
Abstract
Transforming growth factor beta (TGF-β) is a potent growth factor that regulates the homeostasis of native cartilage and is administered as an anabolic supplement for engineered cartilage growth. The quantification of TGF-β activity in live tissues in situ remains a significant challenge, as conventional activity assessments (e.g., Western blotting of intracellular signaling molecules or reporter cell assays) are unable to measure absolute levels of TGF-β activity in three-dimensional tissues. In this study, we develop a quantification platform established on TGF-β’s autoinduction response, whereby active TGF-β (aTGF-β) signaling in cells induces their biosynthesis and secretion of new TGF-β in its latent form (LTGF-β). As such, cell-secreted LTGF-β can serve as a robust, non-destructive, label-free biomarker for quantifying in situ activity of TGF-β in live cartilage tissues. Here, we detect LTGF-β1 secretion levels for bovine native tissue explants and engineered tissue constructs treated with varying doses of media-supplemented aTGF-β3 using an isoform-specific ELISA. We demonstrate that: 1) LTGF-β secretion levels increase proportionally to aTGF-β exposure, reaching 7.4- and 6.6-fold increases in native and engineered cartilage, respectively; 2) synthesized LTGF-β exhibits low retention in both native and engineered cartilage tissue; and 3) secreted LTGF-β is stable in conditioned media for 2 weeks, thus enabling a reliable biological standard curve between LTGF-β secretion and exposed TGF-β activity. Accordingly, we perform quantifications of TGF-β activity in bovine native cartilage, demonstrating up to 0.59 ng/mL in response to physiological dynamic loading. We further quantify the in situ TGF-β activity in aTGF-β-conjugated scaffolds for engineered tissue, which exhibits 1.81 ng/mL of TGF-β activity as a result of a nominal 3 μg/mL loading dose. Overall, cell-secreted LTGF-β can serve as a robust biomarker to quantify in situ activity of TGF-β in live cartilage tissue and can be potentially applied for a wide range of applications, including multiple tissue types and tissue engineering platforms with different cell populations and scaffolds.
Impact Statement
The quantification of transforming growth factor beta (TGF-β) activity in live tissue has remained a significant challenge. Here, we propose a TGF-β activity measurement platform using its autoinduction response, where the proportional relation between levels of cell-secreted latent TGF-β (LTGF-β) and exposed TGF-β activity serves as a standard curve. As such, LTGF-β secretion monitoring can allow for enhanced quantitative characterizations of endogenous TGF-β activity regulation in native tissues and optimization of TGF-β delivery systems for regenerative medicine. This platform may be compatible with a wide array of ex vivo native tissue systems and regenerative medicine platforms with varying cell sources and biomaterial scaffolds.
Introduction
Transforming growth factor beta (TGF-β) is a multifunctional signaling molecule that plays critical roles in the development, growth, and maintenance of articular cartilage. 1 Research into the chondrogenic effects of TGF-β has largely focused on two areas: its role in the development and maintenance of native synovial joint tissues2–5 and its role as an anabolic supplement for cartilage tissue engineering.6–8 In both realms, the regulation of TGF-β is critical—TGF-β insufficiencies lead to native cartilage degeneration 9 and inadequate engineered cartilage development,6,8 whereas TGF-β excesses are associated with synovial joint pathology10–12 and engineered cartilage growth abnormalities.13–16 Thus, quantifying TGF-β activity levels in cartilage tissues can advance our understanding of TGF-β regulatory mechanisms in native cartilage and guide the development of TGF-β delivery protocols in cartilage tissue engineering.
In native cartilage, TGF-β activity is tightly regulated, in a manner that promotes cellular biosynthesis while avoiding TGF-β-excess-associated pathology. 17 Native cartilage possesses large amounts of TGF-β that exist bound to the extracellular matrix (ECM) in an inactive form, termed latent TGF-β (LTGF-β). 18 To bind to chondrocyte receptors and elicit a biological response, TGF-β must first undergo release from this complex in a process termed TGF-β activation18,19 (Fig. 1A). Recent work has uncovered the important role of physiological mechanical loading of cartilage in activating LTGF-β,25,26 pointing toward the existence of unique mechanobiological feedback control, whereby tissues may exhibit elevated biosynthesis in response to joint activity through TGF-β-mediated signaling. However, quantitative measures of the rate of TGF-β activation in response to mechanical stimuli have yet to be performed, thus limiting our understanding of the physiological implications of these mechanisms.

Schematic of transforming growth factor beta (TGF-β) delivery in native and engineered cartilage and TGF-β autoinduction response.
For cartilage tissue engineering, TGF-β is a widely utilized supplement owing to its ability to promote chondrogenesis and biosynthesis of cartilaginous ECM constituents.27–29 The past decade has seen the emergence of a multitude of scaffold-based delivery platforms that can achieve sustained delivery of TGF-β, 27 such as TGF-β affinity scaffolds, 20 TGF-β releasing microspheres, 21 and direct scaffold conjugation of TGF-β22–24 (Fig. 1B). Previous studies have demonstrated the critical role of TGF-β doses in neocartilage development, where insufficient TGF-β may generate inferior tissue functional properties,6,8 and excessive TGF-β can lead to hypertrophy or fibrocartilage-like tissue growth.13–15 However, quantitative measures of TGF-β cell exposure profiles are typically not performed, thus serving as an impediment for developing platforms that achieve optimal TGF-β delivery outcomes.
