
Abstract
Ex vivo 3D culture of human tissue explants addresses many limitations of traditional monolayer cell culture techniques, namely the lack of cellular heterogeneity and absence of 3D intercellular spatial relationships, but presents challenges with regard to repeatability owing to the difficulty of acquiring multiple tissue samples from the same donor. In this study, we used a cryopreserved bank of human lung microexplants, ∼1 mm3 fragments of peripheral lung from donors undergoing lung resection surgery, and a liquid-like solid 3D culture matrix to describe a method for the analysis of non-small-cell lung cancer adhesion to human lung tissue. H226 (squamous cell carcinoma), H441 (lung adenocarcinoma), and H460 (large cell carcinoma) cell lines were cocultured with lung microexplants. Confocal fluorescence microscopy was used to visualize the adherence of each cell line to lung microexplants. Adherent cancer cells were quantified following filtration of nonadherent cells, digestion of cultured microexplants, and flow cytometry. This method was used to evaluate the role of integrins in cancer cell adherence. A statistically significant decrease in the adherence of H460 cells to lung microexplants was observed when anti-integrins were administered to H460 cells before coculture with lung microexplants.
Impact Statement
We describe a new platform for the study of cell migration and adherence in human lung that incorporates 3D tissue structure, cellular heterogeneity, and donor variability. Using this platform, we highlighted the role of integrins in the adherence of lung cancer cells, demonstrating this method’s usefulness in the study of cancer cell adherence. This method could be used to evaluate the efficacy of therapeutics targeting the adherence and growth of cancer cells or as a platform for the general study of cell tropism and motility in 3D microexplant environments.
Introduction
Cell culture in a monolayer, the traditional method for studying biology in vitro, lacks the in vivo tissue features of interpopulation cell–cell interaction, 1 complex heterogeneity, differentiation, 2 and cell migration. 3 These shortcomings have been implicated as possible explanations for the poor correlation between the results of drug response trials in cell lines as compared with those of in vivo studies.4–5 Efforts to culture tissue in three dimensions, using models such as organoids, organs-on-chips, 6 hanging-drop microplates, 7 and spheroids 8 have sought to improve the correlation between in vitro and in vivo systems. 9 One approach which preserves these features and closely mirrors host tissue structure is that of ex vivo patient-derived explants, 10 which addresses these deficiencies by providing the relevant tissue architecture and cell heterogeneity to the highest degree possible. 11
Ex vivo culture systems have begun using small pieces of host tissue which are no greater than 1 mm in diameter. 12 Converting a patient biopsy into multiple smaller tissues, or microexplants, increases the number of possible replicates available from each tissue donor while maintaining an adequate heterogeneous population of cells.13–14 In addition, the small scale of microexplants makes them ideal for cryopreservation, providing the opportunity to store sizable quantities of samples from multiple donors in a tissue bank for large-scale simultaneous evaluation of disease-specific donor heterogeneity. 15 These advances have mitigated the primary shortcomings of ex vivo culture methods and have made them a desirable avenue for the testing of disease in humans by providing the ability to repeatedly conduct experiments with the same set of donors.15–16
By preserving the intricate architecture of complex tissues such as those found in the human lung, 15 ex vivo culture platforms are uniquely suited to answer questions contingent upon proper recapitulation of these microenvironments.17–18 As they are derived from host tissue, human microexplants are particularly well-equipped for replicating a host microenvironment in the study of cell migration and adherence,19–20 but require substrates for cell migration independent of cultured tissue in order to accurately model migratory behaviors in exogenously administered cells. 21 This can be addressed by conducting ex vivo culture of microexplants within solid support systems. We have previously generated such a system, which we refer to as a liquid-like solid (LLS) hydrogel. 22 LLS hydrogels are composed of a bed of soft granular microgel particles made from inert, charge-neutral polyacrylamide swollen in liquid media. These hydrogels support the growth and development of tissue in 3D, providing a scaffold that holds the tissues stationary and minimizes the effects of local diffusion while also allowing adequate space for the travel of cells.21–23 In this microculture environment, cell migration and chemotaxis toward adherence substrates such as the extracellular matrix of microexplants can suitably be studied. 22
Here, we report a method through which an LLS hydrogel platform can be used to observe and quantify the adherence of human non-small-cell lung cancer (NSCLC) cells to lung microexplants. This culture method was used to assess the role of integrins in the adherence of cancer cells to lung microexplants. Our results indicate that adherence to and invasion of lung tissue by these cells occurs via an integrin-dependent mechanism, with β2, β1, and αV integrin subunits responsible for adherence of H460 cells to lung microexplants. This method provides a novel template for quantification of cell migration and adherence and an ideal platform for development and effective preclinical evaluation of integrin-targeting therapeutics.
