
Abstract
Background:
Resveratrol, a polyphenol found in grapes, exhibits several beneficial health effects by its antioxidant, antiinflammatory, and chemopreventive properties. The aim of the present study was to determine the effect of resveratrol on iodide trapping and efflux as well as its mode of action using FRTL-5 cells, having in mind the pivotal role of the natrium iodide symporter (NIS) in the treatment of differentiated thyroid cancers.
Methods:
Cells were treated with resveratrol for various times and doses, in the presence or absence of thyrotropin (TSH). Iodide trapping, iodide efflux, rat NIS (rNIS) protein expression, and cyclic AMP (cAMP) production were evaluated.
Results:
Resveratrol increased iodide trapping in a time-dependent (optimal 6 hours) and dose-dependent (100 μM) way in the presence of TSH. It showed an additive effect when concomitantly added with an optimal dose of TSH. Resveratrol (50 μM) increased (threefold) rNIS protein expression. In TSH-deprived cells, resveratrol also provoked an increase in rNIS protein (>3-fold in 6 hours) with an optimum at 40 μM. Resveratrol did not inhibit iodide efflux from FRTL-5 cells. It neither increased intracellular cAMP nor induced the arborization of living cells, two TSH-induced effects. A non-cAMP mode of action is highly suspected.
Conclusions:
Resveratrol increases iodide trapping in FRTL-5 cells, increasing iodide influx and rNIS protein level even in the absence of TSH. It has an additive effect with TSH. Consequently, resveratrol could be a promising molecule for radioiodide therapy in follicular and papillary differentiated thyroid carcinoma in association with recombinant human TSH.
Introduction
In this paper we demonstrate that resveratrol activates iodide trapping in the presence of TSH, the natural stimulator of the thyroid gland. Resveratrol increases rat NIS (rNIS) protein expression in the presence or absence of TSH. Its action does not result in an inhibition of iodide efflux. The mechanism of action of resveratrol through adenosine receptors (19), membrane lipids as ceramide (20), nitric monoxide (NO) (21), or cyclic AMP (cAMP) (22) is still controversial. We probed the cAMP pathway and data are clearly in favor of a cAMP-independent effect of resveratrol.
Materials and Methods
Materials
Resveratrol, insulin, transferrin, hydrocortisone, bovine thyrotropin (bTSH), MTT formazan (1-(4,5-dimethythiazol-2-yl)-3,5-diphenyl formazan), and Coon's modified Ham F12 medium were purchased from Sigma (St. Louis, MO). Glycyl-L-histidyl-L-lysine acetate and somatostatin were from Calbiochem-Behring (La Jolla, CA). All reagents were of analytical grade. Purchased from Falcon (Becton-Dickinson, Meylan, France) were 75 cm2 flasks and 24-cluster 2 cm2 plastic tissue culture wells. The cAMP kit was from Beckman Coulter (Marseille, France). Polyclonal antibodies against rNIS were a gift from Prof. N. Carrasco (Albert Einstein College of Medicine, New York). Peroxidase-conjugated anti-rabbit IgG, horseradish peroxidase (HRP) linked, used as secondary antibodies, was from Cell Signaling Technology (Saint-Quentin-en-Yvelines, France).
Cell culture
Fisher Rat Thyroid cells (FRTL-5) kindly donated by Dr. A. Virion (INSERM U 486, Châtenay-Malabry, France) were routinely grown in Coon's modified Ham F12 medium supplemented with 5% fetal calf serum and the six components (6H), that is, 1 mU/mL TSH, 5 μg/mL insulin, 5 μg/mL transferrin, 10 nM hydrocortisone, 10 ng/mL somatostatin, and 10 ng/mL glycyl-L-histidyl-L-lysine acetate (standard medium) (18). Cells were cultured in 24-well clusters or 75 cm2 flasks at 37°C in an atmosphere of 95% air and 5% CO2 in a water-saturated incubator. Where indicated, cells were cultured 7 days in 5H medium (i.e., 6H standard medium minus TSH). In this TSH-free conditions, media were changed every 48 hours (4 medium changes).
MTT assay
To estimate the number of living cells per well, we used the colorimetric MTT assay (23). Briefly, 50 μL of a 5 mg/mL MTT solution was catabolized for 60 minutes by mitochondrial dehydrogenase into formazan blue. Formazan blue was extracted in 450 μL isopropanol–HCl 0.04 N, and the absorbance was measured using a test wavelength of 570 nm and compared against a reference wavelength of 630 nm. Cell number was also estimated by counting under a Thoma hemocytometer, and the linear relationship between cell number and absorbance obtained from the MTT test was determined. This allowed the quantification expression of iodide trapping per living cell.
Measurement of 125I trapping
To each cell culture well containing 450 μL medium and showing about 90% confluency, 50 μL of reducing-agent-free 125I in 10−5 M potassium iodide (KI) was added to get a final radioactivity of about 10 kBq and a concentration in iodide of 10−6 M KI, a value close to the apparent Michaelis constant (Km) of NIS. Specific iodide trapping by cells plateaued at equilibrium between influx and efflux in less than 30 minutes (24,25). Specific iodide trapping is the difference between total counts measured in cell layers after 30 minutes of incubation in the absence of sodium perchlorate (NaClO4) and nonspecific counts measured in cell layers of similar wells incubated 30 minutes in the presence of 1 mM sodium perchlorate, as a competitive inhibitor of iodide uptake (18,25). Nonspecific counts are the sum of iodide passive diffusion, membrane-lipid-linked iodide, iodide adsorption on plastic wells' lateral surfaces, and nonspecific cell surface adsorption. Usually, 2 × 4 wells (±NaClO4) were used to determine one experimental point. In all cases, media were withdrawn after 30 minutes of incubation at 37°C (18). Cell layer extracts were obtained by sequential addition of 2 × 500 μL 1N NaOH. All results mentioned in figures represent intracellular-specific counts. Extracts were counted in a W + W electronic MR 101 gamma counter.
Measurement of iodide efflux
Iodide efflux was measured as described by Fayet and Hovsépian (24). Briefly, FRTL-5 cells were maintained in their regular 6H medium at 95% confluency, treated with 20 μM resveratrol or carrier solution only during 6 hours, and labeled with radioactive 125I− in the presence or absence of 1 mM NaClO4. After 30 minutes (at equilibrium) media were removed and replaced by prewarmed (37°C) identical media without radioactivity. After 0, 5, 10, 15, and 20 minutes at 37°C, media were discarded and intracellular radioactivity was determined. Specific counts for each point were the difference between total counts (without ClO4−) and nonspecific counts obtained with perchlorate. Consequently, we had four different situations in these efflux assays and four identical wells for each experimental point, and these experiments were performed in triplicate: Efflux with carrier solution (total counts) Efflux with carrier solution and NaClO4. NaClO4 was added at equilibrium, that is, at 30 min. Efflux with resveratrol (total counts) Efflux with resveratrol and NaClO4. NaClO4 was added at equilibrium at 30 min.
