it is known that steroid hormones can intersect with mechanosignaling pathways to regulate gene expression that dictates ECM composition in a variety of adult tissues.30–33 17-β-estradiol (E2) and progesterone (P4) are well-known regulators of uterine functions during pregnancy with demonstrated roles in myometrium tissue growth and remodeling. 34 Previous research suggests that the synthesis and assembly of elastic fibers in the gravid uterus is mediated by concerted hormonal actions, supporting a potential role for steroid hormones in regulating elastic fiber synthesis in engineered tissue. 35 However, hormone-mediated effects on ECM synthesis in three-dimensional (3D) culture are not well understood.
The long-term goal of our research is to elucidate how elastogenesis is regulated in the gravid uterus. The goal of this study was to investigate the effects of mechanical strain and pregnancy-related hormones on elastic fiber production by human myometrial smooth muscle cells (hMSMC) in a 3D culture model. Opposed to a single strain regimen, we applied a series of cyclic strain amplitudes and frequencies to systematically compare the elastogenic response to a wide range of strain profiles. This was done first to determine a strain condition that stimulates elastin synthesis in our 3D model, and second to assess the effects of the different strain parameters on cell responses. To determine interactions between cyclic strain and hormones, we applied pregnancy levels of E2 and P4 alone and in combination with the various strain patterns.
Methods
Ethics statement
Primary hMSMC were isolated from myometrial tissue samples obtained from consenting patients undergoing hysterectomy or abdominal myomectomy for the treatment of uterine fibroids between June 2021 and February 2022 at the University of Cincinnati Medical Center or West Chester Hospital under the University of Cincinnati IRB-approved protocol 2020-0678. Patient characteristics are summarized in Supplementary Table S1.
Cell isolation and culture
Primary hMSMC were isolated from myometrial tissue samples using a combination of enzymatic digestion and explant migration techniques as described previously.
36
Primary hMSMC were subcultured under standard culture conditions using culture medium consisting of Dulbecco’s Modified Eagle Medium (DMEM; Corning, Tewksbury, MA, USA) with 5% fetal bovine serum (FBS; Corning), 1% antibiotic-antimycotic (Gibco, Grand Island, NY, USA), 5 ng/mL basic fibroblast growth factor (Sigma-Aldrich, St. Louis, MO, USA), 5 μg/mL insulin (Sigma-Aldrich), 5 ng/mL epidermal growth factor (Peprotech, Rocky Hill, NJ, USA), 10 mM
Fabrication of myometrial tissue constructs
Myometrial tissue constructs (MTC) were created by embedding hMSMC within a collagen hydrogel as described previously, with modifications.37,38 4 mg/mL bovine collagen type 1 (MP Biomedical, Irvine, CA, USA), 2× DMEM, and hMSMC were combined to form MTC with a final density of 5 × 105 cells/mL and 2 mg/mL collagen. Collagen–cell mixture (1.5 mL) was added into each well of a Tissue Train® collagen-coated culture plate (Flexcell International Corp., Hillsborough, NC, USA). Following polymerization for 1 h at 37°C, the MTC were statically cultured in quiescent culture medium consisting of phenol red-free DMEM/F12 50:50 (Gibco) with 1% antibiotic-antimycotic, 2 mM
Hormone treatment and application of mechanical strain
Treatment culture medium had the same composition as the quiescent culture medium, but with the addition of 5% charcoal-stripped FBS (CS-FBS; Corning) and physiological concentrations of E2 and P4 (both from Sigma-Aldrich). The majority of elastogenesis in the gravid human uterus occurs during the last trimester of pregnancy, therefore MTC were treated with E2 and P4 concentrations based on the average circulating levels in the third trimester of pregnancy (3TP; 130 µg/mL P4, 20 µg/mL E2) and compared with treatment with nonpregnant levels (NP; 13 µg/mL P4, 0.15 µg/mL E2) as a negative control. 39
Upon the addition of treatment medium, equibiaxial tensile strain was applied to MTC for 6 days using the Flexcell® FX6000 Tension System (Flexcell Int. Corp.) at a range of amplitudes (5%, 10%, and 15% at 0.5 Hz frequency) and frequencies (0 Hz, 0.1 Hz, 0.5 Hz, and 1 Hz at 10% amplitude). Static control MTC were cultured on the same Flexcell® plates, but without applied strain.