Method
LLS microgels
LLS microgels were obtained from Aurita Bioscience as mechanically disaggregated polyacrylamide, comprising particles 50–100 µm in characteristic size. LLS solutions used in experiments were centrifuged at 1,000 g for 10 min and the supernatant aspirated before the remaining volume was diluted 1:2 with lung microexplant medium (composition in Table 1) and allowed to equilibrate in a 37°C water bath for at least 30 min before use. Equilibrated LLS was centrifuged once more at 1,000 g for 10 min and all media aspirated before use in the culture of cells or tissues. For all steps involving LLS, either 200 µL or 1 mL wide bore micropipette tips and slow, gentle pipetting were used to facilitate accurate transfer of LLS at desired volumes.
List of Reagents Used in the Formulation of Lung Microexplant Medium.
Lung explant collection and microexplant cryopreservation
Whole or partial-lobe lung biopsies were retrieved from patients undergoing lung resection surgery (donor demographics in Table 2). Regions of lung tissue deemed to be free from pathology were collected in nonsupplemented RPMI-1640 and transported on ice to a biosafety cabinet, where any pleura was removed and the remaining tissue dissected and divided between the wells of a 24-well plate in 0.6 g segments. These segments were dissected into ∼1 mm diameter microexplants using histology scissors and then resuspended in 1.4 mL of a DMSO-free cryopreservation medium (CryoSOFree, Sigma-Aldrich, #C9249, St. Louis, MO, USA) before distribution in 200 µL aliquots to cryotubes containing 300 µL of CryoSOFree for a final cryopreservation volume of 500 µL. Samples were cooled overnight to −80°C at a rate of 1°C/min using a freezing container (Mr. Frosty, Fisher Scientific, #5100-0001, Waltham, MA, USA) before transfer to a dewar containing liquid nitrogen (Fig. 1A).

Overview of lung microculture procedure.
Demographics of Tissue Donors. All Known or Disclosed Comorbidities Are Indicated.
Cell lines and culture
Cryopreserved ATCC NSCLC lines NCI-H460 (human lung large cell carcinoma), NCI-H226 (human lung squamous cell carcinoma), and NCI-H441 (human lung adenocarcinoma) were obtained from a collaborator who purchased them from ATCC. All cell lines were grown in NSCLC media, composed of RPMI-1640 (Gibco, #21063-029, Grand Island, NY, USA) supplemented with 10% v/v fetal bovine serum (FBS, Atlas Biologicals, #F-0500-D, Fort Collins, CO, USA), 1% 100× penicillin–streptomycin (Corning, #30-002-CI, Corning, NY, USA), and 1% 100× nonessential amino acid mixture (Lonza, #13-114E, Walkersville, MD, USA), at 37°C under 5% CO2 to 80–90% confluence (2–4 days) before passage. For cell passage, cells were first washed with 3–5 mL of 0.25% trypsin–EDTA solution (Gibco, #25200-056) and then incubated at 37°C under 5% CO2 in 3–5 mL of the same solution for ∼6 min. The trypsin–EDTA solution was diluted 1:4 with NSCLC media, and cells were either passaged into a new flask at 50% concentration or collected in a conical tube and counted for use in experiments.