Quantifying TGF-β activity in live tissues remains a significant challenge. Activated TGF-β exhibits a short half-life in situ, 30 owing to its characteristic rapid clearance from the extracellular space via receptor internalization/degradation kinetics. 31 As a result, TGF-β activity characterizations typically rely on assessments of downstream signaling events, such as immunostaining or Western blotting of phosphorylated intracellular TGF-β signaling proteins (e.g., p-SMAD2/3),26,32–34 or qPCR of TGF-β-inducible genes. 25 These assessments are limited, as they are sample destructive and may exhibit low sensitivity for quantifying in situ levels of TGF-β activity in a tissue. Reporter cell assays can provide quantitative metrics35–37 but are generally not compatible with three-dimensional tissue specimens. For TGF-β delivery scaffolds in tissue engineering applications, TGF-β delivery profiles are conventionally assessed via measures of TGF-β desorption from the scaffold.21,23,32,34 However, these assessments may provide limited insights, as levels of in situ TGF-β activity to embedded cells may be considerably different from desorption profiles.
A notable characteristic of TGF-β is its well documented autoinduction behavior.38,39 Active TGF-β (aTGF-β) signaling in cells induces biosynthesis and secretion of new TGF-β in its latent form (LTGF-β). Cell-secreted LTGF-β is highly stable, making it an attractive biomarker for the quantification of aTGF-β activity acting on cells. As such, rates of LTGF-β secretion from cartilage explants and tissue constructs into culture media may reflect in situ aTGF-β activity levels in the tissue. LTGF-β secretion assessments in conditioned media for tissues with unknown TGF-β activity can be performed in conjunction with quantitative live tissue standards that relate LTGF-β secretion rates to administered aTGF-β activity levels, allowing for fully quantitative assessments of in situ TGF-β activity (Fig. 1C). Furthermore, one can confirm that LTGF-β secretion enhancements result from aTGF-β activity, as opposed to other signaling cascades, via use of TGF-β receptor kinase inhibitors, which specifically block TGF-β signaling.
In this study, we examined the utility of TGF-β autoinduction measures to quantify TGF-β activity in cartilage research applications. First, we assessed the feasibility of the platform by: 1) confirming that newly synthesized LTGF-β is predominantly secreted into culture media, rather than retained in the ECM; 2) establishing the quantitative relationship between aTGF-β exposure doses and LTGF-β secretion levels in cartilage tissues; and 3) validating the specificity of TGF-β inhibitors for suppressing TGF-β-mediated increases of LTGF-β synthesis. Second, we utilized TGF-β autoinduction measurements to quantify in situ TGF-β activity in two research applications: 1) quantifying in situ levels of endogenous aTGF-β activity in cartilage explants in response to physiological mechanical loading and 2) quantifying in situ levels of exogenous aTGF-β activity in engineered cartilage tissue constructs generated with TGF-β-conjugated scaffolds.
Materials and Methods
Cartilage explants and engineered tissue constructs
Cartilage explants were harvested from 2-month-old bovine femoral condyles. The superficial and middle zone was excised, yielding cylindrical disks (∅3 × 2 mm) of predominantly deep zone cartilage. Chondrocyte-seeded tissue constructs (∅4 × 2 mm) were fabricated via embedding of bovine chondrocytes (2-month-old from carpometacarpal joints) in 2% w/v agarose (type VII, Sigma) at 30 × 106 cells/mL. For TGF-β scaffold conjugation, human recombinant aTGF-β3 (R&D Systems) was conjugated to methacrylate agarose (MeAgr) 40 (Supplementary Materials). All tissues were cultured in chondrogenic media (DMEM supplemented with 1% ITS+ premix, 100 nM dexamethasone, 1 mM sodium pyruvate, 50 µg/mL L-proline, 1% PS/AM antibiotic-antimycotic, and 50 µg/mL ascorbate-2-phosphate). The culture medium to tissue volume ratio was 40:1.
Assessments of autoinduction-induced LTGF-β secretion levels
Conditioned media from explants and constructs were collected and assayed for cell-secreted LTGF-β1 concentrations via an isoform-specific TGF-β1 ELISA (Duoset, R&D Systems). All media samples were subjected to acid activation to convert LTGF-β1 to the ELISA-reactive active form. 13 Secretion rates are reported as the mass of LTGF-β1 in conditioned medium normalized to the volume of the tissue specimen per units of time.
TGF-β autoinduction: Engineered constructs
Constructs were exposed to media-supplemented aTGF-β3 at 0, 0.1, 0.3, 1, 3, or 10 ng/mL for 14 days with the media being replenished every 3 days. Conditioned media were assayed for the secretion rate of the LTGF-β1 isoform to avoid cross talk with supplemented aTGF-β3.