Measurement of cell density
Cell number was inferred from measurements of cell metabolism assessed using the PrestoBlue HS metabolic assay (Invitrogen, Waltham, MA, USA) similar to previously described.40,41 Briefly, MTC were incubated in culture medium containing PrestoBlue for 2 h under standard culture conditions. Then, the fluorescence of the solution was measured at λex = 560 nm; λem = 590 nm using a BioTek Citation 5 microplate reader (Agilent, Santa Clara, CA, USA). An acellular collagen gel was used as a control.
Elastin protein quantification
Matrix elastin content was measured in MTC using the FastinTM Elastin assay kit (Biocolor, Antrim, UK) following manufacturer instructions. Following strain, MTC were collected and dehydrated overnight in a 50°C dry oven. The next day, dehydrated constructs were weighed before going through three cycles of digestion in 0.25 M oxalic acid for 1 h at 100°C. The supernatant from each extraction was pooled for analysis. Absorbance of the dye-bound elastin solution was measured at 513 nm using the BioTek Cytation 5 microplate reader, and the quantity of elastin in samples was determined against a standard curve. Elastin (µg) was normalized to MTC dry weight (mg).
Gene expression analysis using quantitative polymerase chain reaction
Quantitative real-time reverse transcriptase-polymerase chain reaction (qRT-PCR) was used to evaluate the expression levels of elastin protein precursor, tropoelastin, and fibrillin-1 (FBN1), genes which encode the two main proteins that compose the elastin core and microfibril template of elastic fibers, respectfully. Total RNA was isolated using RNeasy Lipid Tissue Mini Kit (Qiagen, Venlo, Netherlands) per manufacturer protocols. RNA concentration and purity was determined using the BioTek Cytation 5 microplate reader. RNA was reverse transcribed into cDNA using the high-capacity RNA-to-cDNA kit (Applied Biosystems, Waltham, MA, USA). Forward and reverse primers (Integrated DNA Technologies, Coralville, IA, USA) and cDNA templates were used with iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) according to manufacture protocol, and qRT-PCR was performed using the CFX Opus Real-Time PCR System (Bio-Rad). Primer sequences are provided in Table 1. The 18S ribosomal RNA gene was used as an internal control, and relative fold differences in target gene mRNA expression in stretched MTC were calculated according to the 2-△△Ct method. 42
Primer Sequences for Target Genes Measured by qRT-PCR
ELN, tropoelastin; FBN1, fibrillin-1; 18S, 18S ribosomal RNA; qRT-PCR, quantitative real-time reverse transcriptase-polymerase chain reaction.
Immunohistochemistry
Following fixation in 4% paraformaldehyde at 4°C overnight, MTC were embedded in paraffin wax and processed for histological assessment of matrix elastin. 6 µm cross-sections were cut using a rotary microtome, dried overnight, dewaxed, incubated with citrate buffer (Biolegend, San Diego, CA) for antigen retrieval, permeabilized with 0.1% Triton X-100, blocked in 10% normal goat serum, then incubated overnight with primary antibody to elastin (AB9519; Abcam, Waltham, MA). MTC sections were then incubated with fluorescent-tagged secondary antibody FITC-conjugated AffiniPure Goat Anti-Mouse; Jackson ImmunoResearch, West Grove, PA) and counter-stained with Hoechst dye (Invitrogen) for nuclei staining, and finally imaged with an Olympus IX3 microscope. Paraffin-embedded tissue sections of de-identified discard skin tissue from elective procedures performed at the University of Cincinnati Medical Center or Shriners Hospital for Children-Cincinnati were stained as above and used as positive and negative elastin antibody control samples (Supplementary Fig. S1). The University of Cincinnati Institutional Review Board determined that this does not constitute human subjects research and is exempt from requirements for informed consent according to 45CFR46.101(b).