Embedding of cancer cells in LLS
Passaged cell lines were counted before centrifugation at 100g for 5 min at 25°C. Cells were resuspended in 1 mL of 1× Dulbecco’s phosphate-buffered saline (DPBS, Gibco, #10010-023) supplemented with 5% v/v lung microexplant medium and stained with either CellTracker Orange (Thermo Fisher, #C34551, Waltham, MA, USA) at a final concentration of 1 µM for microscopy experiments or CellTrace Violet (Thermo Fisher, #C34571) at a final concentration of 1 µM for flow cytometry experiments. Staining cells were incubated for 30 min in a shaking incubator set to 110 rpm and 37°C. Stained cells were washed using 14 mL of 1× DPBS, spun once more at 100 g for 5 min at 25°C, resuspended in 1 mL of lung microexplant medium, and divided into cell aliquots of 2 × 105 cells. These aliquots were spun at 100 g for 5 min at 25°C and resuspended in equilibrated LLS at a concentration of 2 × 106 cells/mL. Cell pellets were thoroughly mixed throughout the LLS by repeated gentle pipetting (Fig. 1B).
Cell and microexplant coculture for time-lapse microscopy
For time-lapse microscopy experiments, cryovials containing lung microexplants were thawed from cryopreserved stock for 3 min in a 37°C water bath before microexplants were washed twice in 15 mL of 1× DPBS and allowed to settle by gravity between washes. Lung microexplants were stained with CellTracker Deep Red (Thermo Fisher, #C34565) at a final concentration of 1 µM and incubated for 30 min in a shaking incubator set to 110 rpm and 37°C. Stained lung microexplants were washed further with 15 mL of 1× DPBS, which was aspirated as much as possible to leave a small volume of densely packed microexplants. An 18G needle was used to transfer single lung microexplants to the bottom of the wells in a 96-well plate. Cells from the H226, H441, or H460 cell line in LLS were then pipetted atop the lung microexplants to create the microcultures, followed by 100 µL of lung microexplant medium in each well. Plates were transferred to a stage-top incubator at 37°C and 5% CO2 and allowed to equilibrate to temperature for approximately 30 min and were then imaged for 48 h. Images of cell and tissue microcultures in LLS were captured on a confocal microscope (Nikon Eclipse A1R) using the 561 nm and 640 nm lasers in the NIS Elements software package during a time-lapse with the Galvano scanning mode. Images of cells alone in LLS were separately captured on a confocal microscope (Leica SP8) using a 552 nm laser in the associated LAS X software package during a time-lapse with the Galvano scanning mode. Processing of images, involving colorization and maximum intensity projection of all acquired z-stacks for each channel before merging all channels into composite images for each time point, was accomplished with the Fiji software package (Version 2.9.0, on macOS Ventura 13.4.1 by Apple, Cupertino, CA, USA). Colorization choices for each image are arbitrary and no signals were deleted, overwritten, or otherwise modified during processing steps.
Cell and microexplant coculture for flow cytometry analysis
For each donor in flow cytometry experiments, one cryovial containing lung microexplants was thawed from cryopreserved stock for 3 min in a 37°C water bath before microexplants were washed twice in 15 mL of 1× DPBS, allowed to settle by gravity between washes. DPBS was aspirated to leave ∼250 µL of densely packed microexplants, from which 100–200 microexplants (∼60 µL) were transferred to the LLS containing stained cells at a concentration of 2 × 106 cells/mL, and pipette-mixed. The LLS mixture of stained cells and microexplants was pipetted into the wells of a 96-well plate, followed by 100 µL of lung microexplant medium atop the culture mixture in each well. Plates were incubated for 48 h at 37°C in a 5% CO2 incubator, each culture gently treated with 100 µL of additional lung microexplant medium at 24 h. At the conclusion of the culture period, microcultures from the plate were individually pipetted atop a 400 µm cell strainer (pluriSelect, #43–50400-01, Leipzig, Germany) mounted in a 50 mL tube and each gently washed with 15 mL of 1× DPBS to remove any nonadherent cells from the microexplants. A 200 µL aliquot of 1× DPBS was used to recover and transfer microexplants from the cell strainer into a 15 mL tube prepared for each culture well. Each