TGF-β autoinduction: Cartilage explants
Explants were exposed to media-supplemented aTGF-β3 at 0, 0.1, 0.3, 1, 3, or 10 ng/mL for 10 days with the media replenished every other day. The stability of explant-secreted LTGF-β1 in conditioned media after explant removal was monitored at 37°C over 2 weeks. The specificity of the TGF-β-receptor kinase inhibitor, LY364947, on inhibiting LTGF-β secretion increase in response to aTGF-β but not to stimulation with other growth factors was further assessed. Here, LTGF-β1 secretion rates were determined in response to aTGF-β3, FGF-2, IGF-1, PDGF-BB, and BMP-7 (R&D Systems) at 1 or 10 ng/mL and in the absence or presence of LY364947.
LTGF-β tissue retention assessments
The relative fractions of newly synthesized LTGF-β1 secreted into conditioned media and retained in the tissue were determined for constructs and explants. Freshly cast constructs were exposed to 0 or 3 ng/mL aTGF-β3. After 2, 4, and 6 days of exposure, conditioned media were assayed for LTGF-β1 secretion levels and constructs were assayed for levels of LTGF-β1 retained in the tissue. Tissue levels of LTGF-β1 were determined via guanidine hydrochloride extraction and ELISA measure.
41
At each time point, the LTGF-β1 secretion fraction was computed as
Cartilage explant retention analysis was modified to account for the significant levels of LTGF-β1 initially bound within the ECM. Here, explants were initially diametrically halved; one half was cultured in the absence of TGF-β (-TGF-β), whereas the other half was cultured with 10 ng/mL aTGF-β3 (+TGF-β). After 3 or 7 days of culture, the explant halves were assayed for ECM-bound LTGF-β1, and conditioned media were assayed for secreted LTGF-β1. Additional explant halves were assayed at day 0 for the initial ECM-bound content of LTGF-β1.
Quantification of load-induced TGF-β activity in cartilage explants
Cartilage explants were subjected to dynamic loading regimens with a 10% platen-to-platen compressive strain at a frequency of 0.5 Hz or 2 Hz for 15 h. An additional group of explants was maintained unloaded under free swelling conditions. For all groups, explants were maintained in the absence or presence of LY364947 to reveal potential LTGF-β1 secretion increases that result from TGF-β-independent signaling pathways. Media were replenished daily, and LTGF-β1 secretion in conditioned media was assayed for 3 days after loading. For each explant, the LTGF-β1 secretion rate was converted to in situ TGF-β activity levels via an autoinduction standard curve generated by measuring LTGF-β1 secretion from a separate batch of explants in response to media-supplemented aTGF-β3 (0 to 3 ng/mL range) administered for the 15-h loading period.
Quantification of TGF-β activity in aTGF-β-conjugated constructs
aTGF-β3 was conjugated to MeAgr at 3 µg/mL (aTGF-Conj-High) or 0.3 µg/mL (aTGF-Conj-Low) (Supplementary Materials). Constructs from both groups were maintained in the absence or presence of LY364947. Furthermore, to determine the importance of aTGF-β3 scaffold conjugation for promoting in situ TGF-β activity, additional groups were utilized where the aTGF-β3 doses were loaded into nonfunctionalized agarose to achieve delivery in a soluble form (aTGF-β-Sol-High and aTGF-β-Sol-Low). Media were replenished every other day for 8 days, and conditioned media were assayed for the LTGF-β1 secretion rate. For each construct, the LTGF-β1 secretion rate was converted to in situ TGF-β activity levels via an autoinduction standard curve generated by measuring LTGF-β1 secretion from a separate batch of constructs in response to media-supplemented aTGF-β3 (0–10 ng/mL range).
Quantification of TGF-β activity via Western blotting
Constructs were exposed to 0, 0.1, 0.3, 1, 3, or 10 ng/mL aTGF-β3 for 1 h. Subsequently, intracellular proteins were extracted via incubation in protein extraction buffer with complete protease inhibitors (Roche) and phosphatase inhibitors (Pierce). Protein extracts were subjected to SDS-PAGE followed by transfer to PVDF membranes. Membranes were blocked by 5% milk; incubated in primary antibodies for p-SMAD2 (18338, Cell Signaling), total SMAD2 (t-SMAD2, 3122S, Cell Signaling), or GAPDH (2118, Cell Signaling); and subsequently incubated in antirabbit secondary antibody (7074, Cell Signaling). Immunoblots were probed using the ECL detection system (ThermoFisher). Band intensity was quantified using ImageJ, and p-SMAD2 level was normalized to GAPDH values.
Statistical analysis
One-way analyses of variance (ANOVAs; α = 0.05) were performed to determine the effect of: 1) aTGF-β3 doses on p-SMAD2 expression (Fig. 3B) and 2) culture time on the LTGF-β1 stability (Fig. 4C). Two-way ANOVAs (α = 0.05) were performed to determine the effect of: 1) aTGF-β3 exposure and culture time on secreted LTGF-β1 retention in tissue constructs (Fig. 2D) and explants (Fig. 5A) and LTGF-β1 level secreted from explant halves (Fig. 5B), 2) growth factor doses and LY364947 supplementation on explant LTGF-β1 secretion rates (Fig. 4D–H), 3) loading and culture time on explant LTGF-β1 secretion rate (Fig. 6B) and in situ TGF-β activity (Fig. 6C), and 4) culture time and delivery technique (conjugated vs. soluble) on construct LTGF-β1 secretion rate (Fig. 7B) and in situ TGF-β activity (Fig. 7C). Tukey’s HSD post hoc tests were used to examine differences in means.