Western blot analysis
FRTL-5 cells were cultured in their standard medium containing 1 mU/mL TSH. When cell surface saturation of the 75 cm2 plastic flasks reached 95–100%, cells were challenged with various doses of resveratrol for 6 hours, in a refreshing medium. Identical amounts of cells were resuspended in 5 vol of homogenizing buffer (50 mM Tris–HCl [pH 7.5], 150 mM NaCl, 5 mM EDTA, 1% Triton × 100, and 0.1% sodium dodecyl sulfate [SDS]). After gentle homogenization at 4°C, lysates were centrifuged at 700 g for 10 minutes at 4°C. Supernatants were stored at −20°C after protein estimation using the micro bicinchoninic acid (BCA) protein assay reagent kit (Sigma, St. Louis, MO). Aliquots (100 μL) were mixed with the same volume of sample buffer (10% glycerol and 2% SDS in 0.0625 M Tris-HCl pH 6.8) and then loaded on 7.5% SDS polyacrylamide gel electrophoresis according to Sbarra et al. (6). Separated proteins were electrotransferred to nitrocellulose membranes overnight. Ponceau-Red coloration of blots was used to verify the protein loading, and blots were further immunostained according to Hawkes et al. (26). Briefly, after blocking with 3% BSA buffer, blots were exposed to anti-rNIS polyclonal antibodies (1:1000) and mouse antiactin (1:2000, cross-reacting with rat actin) at room temperature for 2 hours. Peroxidase-conjugated anti-rabbit IgG was then added (1:3000) for 1 hour at room temperature. Detection was made with the chemiluminescent peroxidase (POD)-substrate lumic-light Western Blotting substrate (ROCHE) reagent and spots quantified by densitometry (NIH image program at
Radioimmunoassay of cAMP
Cells were cultured 1 week in TSH-free 5H medium (four media changes), then incubated with resveratrol (20 μM), TSH (1 mU/mL), resveratrol + TSH, or carrier solution medium in the presence or absence of 50 μM theophylline, a phosphodiesterase inhibitor. cAMP was estimated in media (27) after 0, 15, 60, 180, and 360 minutes using a radioimmunoassay kit according to the manufacturer's protocol.
Protein determination
Protein concentration was determined by the micro bicinchoninic acid (BCA) protein assay kit (Sigma) using bovine serum albumin as standard (28).
Statistical analysis
Data were analyzed by unpaired Student's t-test or one-way analysis of variance and are expressed as means ± standard error of the mean. Data are representative of six independent experiments and assays have been carried out in triplicate. Statistical analyses were conducted using GraphPad InStat version 3.0a for MacIntosh (GraphPad Software, San Diego, CA). All statistical tests were two-tailed, and a p-value of 0.05 or less was considered statistically significant.
Results
Effect of resveratrol on FRTL-5 cell survival
FRTL-5 survival maintained in their standard medium supplemented with various doses of resveratrol (0–100 μM) was assessed by MTT test after 6 hours of treatment (Fig. 1). A slight toxicity reaching 10–12% at 20 μM and 23% at 100 μM resveratrol was observed. Consequently, all data of iodide trapping were expressed per cell number, keeping in mind the linear correlation between cell number and MTT assay (data not shown).
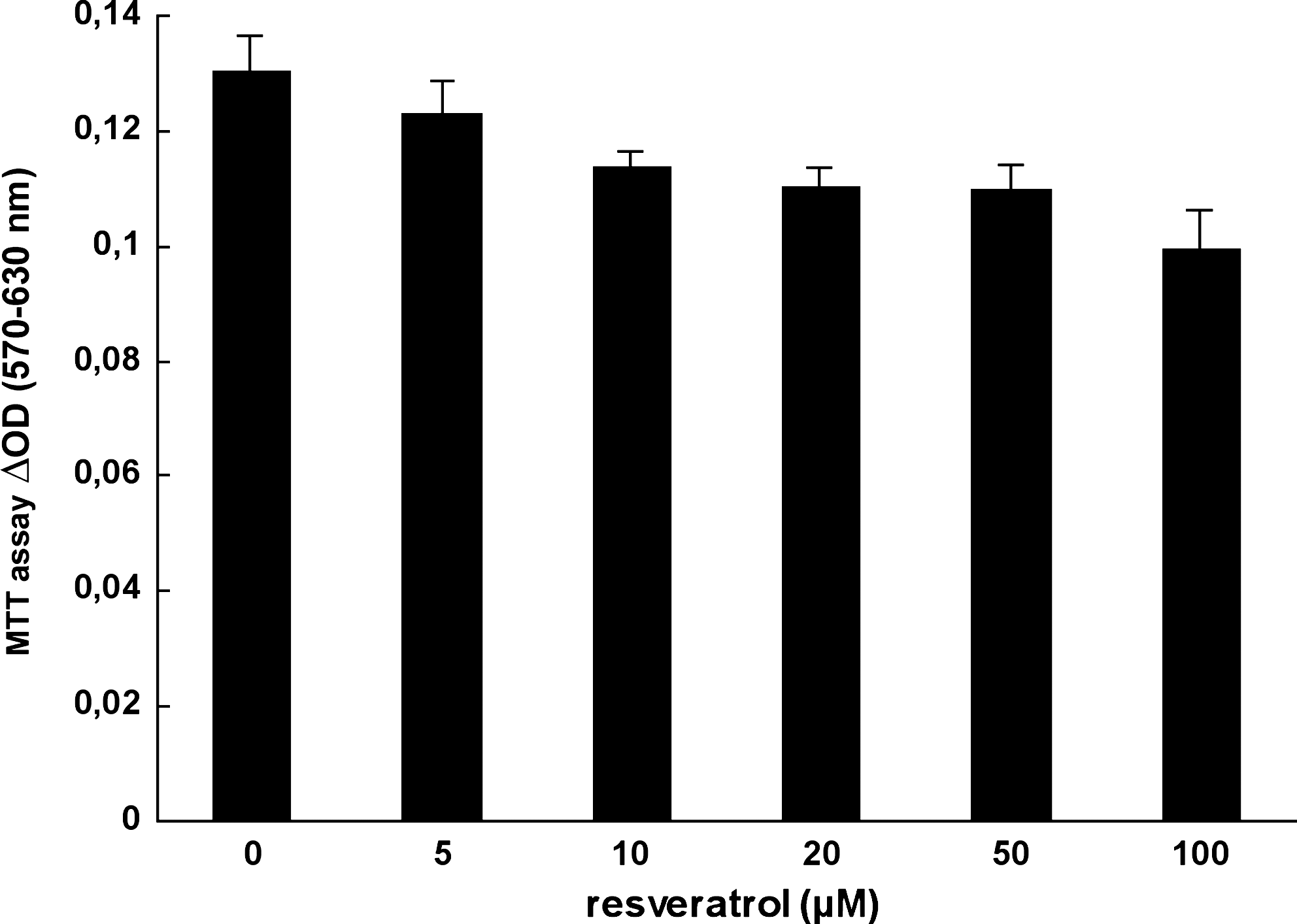
Resveratrol effect on FRTL-5 survival. FRTL-5 cells were grown in F12 Coon's modified medium enhanced with 5% fetal calf serum and 6H. They were treated with increasing doses of resveratrol, and 6 hours later, the cell survival was determined using the MTT formazan assay.