tube of microexplants then received 1 mL of RPMI 1640 treated with 30 units/mL DNase (Sigma-Aldrich, #11284932001) and 0.5 mg/mL Liberase (Sigma-Aldrich, #5401127001) before all tubes were incubated in a shaking incubator set to 37°C and 110 rpm for 1 h. Digested microexplants were triturated 30 times in a 1 mL syringe equipped with a 1.5” 18G blunt-ended needle, washed with 2 mL of flow cytometry buffer (1X DPBS with 2% FBS and 1 mM EDTA), and filtered through 100 µm Nitex mesh into flow cytometry tubes. Tubes were spun at 400 g at room temperature for 5 min, the supernatant was decanted, and the cells were gently resuspended before 10 µL of SYTOX AADvanced Ready Flow Reagent (Invitrogen, #R37173, Waltham, MA, USA) was added to each tube. Samples were incubated a further 10 min in a shaking incubator at 37°C and 110 rpm and run at a rate of 30 µL/min for 3 min or until empty on a Beckman Coulter CytoFLEX using the forward scatter (FSC), side scatter (SSC), 450/45 nm band pass (PB450), and 660/10 band pass (APC) channels. Gating strategy for isolation of single cells can be found in Supplementary Figure S1. Data were collected using CytExpert (Version 2.4, Brea, CA, USA; on Windows 10 by Microsoft, Redmond, WA, USA) and gated using FlowJo (Version 10.9.0, Ashland, OR, USA; on macOS Ventura 13.4.1).
Experiments
Integrin blockade of H460 cells
H460 cells were prepared and stained for flow cytometry as indicated above. After the staining period but before coculture with lung microexplants, H460 cells at a concentration of 2 × 106 cells/mL were incubated in media containing 20 µg/mL of anti-CD18 (BioLegend, #363402, San Diego, CA, USA), anti-CD29 (BioLegend, #921304), and anti-CD51 (BioLegend, #327902). Approximately 4 × 108 of each antibody was present per cell. 24 The blockade was carried out for 20 min in a shaking incubator set to 37°C and 110 rpm. A control sample of cells containing the same concentration of anti-mouse IgG (BioLegend, #406602) was also prepared and incubated in the same manner. Antibody-treated cells were then combined with lung microexplants to create microcultures which were evaluated qualitatively by confocal microscopy and quantitatively by flow cytometry. Cultures incubated for flow cytometry received additional media containing 10 µg/mL of blocking agents or control IgG after 24 h of culture. Technical duplicates were used for each tested donor in each experiment, and the average of duplicates from four experiments was used in analysis, with the same instrument parameters, settings, and gating applied to each experiment.
Statistical analysis
Calculations involving collected data were performed in Numbers (Version 13.1, on macOS Ventura 13.4.1) and statistical analysis was conducted in Prism (Version 10.0.1, San Diego, CA, USA; on macOS Ventura 13.4.1). Because lung microexplant culture wells contained only an approximately similar number of microexplants to one another, and each microexplant is inherently variant in terms of its size and number of component cells, adherence was measured as a ratio of viable, recovered H460 cells to the total number of viable cells, and normalized to a count of H460 cells per 10,000 viable cells. A two-tailed, ratio-paired t test was used for statistical analysis, and a threshold p value of 0.05 was designated for all studies.
Discussion
We have previously characterized the cellular composition, 3D structure, and virally induced inflammatory response profile of cryopreserved lung microexplants cultured in media and analyzed by flow cytometry, microscopy, and qPCR. After thawing, cryopreserved lung microexplants remain viable in culture for up to 96 h, consistently retaining differentiated epithelial cell, endothelial cell, leukocyte, and monocyte populations in quantities consistent with fresh tissue. Epithelial cell tight junctions within the cryopreserved microexplants are preserved in three dimensions throughout the culture period, and microexplants infected with SARS-CoV-2 respond by producing significant quantities of GM-CSF, IL-1β, IL-10, CXCL10, and IFN-α2. 15 These analyses were performed in liquid media, which lacks a structural network for cell migration to occur. 21 To adapt this system for the study of cell migration and adhesion to microexplants, a 3D solid support network in the form of LLS was incorporated.