Experiment
A summary of the current research experiment is described herein and presented graphically in Figure 1. First, MTC were fabricated and quiesced in serum-free medium for 24 h. The next day (considered day one of the experiment), the cellularity of MTC was measured, then hormone treatments were applied, and samples were subjected to mechanical strain or maintained in static culture for six days. To investigate the elastogenic effects of hormones in addition to strain, both stretched and static samples were each treated with NP and 3TP E2+P4, for a total of four treatment groups. In separate experiments, six different patterns of strain were applied to MTC using the Flexcell® Tension System. For the amplitude series, a 0.5 Hz cyclic strain was applied at 5%, 10%, and 15% magnitude. For the frequency series, 10% strain was applied at a constant 0 Hz and cyclic 0.1 Hz, 0.5 Hz, and 1 Hz frequency. At the end of 6 days, cellularity was again measured, then MTC sample replicates were either snap frozen in liquid nitrogen and stored at −80°C until analysis of ELN and FBN1 gene expression or dehydrated overnight for subsequent analysis of elastin protein content.

Graphical summary of experiment and application of strain with the Flexcell® system.
Cell proliferation was assessed by determining the change in cell number on experiment day six from day one relative to static controls. For analysis of strain and hormone effects, NP and 3TP stretched samples and 3TP static samples were normalized to NP static controls. For analysis of strain effects alone, NP and 3TP stretched samples were grouped together following normalization to the respective static hormone controls.
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 10.0.3 (GraphPad Software, Boston, MA, USA). All results were analyzed from n = 3–6 biological replicates per condition; analytical measurements were made in duplicate with a minimum of two technical replicates per unique sample. Differences among strain and hormone treatment groups were analyzed by two-way analysis of variance (ANOVA) with post hoc Fisher’s LSD multiple comparisons test. For analyses of strain effects alone, differences among strain conditions were determined by one-way ANOVA with post hoc Tukey’s multiple comparisons test. All data are expressed as mean ± standard deviation. Except for absolute elastin protein levels, all data are presented as fold differences relative to the static control group. Significance was established for differences with a p value < 0.05.
Results
Patterns of strain amplitude and frequency, but not hormones, affect elastin protein content
No significant differences in matrix elastin protein content in MTC were found because of different hormone levels (Fig. 2A,B); however, when investigating strain level alone, significant effects on matrix elastin content because of both amplitude and frequency were found. For the amplitude experiments, elastin in MTC under 10% strain was significantly greater than in static controls (1.35 ± 0.1-fold; p = 0.0210) (Fig. 2C). The 15% amplitude condition led to a further significant increase in elastin protein from static (1.43 ± 0.1-fold; p = 0.0106) and was also statistically greater than MTC at 5% (p = 0.0360). For the frequency experiments, the 0.5 Hz (1.35 ± 0.1-fold; p = 0.0174) and 1 Hz (1.55 ± 0.16-fold; p = 0.0012) conditions resulted in significant increases; however, there was no difference in matrix elastin at 0.1 Hz or in the 0 Hz constant stretch samples compared with static controls (Fig. 2D).