Effect of resveratrol on iodide trapping
Time course study
FRTL-5 cells were cultured with or without 20 μM resveratrol in their standard 6H (TSH-containing) medium for the indicated times (Fig. 2), and iodide trapping was measured at equilibrium. Specific intracellular counts represent 84–88% of total counts. Results indicated that the level of iodide trapping expressed per cell increased from 25% to 30% between 6 and 12 hours. However, after 24 hours of incubation the stimulation decreased and was not significant. Consequently, resveratrol increased intracellular iodide levels in FRTL-5 cells even in the permanent presence of 1 mU/mL TSH, suggesting an additive effect with TSH.
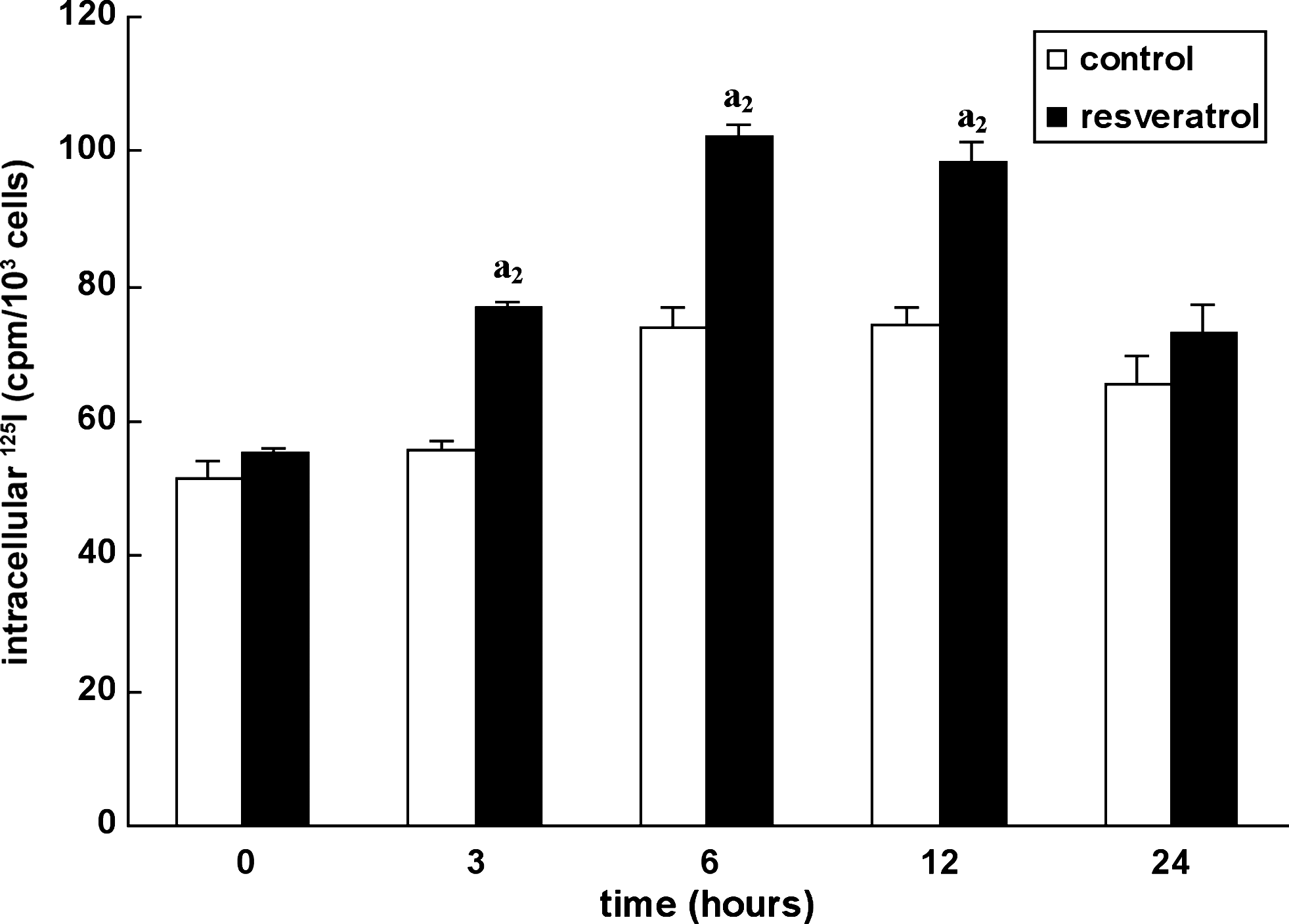
Resveratrol effect on iodide trapping: time course study. Iodide trapping was measured at equilibrium between influx and efflux (30 minutes of incubation) in FRTL-5 cells cultured in 6H standard medium or after addition of 20 μM resveratrol at 0, 3, 6, 12, and 24 hours. Results are expressed as specific intracellular 125I counts per minute (cpm) per 103 cells.
Dose-response study
FRTL-5 cells maintained in their standard 6H medium were challenged with various concentrations of resveratrol ranging from 0 to 100 μM during 6 hours. Iodide trapping measurements indicated that resveratrol had induced a clear increase in iodide uptake reaching 66% at 100 μM (Fig. 3). Specific intracellular counts represent 81–88% of total counts. Once again these data suggested an additive effect between TSH and resveratrol.