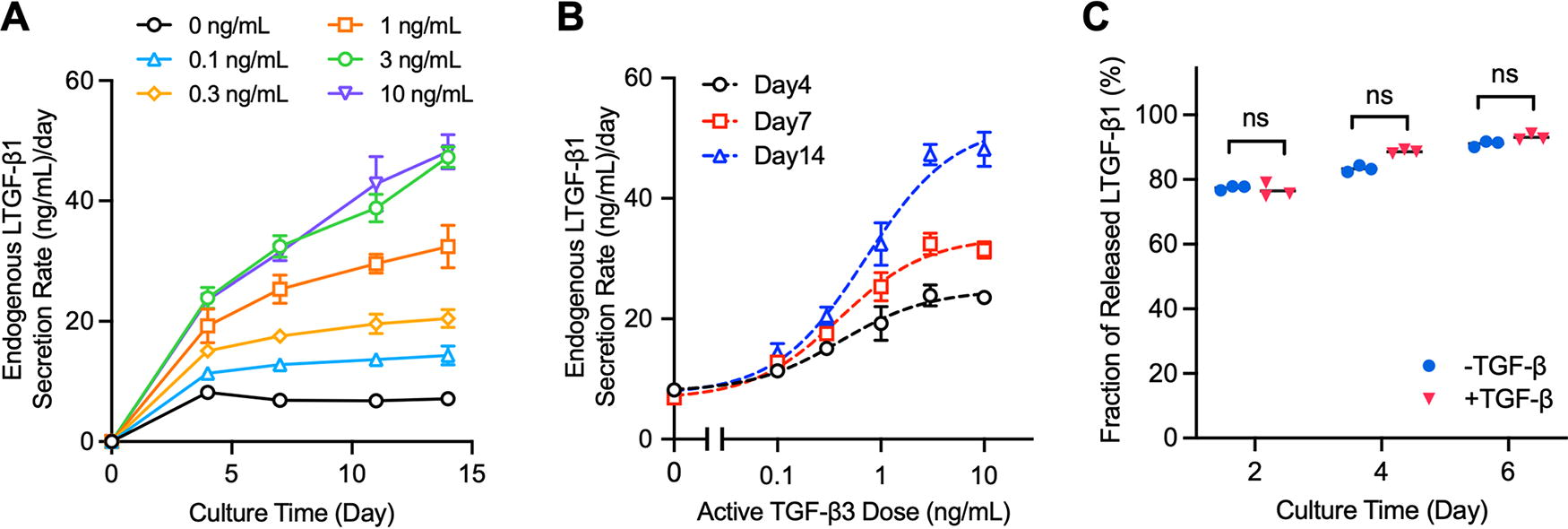
TGF-β autoinduction in engineered constructs.

Western blotting on p-SMAD2 for varying doses of aTGF-β.
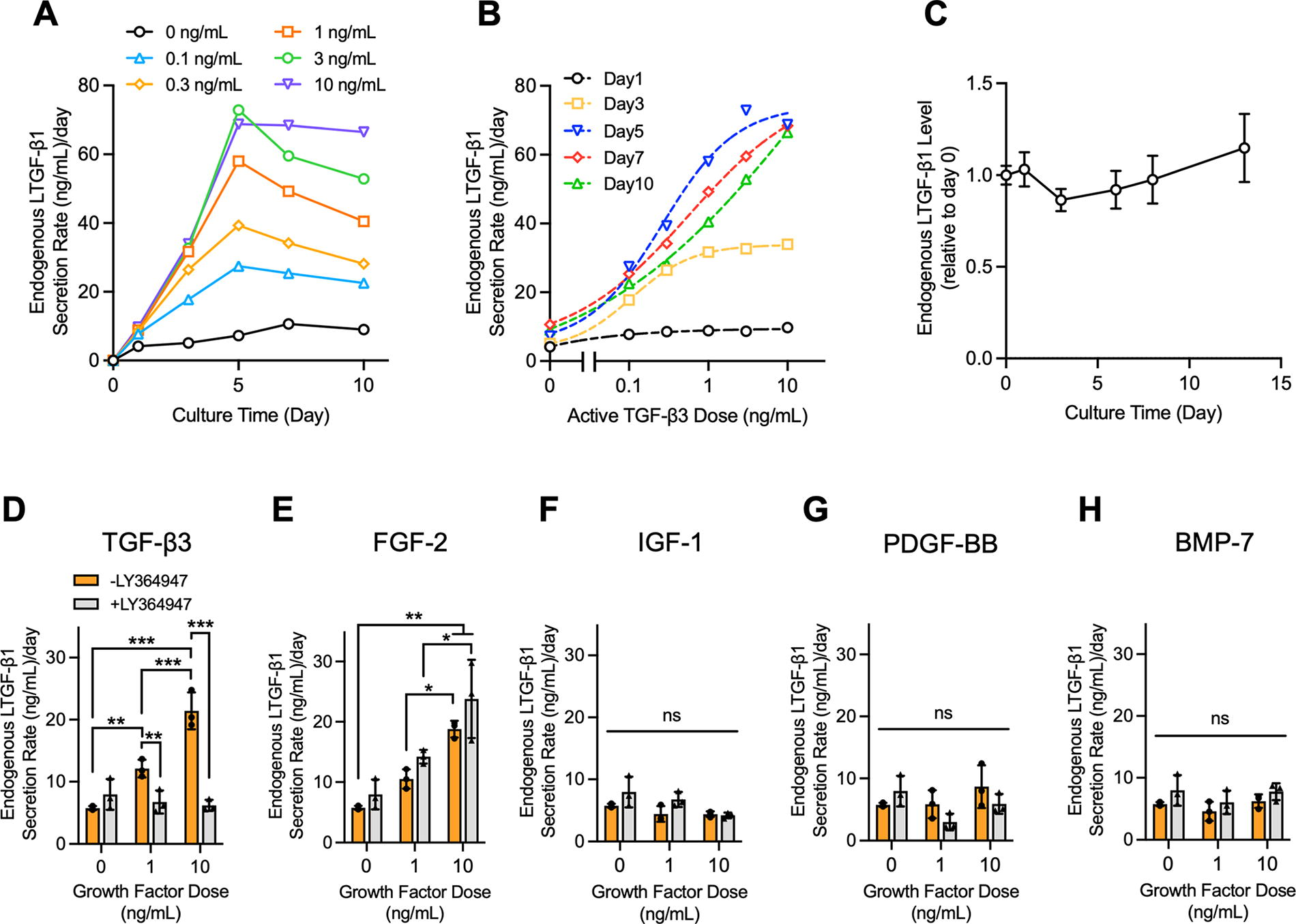
TGF-β autoinduction in cartilage explants.