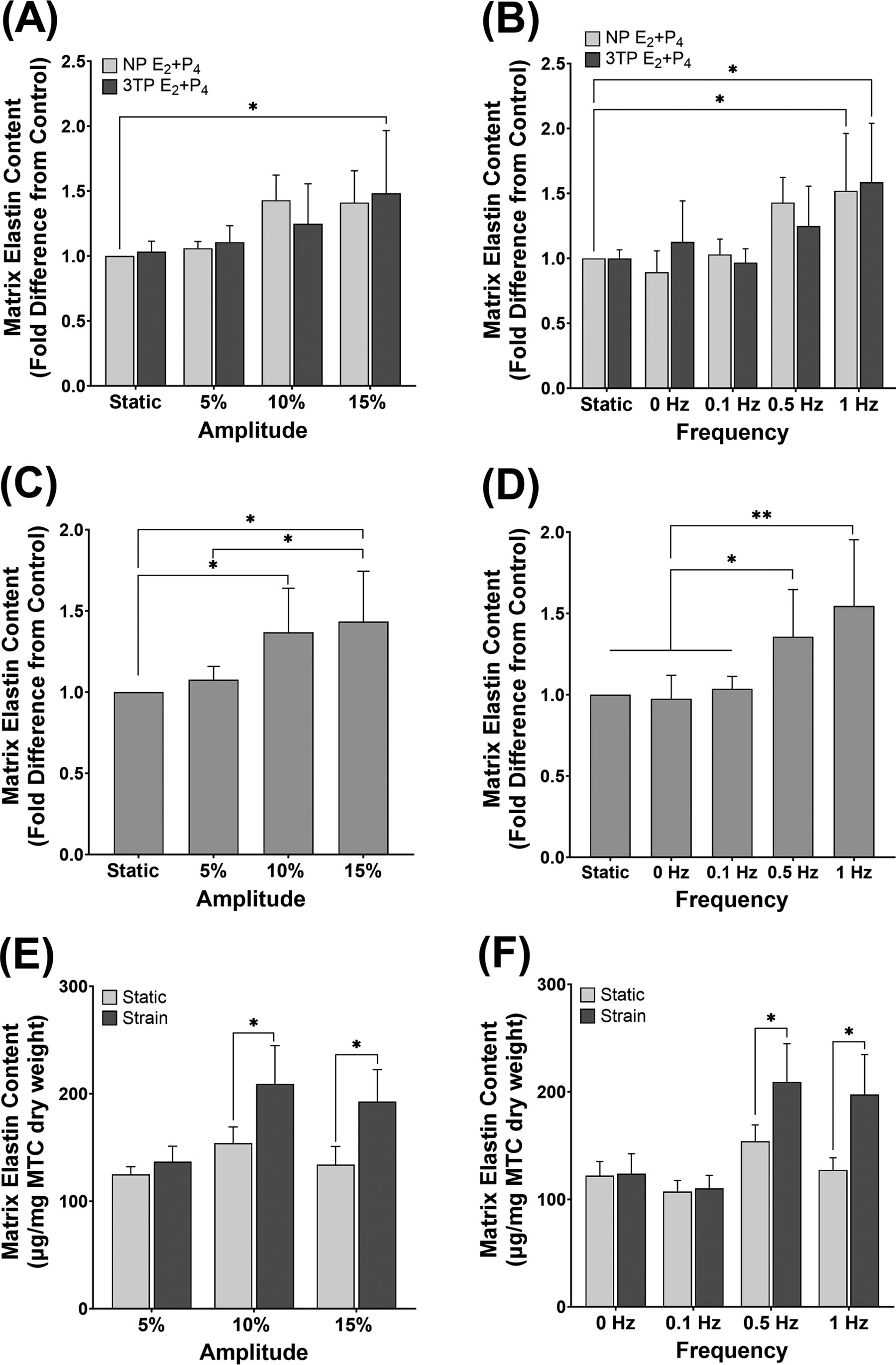
Effect of cyclic strain and pregnancy-level hormones on elastin protein content. MTC were treated with 3TP or NP concentrations of E2 and P4 and subjected to a range of strain
The absolute quantity of elastin protein (µg) normalized to mg dry weight of constructs is shown in Figure 2E,F. MTC strained at the 10%/0.5 Hz (209.17 ± 35.6; p = 0.042), 1 Hz (197.5 ± 37.23; p = 0.0177), and 15% (192.67 ± 29.86; p = 0.0186) conditions all show significantly higher quantity of elastin compared with the respective static controls. Elastin levels in MTC strained at 5% (136.83 ± 14.36), 0 Hz (123.83 ± 18.59), and 0.1 Hz (110.33 ± 11.98) were no different from static.
Figure 3 provides a visualization of matrix elastin in MTC under 15% strain and static control. This is used as a representative image to show the deposition of elastin in MTC under high strain where significant elastin protein levels were measured. The arrangement of elastin in the strained MTC (Fig. 3B) suggeststhe early formation of elastic fibers. Elastin can also be seen in the control MTC sample (Fig. 3A); however, the deposition is less and more fragmented compared with strain. The MTC under 15% strain is notably thinner than the static control because of cell-mediated compaction of the matrix in response to applied strain.