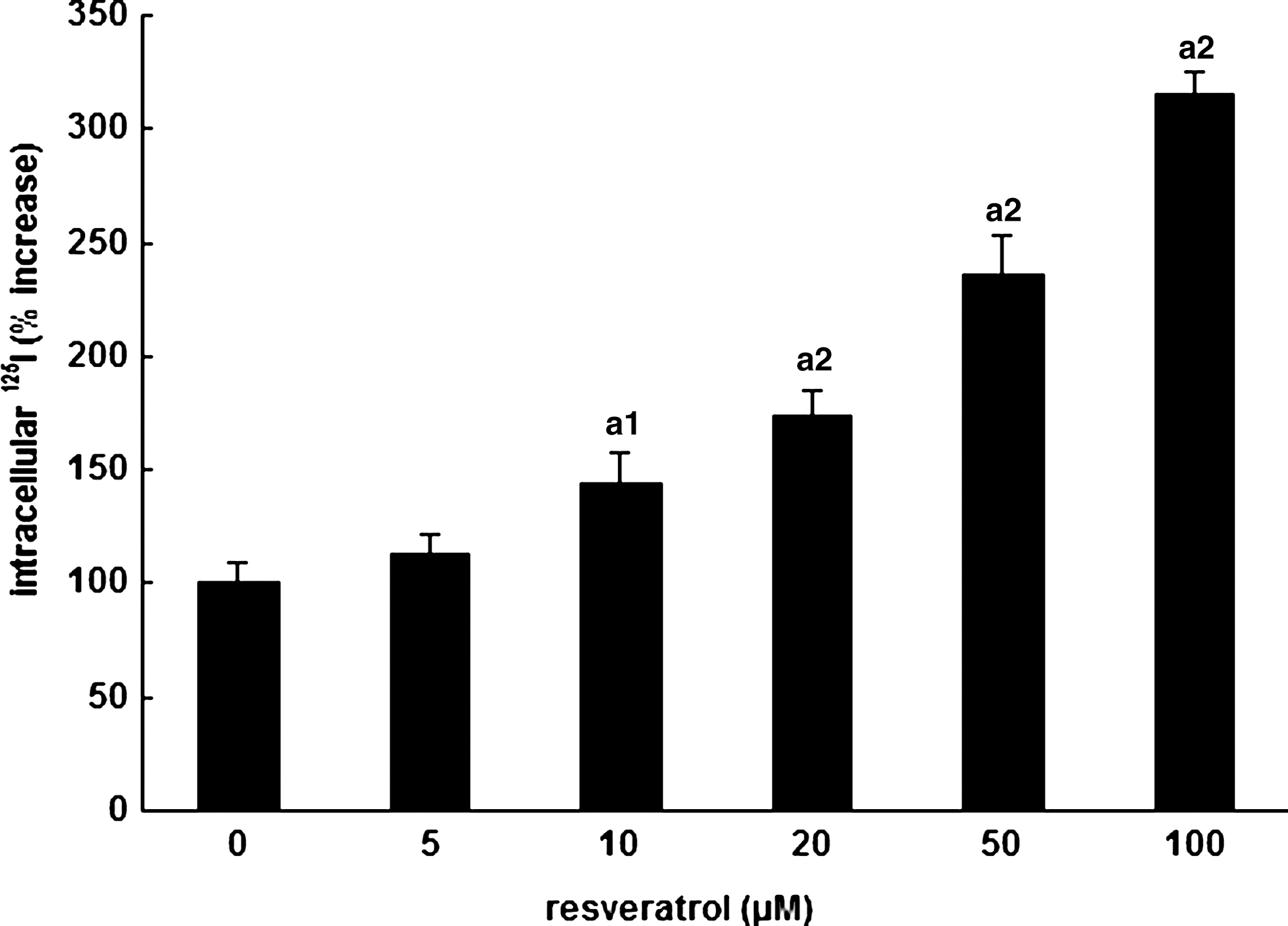
Resveratrol effect on iodide trapping: dose–response study. Iodide trapping expressed as the percent increase versus control of specific intracellular 125I cpm per 103 cells was measured 6 hours after addition of increasing doses of resveratrol. 0 μM represents the carrier solution medium.
Additive effects of resveratrol and TSH on iodide trapping
FRTL-5 cells were cultured for 1 week in 5H medium. At the onset of the experiment, resveratrol was added at 0 (carrier solution only), 20, or 50 μM alone or together with 1 mU/mL TSH. Intracellular iodide levels were measured at equilibrium after 12 hours of incubation. In all cases, the association TSH + resveratrol induced an increase in iodide trapping when compared with resveratrol alone (Fig. 4). Specific intracellular counts represent 69–72% of total counts. The effects of the two molecules are obviously additive.

Additive effect between TSH and resveratrol on iodide trapping. Cells were cultured in the absence of TSH for 1 week and challenged with 0, 20, or 50 μM resveratrol in the presence (+) or absence (−) of 1 mU/mL TSH during 12 hours. Both TSH and resveratrol increased iodide trapping and when added concomitantly showed additive effects.
Effect of resveratrol on iodide efflux
Data shown in Figure 5A first confirmed the expected higher iodide trapping in resveratrol-treated cells versus control at equilibrium (time 0 of efflux). When total trapping at equilibrium is set at 100% and iodide efflux expressed as residual percentage of total trapping versus time, values were always higher in the presence of resveratrol versus control (Fig. 5B). This indicates that resveratrol treatment either inhibited iodide efflux from FRTL-5 cells or activated influx during these efflux experiments. When NaClO4 was added at equilibrium (time 0 of efflux), the residual intracellular 125I was not different in control versus resveratrol-treated cells (Fig. 5C). This indicates that resveratrol does not block iodide efflux. In these experiments, specific intracellular counts represent 95% of total counts.

Effect of resveratrol on iodide efflux: FRTL-5 cells were exposed to medium without or with 20 μM resveratrol during 6 hours, and radiolabeled with 125I during 30 minutes (at equilibrium). Residual-specific intracellular 125I iodide was measured 0, 5, 10, 15, and 20 minutes after equilibrium (
Effect of resveratrol on NIS protein expression
We then wondered whether the physiological effect of resveratrol on iodide accumulation was dependent on rNIS protein expression. FRTL-5 cells were cultured during 1 week in their standard medium containing 1 mU/mL TSH, and treated with various doses of resveratrol for 6 hours. Postnuclear (700 g) fractions were then isolated, and rNIS protein expression was analyzed by Western blotting. Antibodies to rNIS detected a unique protein band with a molecular weight of 80–82 kDa that correlates with that of the rNIS (29). Data shown in Figure 6 unequivocally demonstrated an increase in rNIS protein expression reaching three times the control level in between 20 and 50 μM resveratrol. Densitometric values of rNIS levels versus resveratrol are also represented in Figure 6.