Retention of secreted LTGF-β1 in cartilage explants.
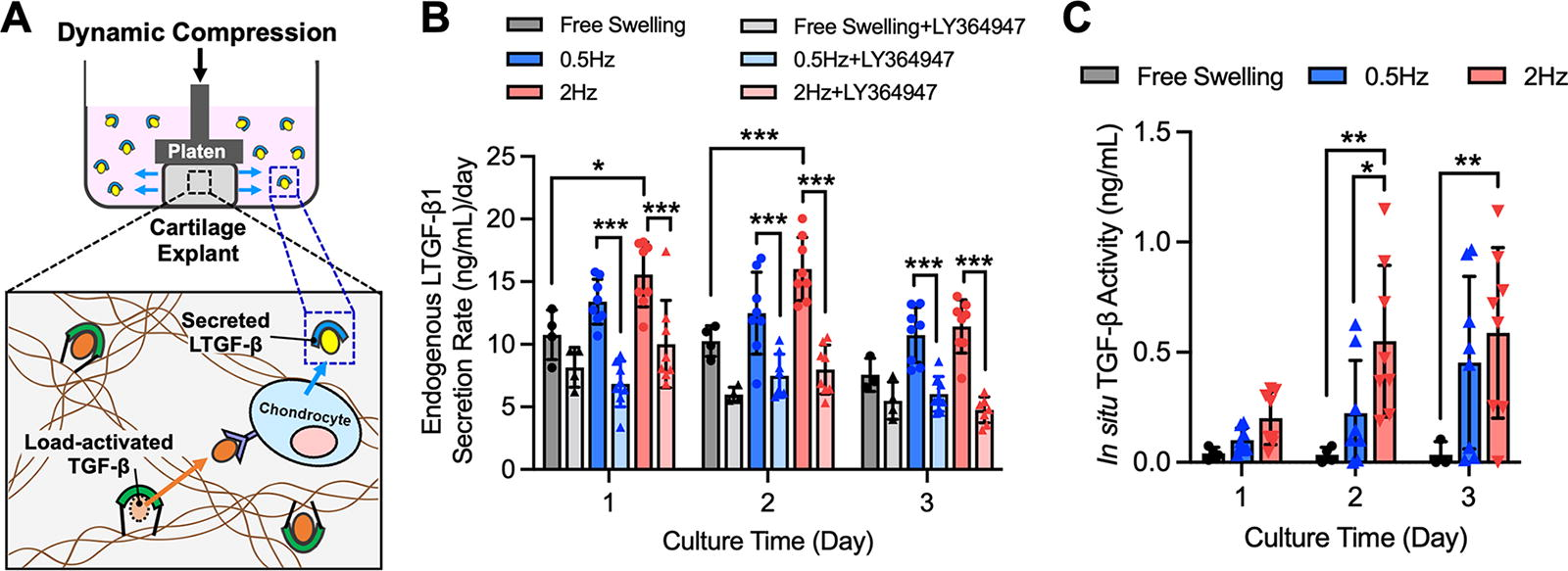
In situ TGF-β activity in dynamically loaded cartilage explants.

In situ TGF-β activity in aTGF-β-conjugated engineered constructs.
Experiment
TGF-β autoinduction: Engineered constructs
The quantitative relationship between aTGF-β exposure levels and LTGF-β secretion rates in engineered constructs was determined. In the absence of exogenous aTGF-β3, constructs exhibited a low basal rate of LTGF-β1 secretion (7.2 ± 0.8 ng/mL/day on average over 14 days) (Fig. 2A). LTGF-β1 secretion rates increased with aTGF-β3 dose and continued to increase further over time; standard curves were generated (Fig. 2B). Increases plateaued at 3 ng/mL, reaching a 6.6-fold increase above 0 ng/mL levels at 14 days of culture. Upon removal of aTGF-β3, LTGF-β1 secretion rates began to decrease (Supplementary Fig. S1). In addition, only a small fraction of newly synthesized LTGF-β1 was retained in the construct; 74%−92% of synthesized LTGF-β1 was secreted into surrounding media (Fig. 2C).