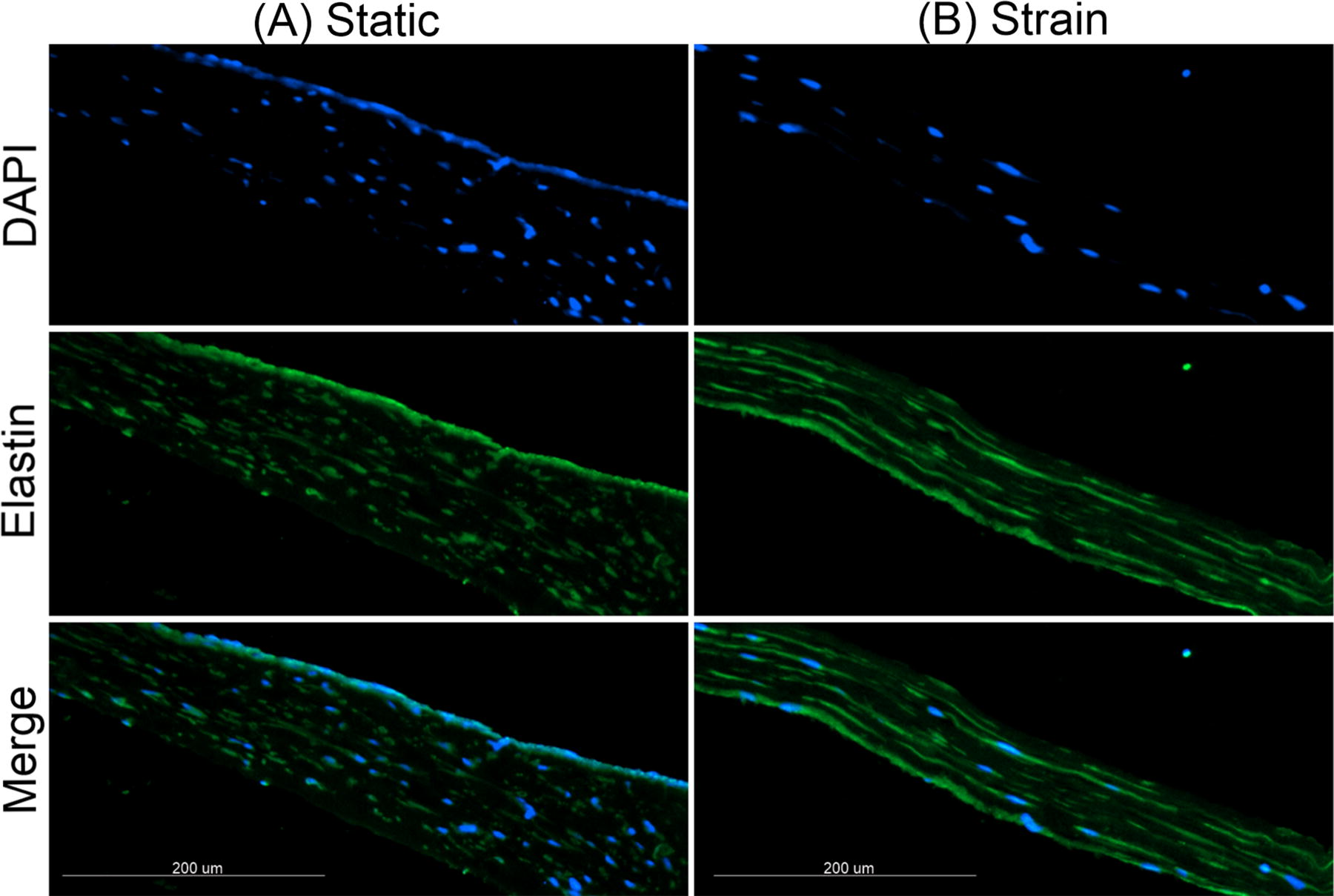
Effect of cyclic strain on synthesis and deposition of matrix elastin. Representative images of 6 µm thick cross sections (scale bar = 200 µm, 10× magnification) of MTC immunolabeled for elastin (green) and nuclei (blue) following 6 days of
Expression of core elastic fiber genes is affected by patterns of strain amplitude and frequency
qRT-PCR was performed to evaluate the effects of cyclic strains on the expression of core elastic fiber genes, ELN and FBN1. A significant increase in ELN mRNA relative to control was observed for MTC stretched at amplitudes of 10% (2.59 ± 0.19-fold; p ≤ 0.0001) and 15% (3.14 ± 0.21-fold p ≤ 0.0001) (Fig. 4A). ELN expression was significantly greater in samples at 0.5 Hz (p = 0.0001) and increasing the frequency to 1 Hz further increased expression to 2.81 ± 0.21-fold (p = 0.0002) compared with static control (Fig. 4B).