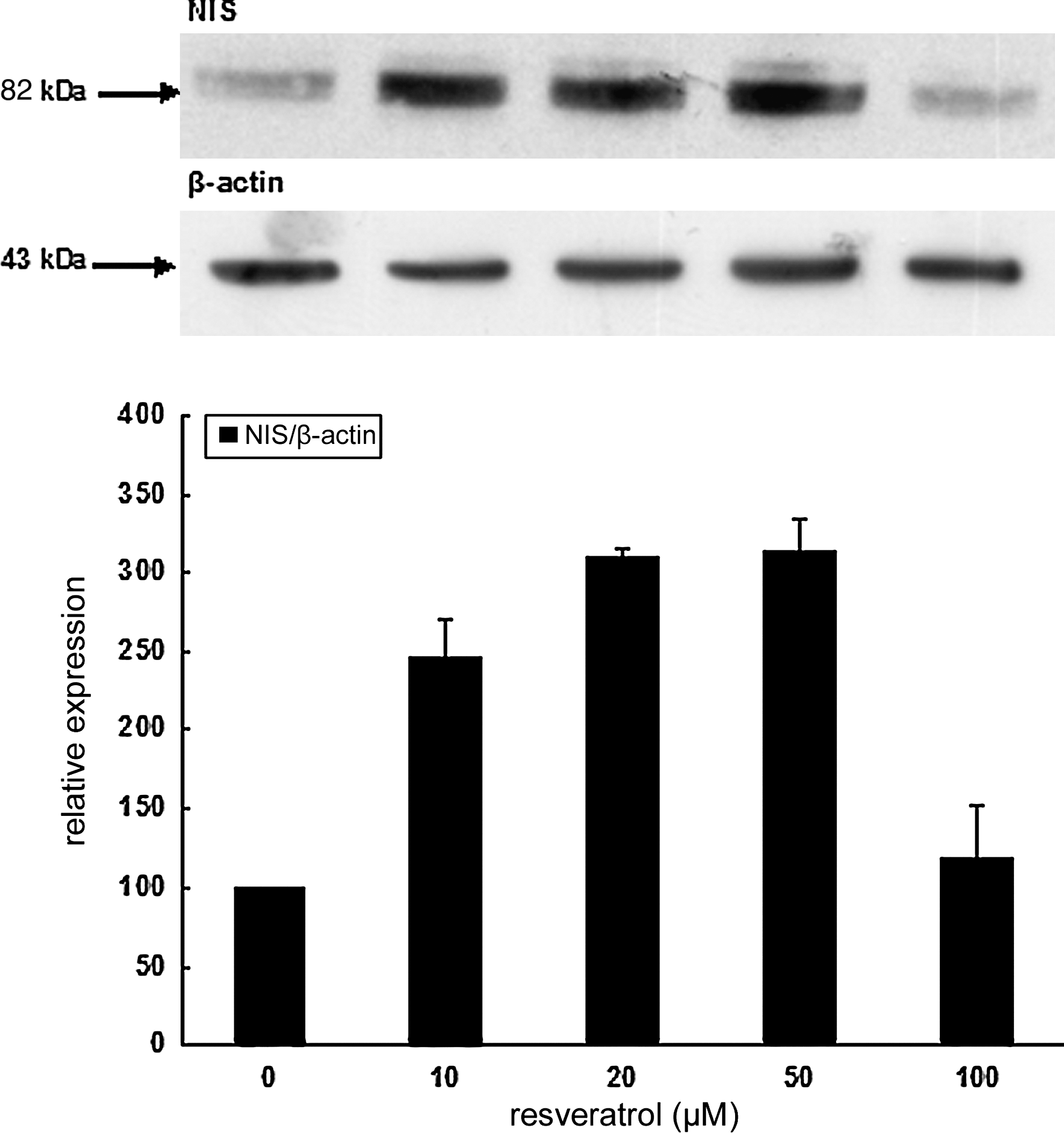
Effect of resveratrol on rat natrium iodide symporter (rNIS) protein expression: Western blot analysis. FRTL-5 cells were cultured 1 week in their regular medium containing 1 mU/mL TSH and challenged with various doses of resveratrol for 6 hours. After cell lysis, proteins from postnuclear supernatants (700 g) were collected. Proteins (20 μg per lane) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis and electrotransferred to nitrocellulose membrane. Protein transfer and loading were checked by Ponceau-Red coloration and replicates were further incubated with antibodies to rNIS. Data are representative of a typical experiment of three independent ones. Densitometric values (mean ± standard error of the mean, n = 3) of rNIS levels versus resveratrol expressed in arbitrary units corrected for β-actin as loading control.
Similar Western blots of the rNIS protein levels were also performed in TSH-free media, that is, in 5H media with increasing doses of resveratol. Results unequivocally show an increase in rNIS protein levels with doses of resveratrol as indicated in Figure 7. A maximal value reaching more than three times the control at 40 μM was found, a value in agreement with the threefold increase obtained with 20–50 μM resveratrol in the presence of TSH (Fig. 6).
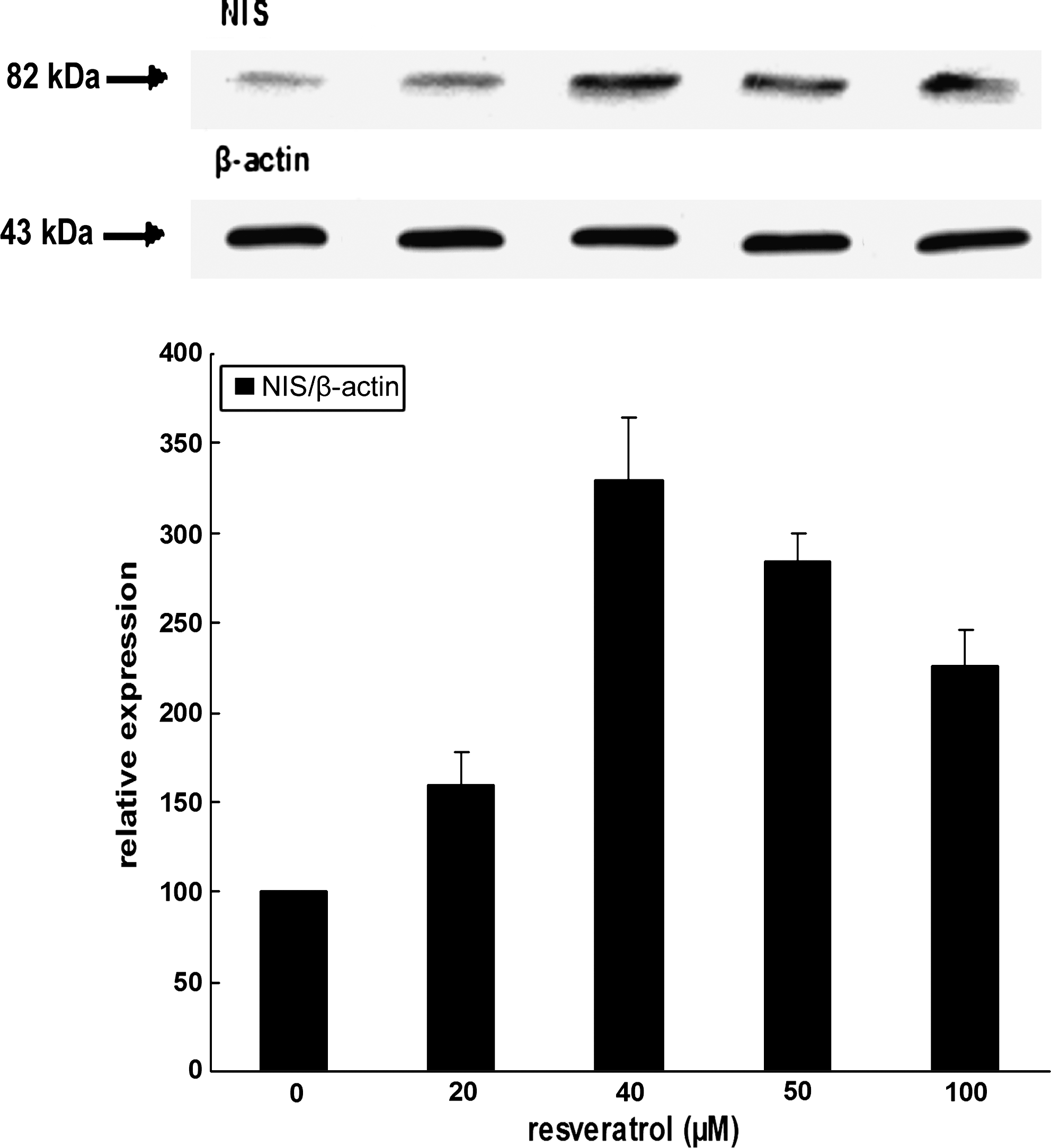