The sensitivity of autoinduction assessments of TGF-β activity was compared to assessments using quantitative Western analysis of SMAD2 phosphorylation. Results showed that p-SMAD2 intensity increased with aTGF-β exposure (Fig. 3A, B), but there was no significant difference in band intensity between 0.1 ng/mL and 10 ng/mL doses (p > 0.5), indicating a lower sensitivity to aTGF-β than autoinduction measures. t-SMAD2 level exhibited a moderate decrease with aTGF-β supplementation (Fig. 3A), consistent with prior reports. 42
TGF-β autoinduction: Cartilage explants
The quantitative relationship between aTGF-β exposure levels and LTGF-β secretion rates in cartilage explants was determined. In the absence of exogenous aTGF-β3, explants exhibited a low basal rate of LTGF-β1 secretion (7.2 ± 2.7 ng/mL/day on average over 10 days). LTGF-β1 secretion rates increased with aTGF-β3 dose and continued to increase further over time, reaching a 7.4-fold increase at 10 ng/mL at day 10 (Fig. 4A, B). No significant degradation of secreted LTGF-β1 in the conditioned media at 37°C was observed over a 2-week period (Fig. 4C, p > 0.5). LTGF-β1 secretion increases from 1 ng/mL and 10 ng/mL aTGF-β3 doses were suppressed by LY364947 (Fig. 4D). In contrast, LY364947 had no effect on LTGF-β1 secretion increases induced by FGF-2 (Fig. 4E), supporting its use in revealing LTGF-β1 increases that result from aTGF-β activity. IGF-1, PDGF-BB, and BMP-7 exposure did not enhance LTGF-β1 secretion (Fig. 4F–H).
Upon procurement, cartilage explants possessed 51.9 ± 13.2 ng/mL of LTGF-β1 (Fig. 5A). In the absence of aTGF-β3, explants secreted 36.7 ± 4.2 ng/mL into surrounding media over 7 days (Fig. 5B). Alternatively, in the presence of 10 ng/mL aTGF-β3, explants secreted 124.5 ± 44.3 ng/mL. No significant increase of ECM-bound LTGF-β1 content of explants was observed for either group over this 7-day period (p > 0.6; Fig. 5A), indicating that newly synthesized LTGF-β1 is predominantly secreted to culture media and not retained in the tissue.
Quantification of load-induced TGF-β activity in cartilage explants
LTGF-β secretion rates measures and autoinduction standard curves were used to quantify the in situ levels of aTGF-β activity in explants in response to applied dynamic loading (Fig. 6A). It was found that 1 and 2 days after loading, LTGF-β1 secretion rates for 2 Hz were significantly elevated above free swelling (p < 0.05; Fig. 6B). LTGF-β1 secretion rates for 0.5 Hz were higher but not significantly (p > 0.3). For both loading frequencies, LY364947 significantly decreased LTGF-β1 secretion rates (p < 0.0005), approaching free swelling+LY364947 levels and confirming that load-induced LTGF-β1 secretion increases result from TGF-β signaling. Following standard curve conversion (Supplementary Fig. S2), autoinduction measures indicated in situ TGF-β activity levels of 0.22 ± 0.24 ng/mL and 0.45 ± 0.39 ng/mL for 0.5 Hz dynamic loading at days 2 and 3, respectively (Fig. 6C), whereas 2 Hz dynamic loading induced 0.55 ± 0.34 ng/mL and 0.59 ± 0.39 ng/mL in situ TGF-β activity at days 2 and 3, respectively.
Quantification of TGF-β activity in aTGF-β-conjugated constructs
LTGF-β secretion rate measures and autoinduction standard curves were used to quantify the in situ levels of aTGF-β activity in constructs conjugated with 3 µg/mL (high) and 0.3 µg/mL (low) aTGF-β3 (Fig. 7A). After 4, 6, and 8 days of culture, aTGF-β-Conj-High constructs exhibited significantly increased LTGF-β1 secretion rates relative to aTGF-β-Conj-High+ LY364947 and aTGF-β-Sol-High constructs (p < 0.0005; Fig. 7B). After 4 and 6 days, aTGF-β-Conj-Low constructs exhibited significantly increased LTGF-β1 secretion rates relative to aTGF-β-Conj-Low+LY364947 and aTGF-β-Sol-Low constructs (p < 0.005). Following standard curve conversion (Supplementary Fig. S3), autoinduction measures indicated in situ TGF-β activity levels of 1.76 ± 0.74, 1.81 ± 0.63, and 1.11 ± 0.36 ng/mL for aTGF-β-Conj-High constructs at day 4, 6, and 8, respectively (Fig. 7C). aTGF-β-Conj-Low constructs exhibited 0.07 ± 0.01, 0.05 ± 0.01, and 0.03 ± 0.01 ng/mL in situ TGF-β activity at days 4, 6, and 8, respectively.
