
Abstract
Background:
A member of the genes encoding WD-repeat proteins, the WDR3 gene, maps in the 1p12 region. This region was shown to be associated with thyroid cancer susceptibility in a previous work. In this study we aim to evaluate the contribution of WDR3 to thyroid cancer risk.
Methods:
A case–control association study was performed in a total of 402 patients and 479 control subjects from a Spanish population. In the initial phase of the study, 10 single-nucleotide polymorphisms covering the WDR3 region were genotyped in a small group (157 patients and 118 control subjects); next, three of the initial single-nucleotide polymorphisms were further genotyped in the overall population. In addition, WDR3 expression was investigated in 10 thyroid cancer cell lines by RT-PCR and Western blot.
Results:
Haplotype analysis revealed that combination of certain WDR3 variants, such as haplotype CAT, increases the risk of thyroid cancer (odds ratio = 1.85, 95% confidence interval = 0.97–3.55, p = 0.063). Further, both messenger RNA transcription and protein expression of WDR3 were altered in human thyroid cancer cells.
Conclusion:
These results indicate for the first time that WDR3 is a risk factor to thyroid cancer, suggesting its implication in the etiology of thyroid cancer.
Introduction
The large group of proteins encoded for the WD-repeat gene family have a related structure, although with diverse functions (10 –12). These proteins are involved in cell cycle progression and signal transduction, among other cellular processes. Yeast-based research have shown that the human WDR3 orthologous encodes a DOM-interacting protein, which is involved in G1 cell progression and cell differentiation (13); subsequently, a role of the WDR3 gene in cell division has been proposed in humans (9). No additional information about the specific function of WDR3 exists in the literature; however, chromosome aberrations involving the 1p12 region have been observed in different types of cancer (14 –17). In addition, several studies have shown irregular expression of some of the WD-repeat proteins in certain types of cancers (18 –21). Altogether and based on our earlier association studies (4,5), we hypothesized that WDR3 could be involved in thyroid cancer development.
In this study, we analyzed single-nucleotide polymorphisms (SNPs) from the WDR3 gene in a two-stage study. Lack of association with thyroid cancer was found for individual SNPs; however, haplotype analysis revealed that the WDR3 gene is associated with thyroid cancer risk. In addition, altered expression of the WDR3 gene was detected in thyroid cancer cell lines, by both RT-PCR and Western blot analysis, supporting the possible implication of the WDR3 gene in the etiology of thyroid cancer.
Materials and Methods
Subjects and DNA isolation
This study was performed in two stages in a Spanish population where individuals from case and control groups were unrelated. Both patients and control subjects were recruited during a 5-year period (2004–2008). Patients with thyroid cancer were from the Nuclear Medicine Services at the University Hospital Vall d'Hebron (Barcelona) and the Hospital Josep Trueta (Girona) and were newly diagnosed individuals. Control subjects were healthy, cancer-free volunteer individuals having Caucasian origin with Spanish ancestors. Initially, the study was carried out in a group of 157 patients and 118 control individuals. The sample size was increased to 402 patients and 479 control individuals in the second phase of the study. All subjects selected for this study reported no previous therapeutic or accidental whole-body exposure to radiation. Clinical information of patients was obtained from medical records, and patients were classified as papillary thyroid carcinoma (PTC) group (339 of 402 patients) and follicular thyroid carcinoma (FTC) group (59 of 402 patients). The information about the type of tumor was not available for four patients during the study; consequently, they were classified as others. Informed consent from all participants and clearance from the Ethics Committees of our institutions were obtained.
DNA was isolated from blood samples of each individual using either a standard phenol chloroform method or the QIAamp DNA Blood Mini Kit (Qiagen). Alternatively, DNA of several control individuals was isolated from saliva using the Oragen kit (DNA Genotek).
SNP selection and genotype analysis
In the initial study group of 157 patients and 118 control subjects, a total of 10 SNPs were analyzed, covering the WDR3 gene and its surrounding sequences at 5′ and 3′. The SNPs 1–10 were rs4233455, rs2208375, rs3754127, rs1321665, rs6680844, rs3765501, rs1321666, rs4658973, rs6685906, and rs6678671, respectively. The relative map positions of these markers are shown in Figure 1. SNPs were selected based on information available in public databases, referred to minor allele frequency (>0.2). All these markers are located in the same linkage disequilibrium (LD) block according to the HapMap information, HapMap CEU genotype data, and Haploview software (22), and with a density of one SNP per 4–5 kb.