Effect of cyclic strain on elastic fiber gene expression. Normalized mRNA expression relative to static control for
Expression levels for FBN1 were in general lower than for ELN across all conditions; however, treatment-specific differences in FBN1 mRNA expression show similar trends to those for ELN mRNA. In the amplitude series, a 1.31 ± 0.04-fold increase at 10% strain (p = 0.0021) and 1.4 ± 0.05-fold increase at 15% strain (p = 0.0003) strain relative to static were observed (Fig. 4C). For frequency conditions, FBN1 expression was significantly increased in MTC stretched at 0.5 Hz (p = 0.0099) and 1 Hz (1.28 ± 0.04-fold; p = 0.0437) from control (Fig. 4D).
Cell proliferation is not affected by patterns of strain amplitude and frequency
There were no significant differences in the change in cell number after 6 days of strain relative to static controls for either the amplitude or frequency conditions, though proliferation in MTC at 15% amplitude was significantly less compared with 5%, (p = 0.0209) (Fig. 5). Similar to the results for elastin protein content, no differences in proliferation were found because of hormone treatment level (Supplementary Fig. S2).
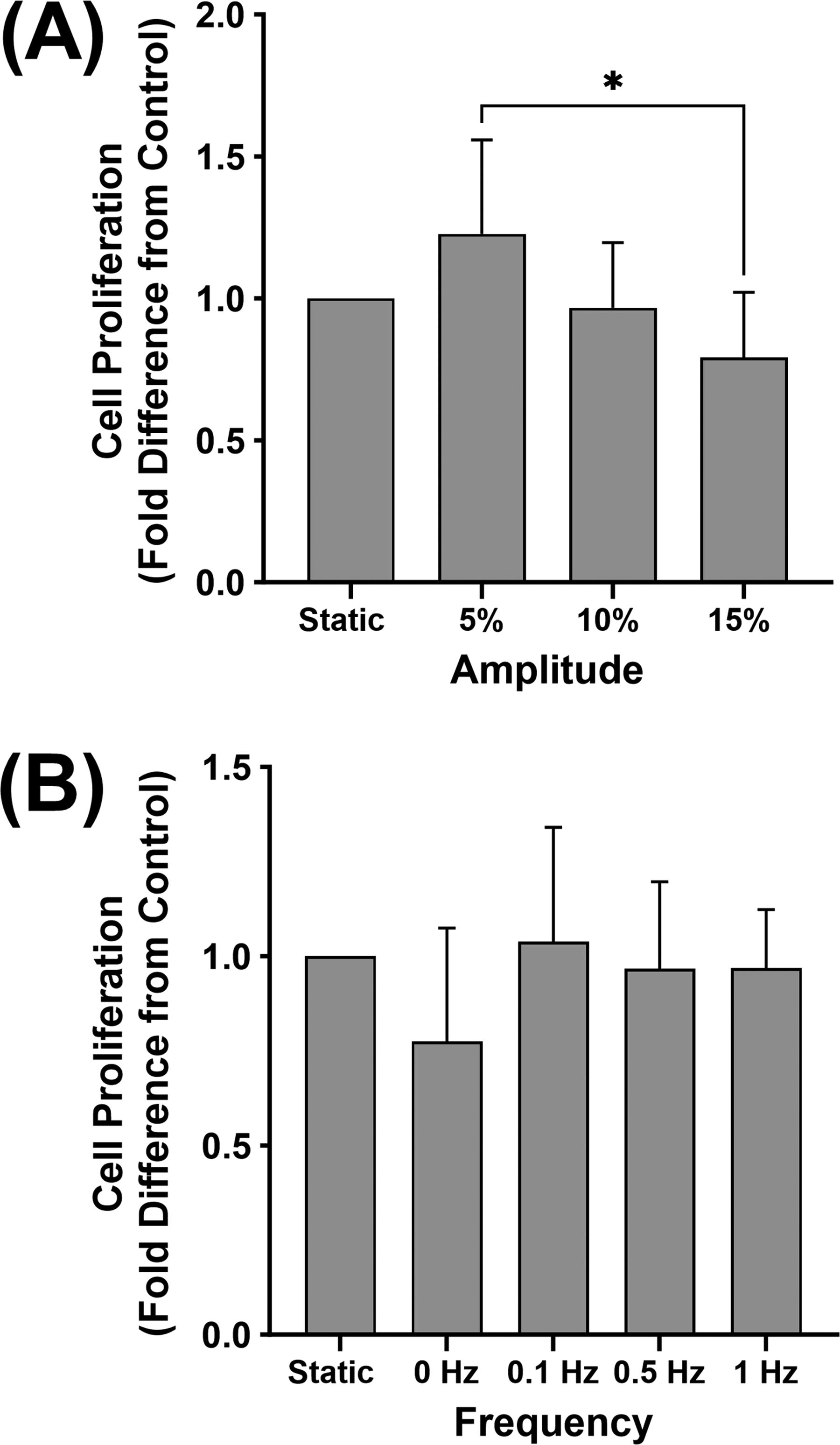
Effect of cyclic strain on cell proliferation. Change in MTC cell densities following 6 days exposure to a range of strain
Discussion
The results of our study show elastin protein and expression of elastic fiber genes increases in both an amplitude and frequency-dependent manner; significantly more matrix elastin and mRNA expression of ELN and FBN1 were produced in MTC stretched at 10%, 0.5 Hz compared with static, and even greater levels are produced by increasing either the frequency to 1 Hz or the amplitude to 15%. Although statistically significant, the elastin content was low because of the short 6-day culture time. We did not expect functional elastic fibers in this short time frame. Despite this and the low concentration of hMSMC, we saw significant increases in elastin at the mRNA and protein level and can see visible elastin in the MTC matrix.