The use of LLS in our microexplant culture system provides a matrix through which individual cells can migrate while simultaneously supporting and maintaining the delicate architecture of the lung microexplants, so that cell–tissue interactions can be observed over the duration of the 48 h culture period. We have previously studied the migration of highly invasive murine glioblastoma tumoroid cells through LLS microgels, noting the porous semisolid nature of LLS as affording the necessary space for cell migration and diffusion of nutrients while still providing responsive tactile feedback to traveling cells. 21 The work presented here expands upon that premise by including primary human tissue within the LLS matrix, providing a basis for the study of interactions between migratory cells and tissue in three dimensions. We selected three NSCLC cell lines representing the most common types of NSCLC 25 to study migration within a relevant tissue architecture provided by our cryobank of primary human lung tissue.
In our study, time-lapse microscopy allowed for the visualization of NSCLC cell migration and adherence to lung microexplants in all three tested NSCLC lines, captured in still images in Figure 2A–C, and in video form in Supplementary Video S1, Supplementary Video S2 and Supplementary Video S3. In each cell line, initial adherence to lung microexplants was observed within the first hour of the culture period. Adherent cells would either remain in place or migrate along the surface of the microexplants throughout the culture period but largely remained affixed to the lung microexplants once adhered. Migration of cells from each cell line through the LLS to the lung microexplants was also observed over the duration of the culture period. Movement into and along the lung microexplants was observed in all cell lines. Stained cells from each line gradually lost fluorescence over time, which we attribute to cellular metabolism and the natural dilution of intracellular stains by cell division, but remained motile for the duration of the culture period (Supplementary Video S4 and Supplementary Video S5). The time-lapse visualization presented in Supplementary Video S1, Supplementary Video S2 and Supplementary Video S3 made clear the adherent capabilities of multiple cancer cell lines within singular microexplants and prompted our efforts to quantify this adherence across multiple microexplants cultured simultaneously in a single well.

Regions of interest from time-lapse confocal microscopy of non-small-cell lung cancer (NSCLC) cell lines. Lung microexplants are stained with CellTracker Deep Red (blue), whereas NSCLC cells are stained with CellTracker Orange (red). Times indicated represent the period elapsed after imaging was commenced. Squamous cell carcinoma cells (H226) are shown in
Adherence capacity of migratory cancer cells to lung microexplants was thus quantified using flow cytometry. Four experiments were performed, each comprised of technical duplicates for four lung donors, LD35, LD36, LD37, and LD38. Flow cytometry signals were gated, and cells identified according to their FSC and SSC profile (Supplementary Fig. S1). Viable cells were gated as SYTOX AADvanced negative in the APC channel based on an unstained control (Fig. 3A, 3B). A gate for viable, microexplant-adherent, CellTrace Violet-stained H460 cells in the PB450 channel was derived by comparison of cultures containing only lung microexplants, as an FMO control (Fig. 3C), with cocultures of lung microexplants and stained H460 cells (Fig. 3D). Samples which received stained H460 cells had a significantly larger population of cells in the PB450 gate than those which did not, in every donor, as compared using a ratio-paired t test (p < 0.01, Fig. 3E, Supplementary Fig. S2). On average, samples which contained only lung microexplants had less than one viable cell signal in the CellTrace Violet gate per ten thousand recorded viable cell signals, indicating the robustness of this method for detection of dyed, adherent H460 cells. Now able to measure a rate of migratory cell adherence, our attention turned to mechanisms governing this process.