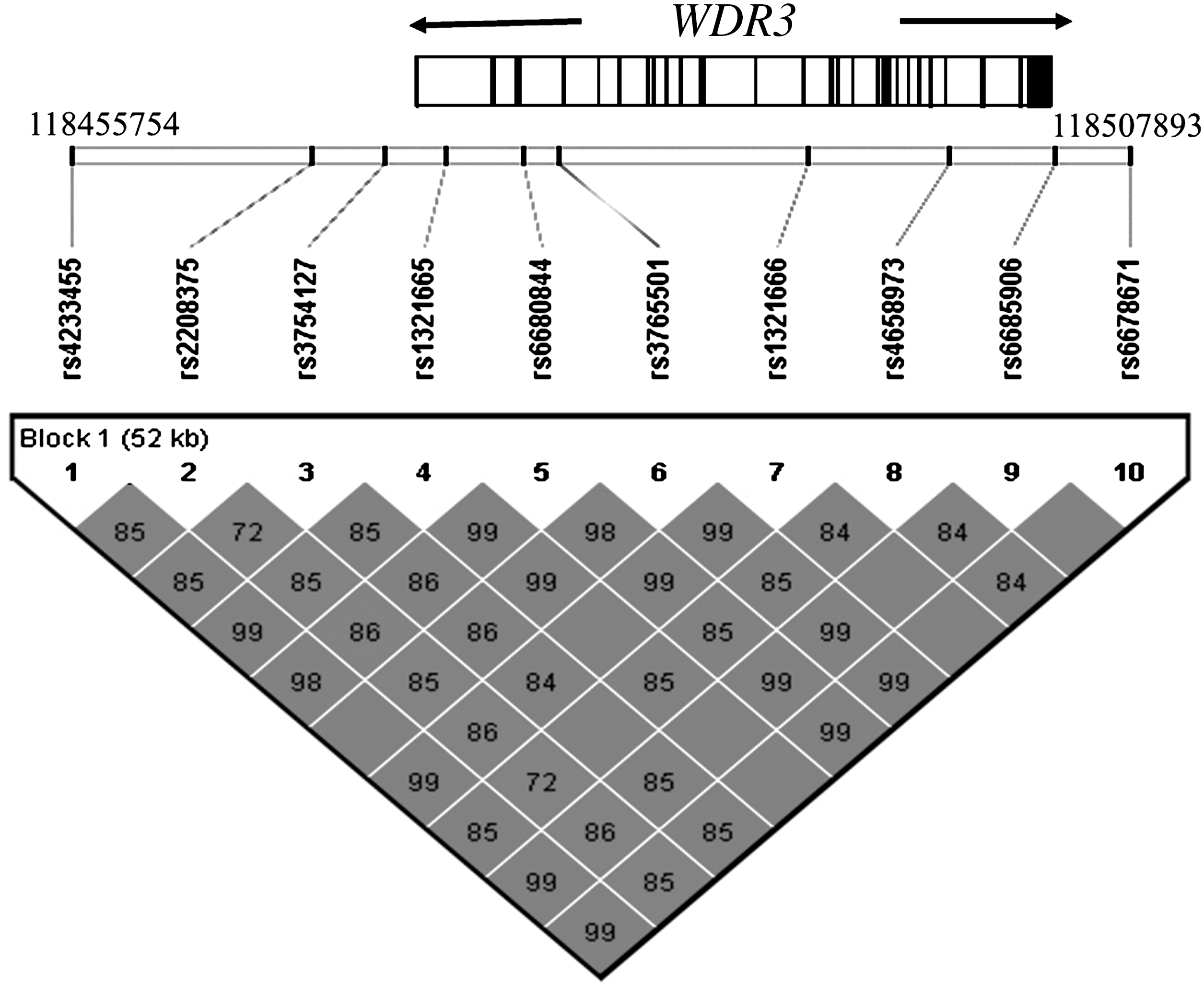
Diagram of the WDR3 gene at 1p12 region, coordinates 118455754 to 118507893. The relative map position of the studied SNPs and pair-wise LD measured using r-squared values are indicated.
In the second genotype phase of 402 patients and 479 control subjects, 3 of the 10 initially selected SNPs were genotyped, rs3754127, rs3765501, and rs4658973 (SNPs 3, 6, and 8, respectively).
All genotype analyses were performed at the Genotype National Centre of Spain (CeGen) using the iPLEX technique (Sequenom, Inc.). To assure the genotyping reliability, 10% of the samples were randomly selected and double genotyped by replicates in multiple 96-well plates. In addition, two HapMAp reference trios were incorporated in plates, and the genotype concordance and correct Mendelian inheritance were verified.