The notion of a tailored mechanical profile has been shared by Isenberg et al. in a study that determined the effects of varying cyclic strain loading parameters (amplitude, pulse frequency, and pulse shape) on the development of collagen-based SMC constructs. 21 Their results show strain amplitude (2.5 − 10%), as well as the stretch and relaxation time, were important factors that influence the strength and stiffness properties of their engineered tissues. A significant increase in insoluble elastin was also observed in stretched constructs at 5%, 0.5 Hz compared with static; however, elastin was not measured under the various other strain profiles. In contrast to these findings, we did not see a significant difference in matrix elastin under 5% strain; however, these results are not directly comparable to ours, and the difference is likely associated with the longer culture period and use of highly compacted, cross-linked scaffolds in their study.
Regarding the influence of strain frequency, our results align with a similar study by Venkataraman et al. that determined the effect of varying cyclic frequency (0.5–3 Hz at 2.5%) on matrix production by aortic SMCs within a 3D collagen gel. 24 They report matrix elastin content and mRNA expression of ELN, FBN1, and the cross-linking enzyme lysyl oxidase were significantly increased in a frequency-dependent manner from 0.5 to 1.5 Hz, but all dropped to static levels at 3 Hz. The dramatic decline in expression levels at the 3 Hz frequency was suggested to be because of cellular damage resulting from the high strain. For our study, the largest differences were observed between the 10%, 0.5 Hz regimen, and 5% and 0.1 Hz profiles, with little difference between the effects at these lower strains and static culture. This indicates that a minimum level of mechanical stimuli is required, and it is not until some threshold value of strain is reached that the cells begin to respond differently, and we are able to observe a measurable response in elastin expression.