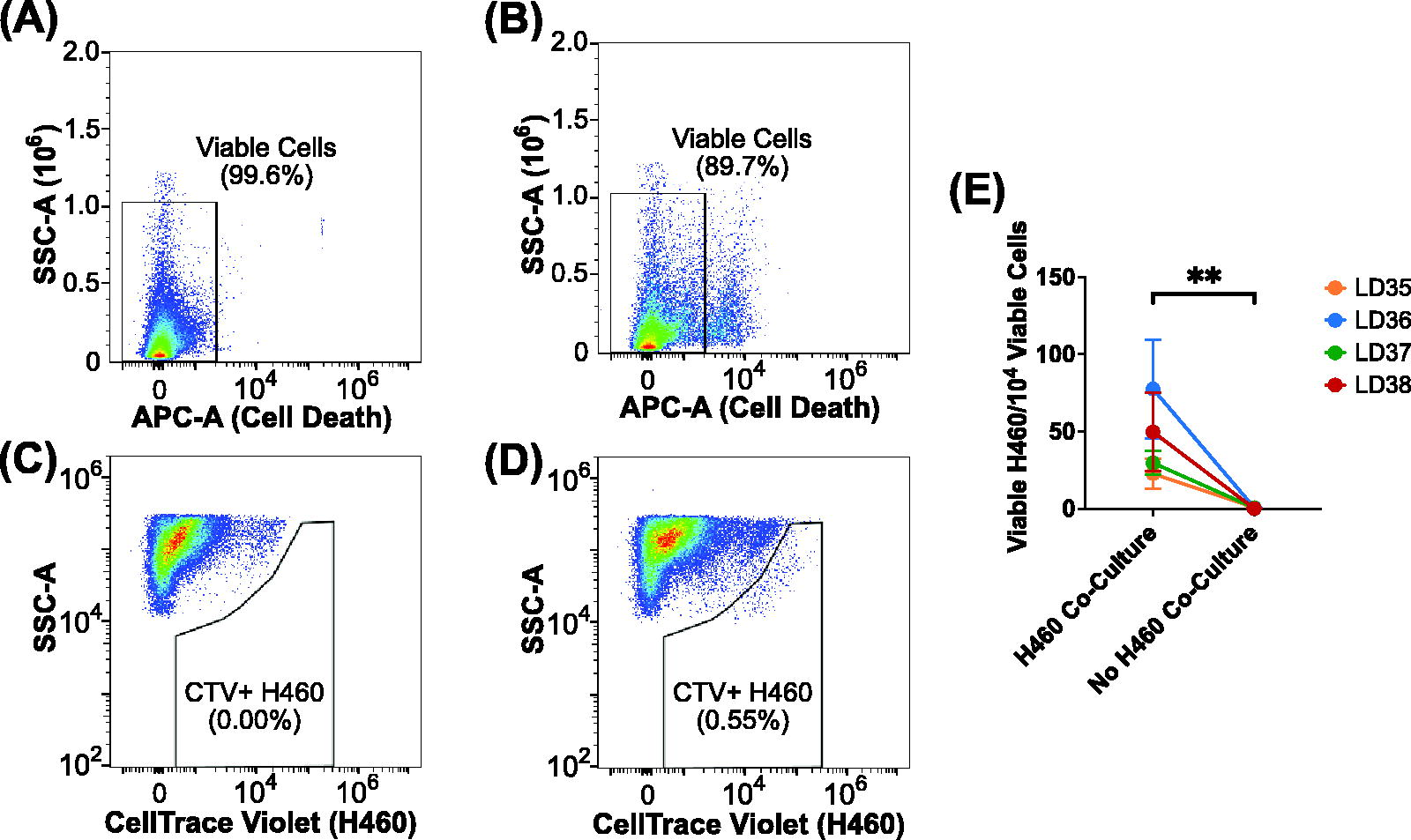
Flow cytometric analysis of large cell carcinoma (H460) cell adherence to lung microexplants following coculture, filtration, and sample digestion.
Cellular adhesion is primarily regulated by integrins, 26 noncovalently heterodimeric cellular adhesion receptors 27 which are rarely mutated in cancers 28 and directly responsible for cell traction in tumor invasion. 29 The integrin family of proteins includes 18 α and 8 β subunits, α-and-β pairs of which recognize extracellular collagen, laminin, and Arg-Gly-Asp (RGD) motif-containing proteins. 30 Increased expression of integrins generally correlates with poor prognosis in invading cancers,31–32 with integrin subunits β2, β1, and αV particularly important in colonization of tissues through recognition of critical extracellular matrix proteins 28 and VCAM-1 on the surface of endothelial cells. 33 As integrins are critical to adherence of tumor cells, 29 they are a desirable therapeutic target in cancer treatment. To test the suitability of the LLS microexplant culture environment as a platform for evaluation of adhesive factors expressed on cancer cells, an antibody blockade of H460 cell-surface integrins in the active binding conformation was performed. The proteins CD18, CD29, and CD51 (integrin subunits β2, β1, and αV, respectively) were chosen as candidates for the blockade because they are essential to integrin heterodimers which regulate nearly all binding interactions between cells and the laminins, collagens, and RGD-containing peptides of the extracellular matrix.28,34 Because subunits of an integrin heterodimer must be in the active conformation and available for the integrin to successfully bind its target, 35 we hypothesized that the availability of these three integrin heterodimers on the surface of H460 cells was necessary for the adherence of these cells to lung microexplants; therefore blocking them would almost completely abrogate the ability of the H460 cells to adhere to the extracellular matrix of the microexplants.