Effect of resveratrol on rNIS protein expression: Western blot analysis. FRTL-5 cells were cultured 1 week in the absence of TSH (5H medium) and challenged with increasing doses of resveratrol for 6 hours. After cell lysis, proteins from postnuclear fractions (700 g supernatants) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis, electrotransferred, and incubated with rNIS antibodies and quantitated as previously (see Fig. 6).
Effect of resveratrol on cAMP production
Figure 8 shows the effects of resveratrol (20 μM), TSH (1 mU/mL), or both molecules on cAMP production in FRTL-5 cells maintained during 1 week in their standard medium without TSH. At each time point and for each experimental condition, extracellular cAMP was measured because it has been previously shown that most cAMP is extruded into the extracellular milieu (27). Only data of cAMP production in the absence of theophylline are shown. TSH alone highly increased cAMP production versus control by 800% after 1 hour and 1375% after 3 hours. Resveratrol alone had no significant effect and slightly decreased cAMP production by 41% at 1 hour, 24% at 3 hours, and 14% at 6 hours when concomitantly added with TSH. These data strongly suggested that resveratrol does not act through the cAMP pathway.
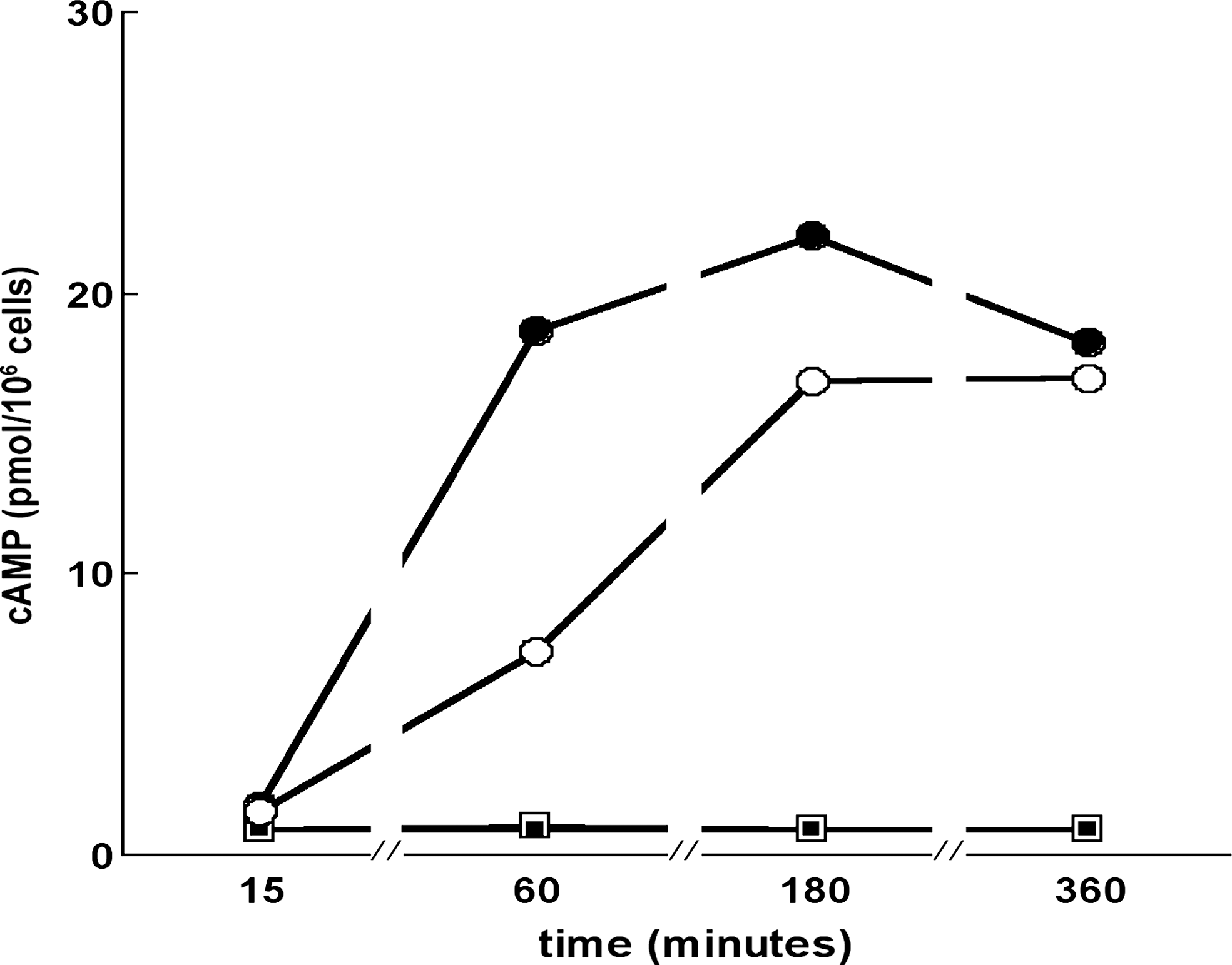
Effect of resveratrol on cyclic AMP production. FRTL-5 cells were cultured 1 week in TSH-depleted media, treated with either 1 mU/mL TSH or 20 μM resveratrol, or both, and cyclic AMP was measured in cell culture media. TSH alone (solid circles); control carrier solution (open squares); resveratrol alone (solid squares); resveratrol + TSH (open circles) compared to TSH alone. Each point is the average of two independent, very close experimental values.