That the first genotyped group of 157 patients and 118 control subjects was also genotyped in the second phase for three repeated SNPs was an additional validation of the genotyping procedure.
Statistical analysis
Chi-square test was used to compare the sex proportion between case and control groups, and to examine the Hardy–Weinberg equilibrium of the genotype distribution in the control population with a 5% level of significance. Mean ages of patients and control subjects were compared by the Mann–Whitney test. Unconditional logistic regression analyses, adjusted for sex and age, were performed to calculate the odds ratios (OR) and 95% confidence intervals (CI) for risk of thyroid cancer. A model of multiplicative allelic effect was assumed (codominant model).
Haploview software (22) was used to examine the LD between SNPs. A logistic regression analysis was used to obtain the adjusted OR and 95% CI of haplotypes. The relation of haplotypes and clinicopathological variables in patients was evaluated by the chi-square test.
All statistical analyses were performed using SNPStats software (23) with a 5% level of significance.
Cell lines
The S11N human thyroid normal cell line, provided by Dr. Ludwig Hieber
RNA extraction and reverse transcription PCR
RNA was extracted using TRIzol Reagent (Invitrogen), according to the manufacturer's instructions. RNA was treated with DNase-I using TURBO DNA-free kit (Ambion). Quality of RNA was verified by agarose gel electrophoresis and quantified with the spectrophotometer (NanoDrop-1000; Termoscientific). Semiquantitative RT-PCR analysis was performed using 100 ng of RNA and the SuperScript™ One-Step RT-PCR System kit (Invitrogen), according to the manufacturer's instructions. Two regions of the WDR3 mRNA, corresponding to exons 3–4 and exons 13–15, were tested in separate RT-PCR reactions, and the endogenous control β-actin mRNA was included in each reaction. Primers for exons 3–4 of WDR3 were F: 5′-TCGATCCGAATCTTCAGTCT-3′ and R: 5′-AACAATGCTTGTGTGATGGC-3′, and amplified a 209 bp sequence from cDNA. Primers for exons 13–15 of WDR3 were F: 5′-TGACACACAGAACATCTTCA-3′ and R: 5′-AATCTGCTGGAGACAATAGA-3′, and amplified a 210 bp sequence from cDNA. The primers for β-actin mRNA were F: 5′-CCAGAGCAAGAGAGGCATCC-3′ and R: 5′-AATCTGCTGGAGACAATAGA-3′. The reaction temperature was 55°C. RT-PCR products were separated by electrophoresis on a 2.5% agarose gel and observed with SYBER green.
Western blot
A total of 30 μg of each protein lysate was heat denaturized in the presence of β-mercaptoethanol (5 minutes, 90°C), loaded onto 7% polyacrilamide sodium dodecyl sulfate gel, and transferred to a nitrocellulose membrane by electroblotting. The membrane was incubated with a monoclonal rabbit anti-WDR3 antibody (Bethyl Laboratories, Inc.) and a secondary anti-rabbit antibody conjugated with horseradish peroxidase (Abcam) at 4°C overnight. Protein bands were observed using the SuperSignal Western Pico Chemiluminescent Substrate kit (Pierce). A vinculin antibody (Abcam) was used as a loading control. The Western blot signal was quantified using GeneTools Analysis Software (Syngene).
Results
In a previous case–control association study carried out in a Spanish population of 227 patients and 202 control subjects, we showed that the 1p12 region was associated with thyroid cancer susceptibility (5). The WDR3 gene maps in this region, and to determine the thyroid cancer contribution of WDR3 we performed a two-stage case–control association study. As DNA samples from individuals participating in the previous studies (4,5) were run out, a new collection of samples was used for this study. Initially, 10 SNPs covering the WDR3 gene and its surrounded regions at 5′ and 3′ were genotyped (Fig. 1) in a group of 157 patients and 118 control subjects. Next, to increase the power of the first study, we genotyped 402 patients and 479 control subjects of the same Spanish population, using 3 of the 10 first genotyped SNPs. In this large group the initial 157 patients and 118 control subjects were also included. All selected SNPs were within the same LD block and showed a minor allele frequency >0.2, based on HapMap and public databases.