Using the described method, we observed by confocal fluorescence microscopy that a blockade of CD18 (β2 integrin), CD29 (β1 integrin), and CD51 (αV integrin) yielded a reduction in adherent H460 cells to lung microexplants compared against H460 cells which did not receive an integrin blockade (Fig. 4A). This effect was confirmed quantitatively in flow cytometric analysis, in which integrin-blocked, CellTrace Violet-stained H460 cells cocultured with lung microexplants demonstrated a smaller adherent population compared with H460-microexplant cocultures that did not receive an integrin blockade (Fig. 4B, 4C). To confirm this effect, this experiment was repeated across four donors, with four experimental replicates each. The results of these experiments indicated a significant reduction in the adherence capacity of integrin-blocked H460 cells to primary human lung microexplants, as compared with H460 cells which did not receive an integrin blockade (Fig. 4D, Supplementary Fig. S2).
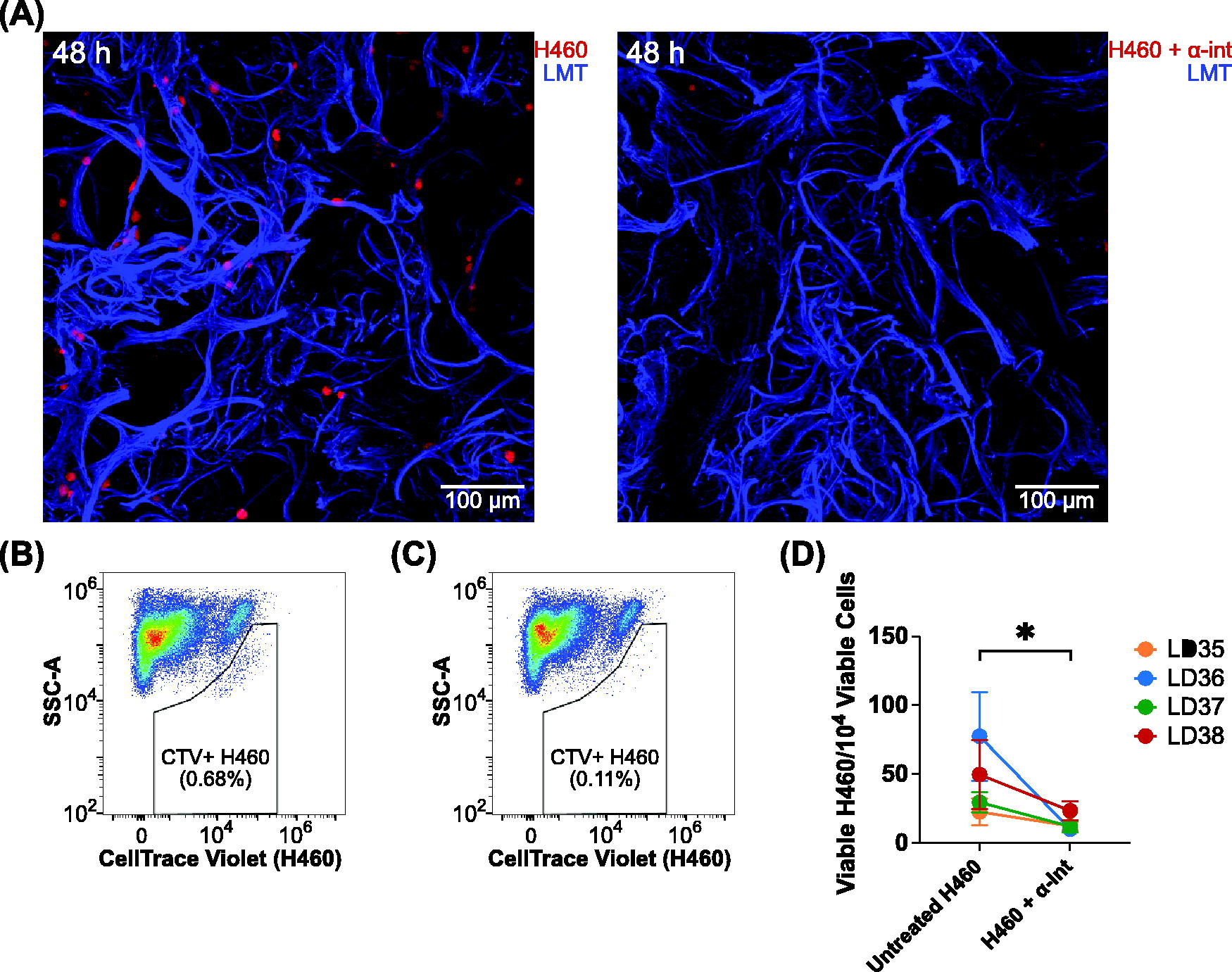
Administration of antiintegrins to large cell carcinoma (H460) cells abrogates adherence to lung microexplants.
We used the advantage of reproducibility and replicability across multiple human subjects afforded to us by our cryobank and the utility of the method described here to study the importance of integrin availability in human large cell carcinoma adherence to human lung microexplants. Our results corroborate established literature indicating that integrins play a critical role in the adherence of cancer cells to tissues36–37 in the process of migration as well as metastasis38–39 and provides a new way to evaluate treatments targeting these mechanisms in multiple donors of human cells and tissues. Our study demonstrates the ability of our cryobank to observe potential effects related to donor heterogeneity and identifies this method as a suitable platform for the testing of therapeutics targeting the adherence of cancer cells, particularly via integrin-related mechanisms.