Effect of resveratrol on FRTL-5 living cell morphology
When TSH (1 mU/mL) was added to the media of 5H cells, that is, in TSH-free conditions (see Materials and Methods), cells underwent a dramatic change in morphology, starting 30 minutes after TSH addition. Six hours after TSH addition, all cells were stellate shaped, membranes displaying arborizations as previously mentioned for dog thyroid cells (30). Such observation (Fig. 9A), which is common, is in total agreement with the literature considering that such TSH action is mimicked by cAMP not only in dog but also in human, porcine, and ovine thyroid cells. When resveratrol (0–100 μM) was added, such morphological modifications had appeared neither in the following 30 minutes nor in 6 hours (Fig. 9B), strengthening the hypothesis of a non-cAMP mode of action for resveratrol.
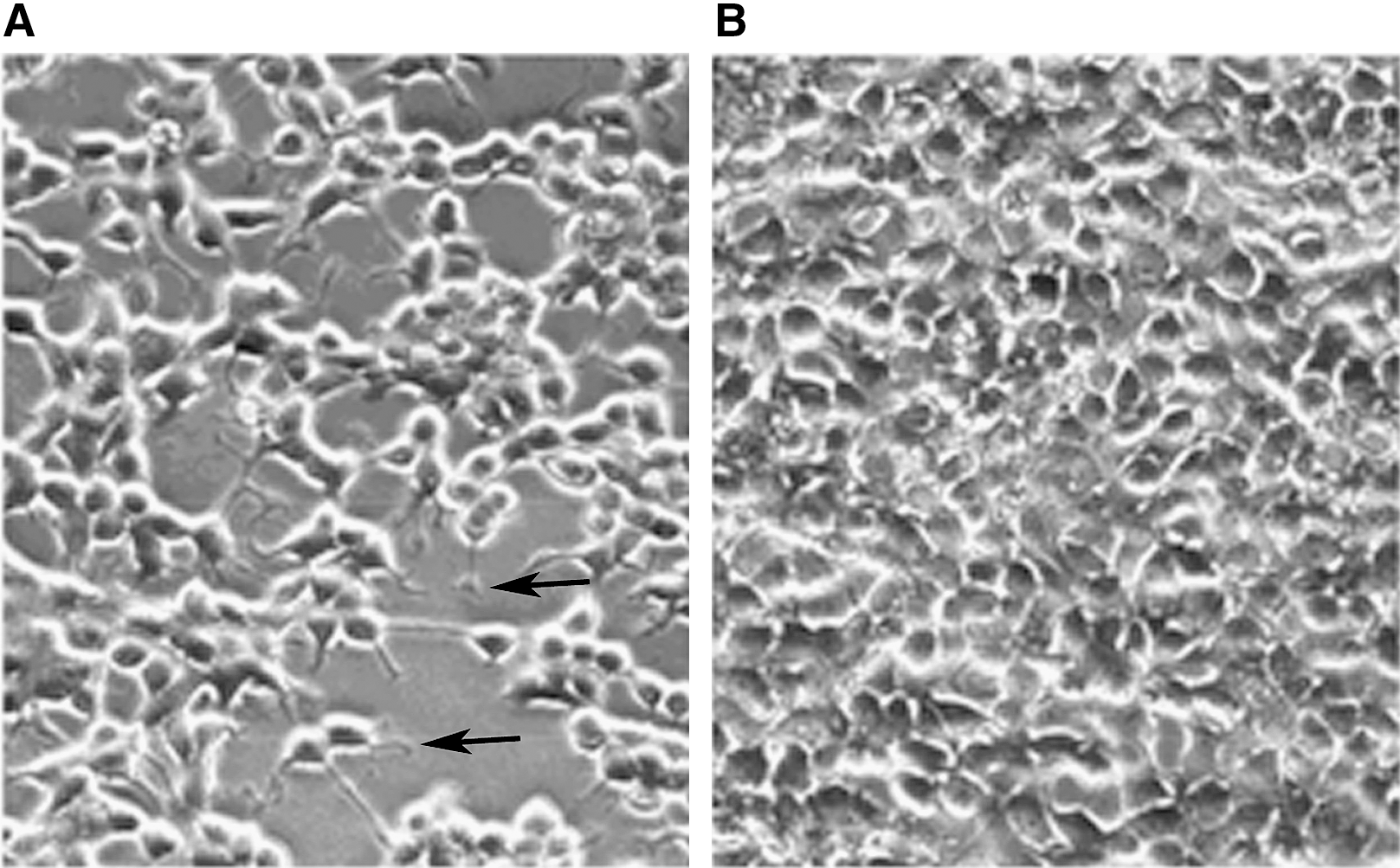
TSH-induced expression of cytoplasmic arborization of thyroid cells cultured in 5H TSH-free media. Arborization is obtained in 30 min (not shown) and still present 6 hours later as shown in (
Discussion
In this article we demonstrate that resveratrol, previously described as a hypolipidemic, antioxidant, antimutagenic, antiinflammatory, and chemopreventive agent, also increases iodide trapping in the rat thyroid cell line FRTL-5. Resveratrol increased iodide trapping in a dose-dependent manner evenin the presence of optimal concentrations of TSH. Our experiments performed with the combination of the two molecules suggested an additive rather than a synergistic effect. This efficiency obtained with a low dose of the polyphenol (20 μM), which provoked only a slight growth inhibition (10–12%), suggests a relevant pharmacological effect.
Thyroid cancer is the most common endocrine malignancy. Patients with anaplastic thyroid carcinoma have a mean survival of less than 1 year after diagnosis, despite surgery, chemotherapy, and/or radiotherapy. Well-differentiated thyroid carcinomas have a more favorable prognosis. Remnant thyrocytes that escape surgery are eliminated after radiolysis by 131I iodide active trapping. This property is, in part, dependent on the sodium iodide symporter NIS (16), also referred to as SLC5A5, which transports iodide and sodium from the extracellular milieu to the intracellular space. Unfortunately, in long-term follow-up of differentiated thyroid carcinoma, iodide trapping sometimes decreases, and thus radioactive iodide treatment becomes inefficient with time in a subset of patients. Consequently, any molecule able to positively regulate iodide transport is of great interest both for basic and clinical thyroidology.