Discussion
Quantification of in situ TGF-β activity in biological tissues has remained a long-standing challenge. In this study, we capitalize on TGF-β’s established autoinduction behavior and show that cell-secreted LTGF-β can serve as a robust, non-tissue-destructive, label-free biomarker for measuring the in situ activity of TGF-β in live cartilage tissues. The reliability of the quantification platform is supported by several key characterizations: 1) LTGF-β secretion levels are proportional to aTGF-β activity over a wide range of doses, 2) newly synthesized LTGF-β is predominantly secreted from the tissue into conditioned media, and 3) secreted LTGF-β is stable in conditioned media for durations of analysis. Overall, these characterizations support the ability to predict in situ TGF-β activity in a tissue specimen by measuring the rate of LTGF-β secretion into its conditioned media.
To determine absolute activity levels of aTGF-β in tissues, we developed a biological autoinduction standard curve that related rates of LTGF-β secretion to aTGF-β activity levels (Figs. 2B and 4B). Importantly, standard curve generation requires the ability to differentiate between supplemented exogenous aTGF-β and secreted endogenous LTGF-β in a specimen’s conditioned medium. To address this issue, we administered and analyzed different isoforms of TGF-β—exogenous aTGF-β was administered in its β3 isoform, whereas the secretion of endogenous LTGF-β was measured in its β1 isoform via an isoform-specific ELISA. Similar assessments can be performed by monitoring secretion of the β2 isoform of endogenous LTGF-β (Supplementary Fig. S4). An underlying assumption of this technique is that different isoforms give rise to similar autoinduction increases. To this end, it is important to consider that LTGF-β secretion increases in dynamically loaded cartilage explants likely result from the combined activity of endogenous β1, β2, and β3 isoforms in the tissue ECM. For bovine chondrocytes, β1 and β3 isoforms give rise to similar autoinduction responses (Supplementary Fig. S5), supporting that an aTGF-β3-based autoinduction standard curve can be used to estimate the combined isoform activity in cartilage. However, we acknowledge that different isoforms could trigger differential autoinduction responses and may need to be accounted for other tissue systems.
The autoinduction platform assumes that newly synthesized LTGF-β is predominantly secreted from the tissue (i.e., not retained in the ECM) and stable in conditioned media for several days (i.e., the duration in between media acquisition points). Although activated TGF-β is associated with a short half-life in vivo, 30 the secreted LTGF-β in conditioned media is stable for at least 2 weeks (Fig. 4C), which is in far excess of the requisite stability for analysis. In addition, we observed low levels of LTGF-β retention in both explants and tissue constructs, indicating that LTGF-β measured in conditioned media accurately reflects synthesis increases induced by aTGF-β activity. The low retention levels of synthesized LTGF-β in cartilage explants may be somewhat surprising, given that cartilage is noted for possessing high levels of LTGF-β bound to its ECM. LTGF-β incorporation into the ECM is promoted by transglutaminase-induced crosslinking between the latent TGF-β binding protein and ECM constituents, such as fibrillin. 43 The observed low retention levels suggest that the rate of LTGF-β diffusive flux out from the tissue exceeds the rate of chemical crosslinking. In addition, inhibition of transglutaminase does not lead to an increase in LTGF-β1 secretion (Supplementary Fig. S6), further supporting that only a small fraction of newly synthesized LTGF-β1 is crosslinked to the ECM.
The implementation of the autoinduction standard curve further requires confirmation that LTGF-β secretion increases result predominantly from TGF-β activity. Although LTGF-β biosynthesis and secretion is markedly enhanced by aTGF-β, it may also be influenced by other endogenous growth factors and signaling pathways,2,44 which can confound autoinduction-based measures. Here, we utilized LY364947 to block TGF-β signaling and confirm that the LTGF-β secretion increases do indeed result from in situ aTGF-β activity. As a proof of concept of this approach, we demonstrated that LY364947 leads to inhibition of LTGF-β secretion increases in response to aTGF-β signaling but no significant suppression of increases induced by an alternative FGF-2 pathway (Fig. 4E). Accordingly, LTGF-β secretion increases that are not suppressed by LY364947 can be attributed to non-TGF-β-induced signaling events.