We have previously reported the utility of ex vivo lung microexplant cryopreservation as a basis for the study of antiviral agents 15 and the use of LLS microgels to observe cell migration. 21 This work combines these concepts to describe a robust method for the study of cell tropism, migration, and adherence within the context of primary human tissue. Apart from its obvious potential to enhance the accuracy of therapeutics created to target cell migration and adherence, this system could also be used to study tumor formation, observe immune cell migration, or evaluate host–pathogen interactions dependent on human host tissue architecture or cellular heterogeneity in a wide variety of tissues. Considering these factors in the context of the enhanced reproducibility and replicability in human subjects offered by a cryobank, this system is an improvement over traditional cell line monolayer culture and presents a new frontier in the use of 3D and ex vivo culture systems for the study of human disease and the development of cancer therapeutics.
Footnotes
Acknowledgments
The authors would like to thank all the donors who graciously consented to the collection of their tissue for use in our experiments.
Authors’ Contributions
N.G.D.: Conceptualization (equal), data curation (lead), formal analysis (lead), investigation (lead), methodology (lead), project administration (lead), resources (lead), validation (lead), visualization (lead), and writing—original draft (lead). Z.E.D.: Resources (supporting) and validation (supporting). F.T.L.: Resources (supporting). J.F.: Methodology (supporting) and resources (supporting). R.A.S.: Methodology (supporting) and resources (supporting). G.Q.: Resources (supporting). Y.G.: Resources (supporting). D.T.N.: Methodology (supporting) and resources (supporting). W.G.S.: Funding acquisition (equal) and resources (supporting). E.A.P.: Resources (supporting). B.M.: Conceptualization (supporting), formal analysis (supporting), funding acquisition (equal), methodology (supporting), resources (supporting), supervision (supporting), and writing—review and editing (equal). M.A.S.: Conceptualization (equal), formal analysis (supporting), funding acquisition (equal), methodology (supporting), resources (supporting), supervision (lead), and writing—review and editing (equal).
Disclosure Statement
No conflicts of interest exist.
Funding Information
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.