We demonstrated that resveratrol-induced iodide trapping is at least partly mediated by an increase in rNIS protein expression. This was also the case for adenosine-induced iodide trapping with some differences however. For instance, Harii et al. (31) described a discrepancy between adenosine-induced iodide uptake activity (eightfold) and NIS protein levels (threefold). In our study, a closer correlation between resveratrol-induced iodide uptake (3.5-fold, Fig. 3) and rNIS protein level (threefold, Fig. 6) was observed. Adenosine was shown to increase iodide accumulation by acting both on NIS mRNA and protein expression (31). Such effects of adenosine, which are mimicked by N6-(L-2-phenylisopropyl) adenosine and antagonized by 1,3-dipropyl-8-cyclopentylxanthine, suggest that adenosine acts through the adenosine A1 receptor. In the study by Harii et al. (31), the authors studied the effects of adenosine on iodide transport in the absence of TSH, or compared adenosine-induced effects to TSH alone. However, they did not report an additive or synergistic effect between the two compounds, suggesting that adenosine and TSH impact on the same signaling pathway despite differences in plasma membrane receptors. To explain effects of resveratrol on rat heart, Das et al. (32) suggested a role of cAMP response element binding (CREB)-dependent Bcl-2 signaling via A3 receptor activation.
Another original new result given in this article concerns the effect of resveratrol on TSH-deprived cells. It increases NIS in a dose-dependent manner even in the absence of TSH (Fig. 7). This fits with the additive effect of resveratrol and TSH on the physiological transport of iodide and reinforces the concept of a distinct mode of action. TSH works through the cAMP pathway, whereas resveratrol certainly acts through a different, still unknown molecular pathway, following different kinetics as indicated in the next paragraphs.
According to the potential interest for using resveratrol in therapeutic strategies aiming at NIS-mediated 131I therapy in thyroid cancers (17), which is dependent on time of contact with the isotope, we tried to perceive whether resveratrol could limit iodide efflux. Our data show that resveratrol (Fig. 5C), on the whole, does not inhibit iodide efflux. It is noteworthy that adenosine had no effect on iodide efflux (31), whereas decitabine and 5-azacytidine were only tested in terms of effects on iodide trapping and NIS mRNA expression (33).
The antioxidant properties of resveratrol are well known (34), and are shared with methimazole (1-methyl-2 mercaptoimidazole (MMI) or its precursor carbimazole), an inhibitor of thyroperoxidase. In our hands, MMI at 1 mM, a level known to completely block thyroperoxidase, did not enhance the level of iodide trapping as expected from previous studies obtained with primary cultures of porcine thyroid cells (24). However, MMI increased thyroglobulin mRNA level (35,36). Resveratrol and MMI are double bond-rich molecules. They are free radical scavengers, a property linked to their antioxidant action. The thyrocyte is a cell producing high amounts of H202. The interaction of resveratrol with components of the hydrogen-peroxide-generating system needs to be analyzed. It would be rational also to test the effects of resveratrol on thyroglobulin levels. This has not yet been performed as far as we know.
We also show (Fig. 8) that resveratrol does not increase iodide trapping through the cAMP pathway. In this respect, MMI (35,36), hexamethylene bisacetamide (37), and adenosine (31) have been shown not to act via cAMP. At the morphological level, in living cells, resveratrol does not provoke the well-known arborization of FRTL-5 cells observed with TSH in 5H cultures. This morphological effect is cAMP mediated (30). Resveratrol has been shown to increase only cAMP levels in both time- and concentration-dependent manners in a few instances and acted as an agonist for the cAMP/kinase A system in breast cancer cells (38). Resveratrol was also shown to stimulate cAMP-dependent chloride secretion in intestinal cells in vitro (39). Our results are rather in accordance with those demonstrating a cAMP-independent effect of resveratrol on prostaglandin-induced platelet aggregation (40), on progesterone secretion by mouse tumor Leydig cells (41), in coronary smooth muscle (42), and in lung epithelial cells (43).
The mode of action of resveratrol is still unclear. Although we do not know whether the observed effects of resveratrol on rNIS are transcriptional, translational, or posttranslational, its relatively fast effects are in favor of a postranscriptional mode of action.
An increased iodine uptake in thyroid carcinoma after treatment with sodium butyrate and decitabine (azacytidine), which are respectively histone deacetylase and histone methylation inhibitors, has recently been described (33). Our present results obtained with resveratrol may appear as contradictory because resveratrol is a well-known histone deacetylase activator (44), a characteristic demonstrated in Saccharomyces cerevisae. This eukaryotic cell is, however, clearly distinct from thyroid cells.
In some cases resveratrol seemed to act via NO as in rat hearts after ischemia-reperfusion injury (45), in cyclosporine-induced nephrotoxicity in rats (46), in cultured rat cardiac fibroblasts (47), or after focal cerebral ischemia injury (48). More recently some of us demonstrated strong cardioprotective effect of resveratrol in rat hearts after ischemia-reperfusion injury that was not mediated by NO (49). On the other hand, resveratrol was shown to act independently from NO by abrogating lipopolysaccharide-induced NO production in microglia (50) or in macrophages (51). Resveratrol was also shown to protect myocardial ischemia-reperfusion injury through both NO-dependent and NO-independent mechanisms by simultaneously inducing endothelial NO synthase and repressing inducible NOS (52). The implication of NO in the mode of action of resveratrol is still unresolved, including in thyroid cells, as we do not yet know if resveratrol acts either intracellularly or via receptors on the cell surface membrane. Interestingly, resveratrol has been recently shown to induce apoptosis via signaling through plasma membrane lipid rafts involving the death receptor pathway (53), or via ceramide (54 –56). Such receptors could be compared to high-affinity plasma membrane resveratrol binding sites as described in rat brain (57). The putative NO or ceramide implication in the mode of action of resveratrol is still a subject of debate.
In conclusion, we have shown that resveratrol simultaneously stimulates iodide trapping by increasing iodide influx and rNIS protein expression. It does not inhibit iodide efflux in FRTL-5 cells. These effects of resveratrol should be tested on differentiated human thyroid cancer cells and in normal human thyroid cells in primary culture. According to its additive effects with TSH, resveratrol may be a promising molecule for the treatment of thyroid differentiated carcinoma, considering it could work as an adjuvant to recombinant human TSH.
Footnotes
Acknowledgments
The authors would like to thank Prof. Nancy Carrasco and Dr. Cécile Basquin for the kind gift of polyclonal antibodies directed against rNIS and fruitful discussion.
Disclosure Statement
The authors declare no competing interests.