Significant efforts have been undertaken to assess TGF-β activity in biological tissues. Although Western blotting of downstream signaling proteins (e.g., p-SMAD2/3)32–34,45,46 and qPCR of TGF-β inducible genes (e.g., Serpine1, SMAD7, ALK5) 25 have been used to assess TGF-β activity, they are destructive analyses and have not been used to quantify TGF-β activity levels. Our results further demonstrate that Western blotting on p-SMAD2 exhibits limited capability to differentiate TGF-β activity levels range in 0.1–10 ng/mL (Fig. 3). Reporter cell-based assays can quantify TGF-β bioactivity35–37 but are not compatible with intact three-dimensional tissue specimens. A significant benefit of the current autoinduction platform is its capability to perform nondestructive, quantitative measures of in situ TGF-β activity in intact tissue specimens. Yet, it is important to note that this autoinduction platform does possess some limitations. LTGF-β secretion increases exhibit a plateau above 3 ng/mL aTGF-β (Figs. 2B and 4B), thus placing a saturation limit on the range of aTGF-β activity that can be measured. Furthermore, as it relies on a biological downstream response, it would not be applicable for cells that do not express TGF-β or do not exhibit a strong autoinduction response. 47
This autoinduction measurement platform serves as an important analytical tool for TGF-β activity in musculoskeletal tissue applications. We demonstrated the capability of the platform to contribute to an important, growing area of cartilage mechanobiology: the role of mechanical loading in activating ECM-bound LTGF-β. Prior investigations have characterized the activation of LTGF-β in cartilage through qualitative assessments.25,26,46 Here, we achieved quantitative measures of the in situ load-induced activation of LTGF-β in articular cartilage using an autoinduction standard curve. We demonstrated that in the absence of loading, LY364947 induces a decrease in the LTGF-β secretion rate of cartilage explants (Fig. 6B), suggesting that chondrocytes can exhibit a low rate of constitutive LTGF-β activation. This activation may result from cell-secreted enzymes,48,49 thrombospondin-1, 50 or integrin-mediated traction forces on LTGF-β from chondrocytes.19,51–53 In response to dynamic loading, autoinduction-measured activity levels are markedly increased, reaching 0.59 ng/mL of TGF-β activity (Fig. 6C). LY364947 inhibition confirms that LTGF-β secretion increases do indeed result from load-induced TGF-β activation rather than alternative pathways that may also be activated by loading (Fig. 6B). 54 Interestingly, load-induced activity levels represent only a modest fraction of the LTGF-β content in the cartilage ECM (∼300 ng/mL).4 These results are intriguing, suggesting that even low levels of TGF-β activity may have a powerful impact on chondrocyte biosynthesis and homeostasis. Experiments were performed on a simplified cartilage tissue model of deep zone explants due to the uniformity of their composition and material properties. 55 In the future, we aim to use this platform to achieve novel characterizations of: 1) ECM-bound LTGF-β activation rates for physiological and pathophysiologic loading regimens in different zones (superficial, middle, deep) of cartilage tissues, 2) identification of the molecular mediators involved in load-induced activation cascades, and 3) the dissipation of these mechanisms with aging and/or tissue degeneration.
In the second application, we demonstrated that the autoinduction measurement platform can quantify the in situ activity of TGF-β delivered within engineered constructs. Conventional assessments of TGF-β predominantly consist of measures of TGF-β release from acellular scaffolds.21,23,32,34 However, these measures may not accurately reflect its bioactivity within constructs due to chemical processing techniques required to achieve TGF-β scaffold incorporation (e.g., scaffold conjugation or microsphere embedding) and the influence of cell/ECM reactions acting on TGF-β when cells are present. Using the autoinduction platform, we directly quantified the bioactivity of scaffold-conjugated TGF-β on embedded cells. Our results demonstrate that the in situ activity level of delivered TGF-β is <0.1% of the conjugated dose (Fig. 7C), indicating that an overwhelming majority of the growth factor may be inaccessible to cells. Interestingly, these moderate TGF-β levels can significantly enhance the mechanical properties and biochemical content of engineered cartilage (Supplementary Fig. S7), consistent with our recent report on the effects of moderated TGF-β dose administration. 15 In the future, this technique can be used to quantify in situ TGF-β activity for a multitude of scaffold delivery platforms.
Overall, TGF-β autoinduction can serve as a robust measurement platform to quantify in situ activity of TGF-β in live tissue specimens. We demonstrate its utility in two important areas of research: 1) quantifying the activity of mechanical load-activated TGF-β in native cartilage and 2) quantifying the activity of scaffold-delivered TGF-β in tissue engineering systems. Given the demonstrated versatility of this quantification platform, in the future it may be utilized for a wide range of cell types (e.g., chondrocytes, fibroblasts, stem cells) and tissue sources (e.g., cartilage, meniscus, tendons, bone, intervertebral disk), and for a variety of applications, such as tissue culture model and regenerative medicine platforms. In a broader context, this work advances the potential for adopting cell-secreted protein monitoring as a novel technique for quantifying specific in situ cellular processes, which may remain difficult to measure. As such, the identification of novel protein biomarkers that reflect specific cellular biochemical processes and exhibit minimal retention in the tissue ECM may allow for useful, easy-to-implement, nondestructive platforms for advancing insights into an assortment of tissue behaviors.
Footnotes
Acknowledgments
The authors thank Dr. Enkhjargal Budbazar for his assistance with Western blot analysis.
Authors’ Contributions
T.W., S.Y.K., and M.B.A.: Conceived the idea and designed the experiments. T.W., S.Y.K., Y.P., and J.Z.: Performed the experiments and analyzed the data. T.W., S.Y.K., and M.B.A.: Interpreted the data and wrote the article. M.D.L. and J.E.M.: Contributed to the scientific discussions, data interpretation, and the article.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Disclosure Statement
All authors declare no competing financial interests exist.
Funding Information
Research reported in this publication was supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases under award number R01AR078299, the National Science Foundation under award number CMMI 1906469, the Boston University Micro and Nano Imaging Facility and the National Institutes of Health under award number S10OD024993, and the Boston University Distinguished Summer Research Fellowship.
Disclaimer
The opinions, findings, conclusions, or recommendations expressed are those of the authors and do not necessarily reflect the views of the National Institutes of Health or the National Science Foundation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.