The changes in cell number after 6 days were not significantly different between strained and static MTC in accordance with other studies that applied 2.5–10% strains at frequencies of 0.5–3 Hz to SMC-seeded collagen gels.20–22,24 Cyclic strain can either stimulate or inhibit proliferation depending on the level of stimuli.43,44 As we applied a wide range of low to high strains, it was unexpected that no differences in proliferation were found between static and strained MTC for any strain profile. However, in addition to the level of applied strain, the effects of mechanical stretch on cell proliferation can vary depending on the specific cell type, culture conditions, and experimental setup.45–47 Although the reason for the results of strain on proliferation is currently unknown, these findings align with previous research conducted in our lab where various patterns of strain applied to hMSMC in monolayer had no effect on proliferation after 3 days. 36
Although our results show differences in response to the level of applied strain, there was no effect because of the level of E2 + P4 treatment on elastin protein content or cell proliferation alone or in combination with strain. Though we hypothesized that the addition of pregnancy-level hormones would enhance the elastogenic responses to strain, these results concur with the theory that mechanical stretch plays a more dominant role in driving uterine growth and ECM production. 48 Our results demonstrate strain alone is sufficient to stimulate elastic fiber production in MTC and suggest hormones may not be a significant factor in regulating elastin synthesis. In support of this notion, previous in vivo research shows strain is required for induction of elastin and other structural matrix proteins in rat uterus, and elastin expression levels are unaffected following treatment with a P4 receptor antagonist at late gestation. 12 Another study shows treatment with E2 increases elastin in the ovariectomized mouse uterus; however, treatment with P4 or the combination of E2 and P4 has no effect or even decreases ELN expression. 49 Their study also shows FBN1 expression was not significantly altered in response to either E2, P4, or both.
CS-FBS was required to control the concentrations of E2 and P4 in the treatment media; however, this also presents a limitation in the current study. CS-FBS has been shown to affect insulin-like growth factor (IGF) signaling pathways that promote cell proliferation and protein translation by removing several major proteins involved in this pathway. 50 The mitogenic effect of growth factors and the influence of serum concentration on stretched SMCs have been demonstrated by others.51–53 Further, MSMC hyperplasia during early pregnancy is suggested to be induced by E2-regulated growth factors such as IGF-1, thus the use of CS-FBS devoid of these endogenous cytokines and the importance of other factors that are elevated during this time may have contributed to the limited impact of strain on cell densities. Second, we did not explore the influence of strain on cell phenotypic state; however, this would have been beneficial to provide insight on the mechanisms underlying the observed effects on elastic fiber production. The significant levels of elastin synthesis observed in MTC under higher strain conditions could suggest a more synthetic phenotype. Likewise, lower strain conditions resulting in nonsignificant elastin production could suggest these hMSMC may be more contractile. While not a point of focus in the current study, understanding the impact of culture conditions in the control of SMC phenotype is highly important during engineered tissue development. 54 Future-related research is therefore suggested to determine effects on the expression of specific markers of differentiation.
Conclusions
This is the first study to substantiate the role of differential parameters of mechanical strain in combination with pregnancy-related hormones on elastic fiber synthesis by hMSMC in an engineered 3D culture environment. Our results demonstrate elastin protein and ELN and FBN1 gene expression increase in both an amplitude- and frequency-dependent manner. Treatment with pregnancy levels of E2 and P4 did not have a significant effect on these responses, suggesting these hormones do not directly influence elastin expression. Additional investigations are required to elucidate the intimate relationships existing among exposure to mechanical stretch and other biochemical stimuli in regulating MSMC elastic fiber deposition and remodeling in the gravid uterus.
Footnotes
Acknowledgments
The authors would like to extend special thanks to Rachel Warwar, MD, in the University of Cincinnati College of Medicine, Department of Obstetrics and Gynecology, for acquiring the human myometrium tissue samples from which the cells used in this study were derived. Also, the authors thank Dr. Greg Harris, Associate Professor in the University of Cincinnati Chemical Engineering Department, for allowing the use of his PCR machine.
Authors’ Contributions
C.A.N.: Conceptualization, experimental design, methodology, formal analysis, investigation, writing-original draft, and visualization. A.M.Z.: Experimental design, investigation, writing—review and editing. S.C.S.: Conceptualization, writing—review and editing, and supervision.
Disclosure Statement
No competing financial interests exist.
Funding Information
No external funding.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.