In the group of 157 patients (118 women and 39 men) and 118 control subjects (78 women and 40 men), mean age ± SD was 44.44 ± 14.52 and 39.7 ± 13.44 years, respectively (p = 0.013). The sex distribution was 75.1% and 66.1% of women in patients and control subjects, respectively (p = 0.10). In the second genotyped group of 402 patients (312 women and 90 men) and 479 control subjects (283 women and 196 men), mean age ± SD was 47.06 ± 15.48 and 45.99 ± 17.24 years, respectively (p = 0.089), with a sex distribution of 77.6% and 59.1% of women in patients and control subjects, respectively (p < 0.0001). Therefore, before performing the association analysis, the genotype distribution within sex was analyzed in both groups, but no significant differences were found (data not shown). Thus, the different sex proportion in control and patient groups does not influence our association analysis.
In our control population, genotype frequency distribution was consistent with those of the Hardy–Weinberg equilibrium; where p > 0.05 for the 10 selected SNPs (data not shown). The allele frequencies of these SNPs were similar to those reported in the dbSNP database. In the first stage of this study, the logistic regression analysis did not show any significant association for individual SNPs; nevertheless, we could observe that all 10 SNPs exhibited an increased frequency of the homozygous variant genotype in patients compared with control subjects (Table 1). Subsequently, haplotype analysis was performed combining the 10 genotyped SNPs. Four of the inferred haplotypes represented >95% of the individuals, the remaining haplotypes showing frequencies <1% were classified as others. As shown in Table 2 one of the four haplotypes (Hap III, i.e., TACCTACTGC) revealed an increased risk to thyroid cancer though it did not reach statistical significance (OR = 3.23, 95% CI = 0.90–11.61, p = 0.073). To confirm the possible role of WDR3 in thyroid cancer incidence, a second genotype analysis was performed in 402 patients and 479 control subjects in a second phase association analysis. In this case we genotyped only the rs3754127, rs3765501, and rs4658973 SNPs (SNPs 3, 6, and 8, respectively) because, according to our initial study, these polymorphisms served as tag SNPs for haplotype association analysis of the WDR3 gene with thyroid cancer. The haplotypes combining the SNPs 3, 6, and 8 are shown in Table 3. In agreement with our initial results, in this study the CAT haplotype, which corresponds to Hap III of the first study, showed a different distribution between patients and control subjects. The thyroid cancer risk estimated for this haplotype almost reached statistical significance (OR = 1.85, 95% CI = 0.97–3.55, p = 0.063). Stratification of haplotype data by histological type of thyroid cancer revealed a significant association of CAT haplotype with PTC (OR = 2.00, 95% CI = 1.02–3.92, p = 0.044) (Table 3), which likely is a manifestation of the nonstratified data. Moreover, the lack of haplotype association found with FTC might be due to the reduced number of follicular cases.
Adjusted for age and sex.
p-Value corresponding to a codominant model.
CI, confidence interval; MAF, minor allele frequency; n, number of subjects; OR, odds ratios; SNP, single-nucleotide polymorphism.
Adjusted for age and sex.
Rare haplotypes of frequencies <0.01.
Adjusted for age and sex.
Rare haplotypes of frequencies <0.01.
FTC, follicular thyroid carcinoma; PTC, papillary thyroid carcinoma.
Next, we examined if WDR3 haplotypes were related to the clinicopathological characteristics in the patient group. The results are shown in Table 4. No differences in the distribution of haplotypes were found for sex or type of thyroid cancer, PTC and FTC; however, the distribution of haplotypes was different between patients with age of diagnosis ≤45 and >45 years (p = 0.0192).
Haplotypes are equivalent to haplotypes specified in Table 3.
Chi-square test.
On the basis of the association studies described above, we reasoned that, if WDR3 plays any role in thyroid cancer development, expression of WDR3 might be affected in thyroid cancer. To test this hypothesis, we carried out RT-PCR and Western blot analyses in a human thyroid normal cell line, and in nine human thyroid cancer cell lines from different origins: two PTC, three FTC, and four anaplastic thyroid carcinomas. Semi-quantitative RT-PCR analysis was performed amplifying both exons 3–4 and exons 13–15 from the WDR3 mRNA. As it is shown in Figure 2 both segments of the mRNA were successfully amplified in all cell lines analyzed (Fig. 2A). The analysis of WDR3 expression by Western blotting revealed a band of 103 kDa corresponding to the estimated full-length of the protein. Expression of WDR3 in thyroid cancer cell lines was higher than in the thyroid normal cell line, except in CAL-62 (see Fig. 2B). Quantification of the Western blot signal has shown a relative increase of WDR3 in the thyroid cancer cells, with a range of 6–106-fold. Correlation of the Western blot signal with RT-PCR amplification referred to exons 3–4 of the WDR3 cDNA can be observed (Fig. 2). These results show that the WDR3 gene is upregulated in thyroid cancer cells, suggesting its possible implication in thyroid cancer tumorigenesis.
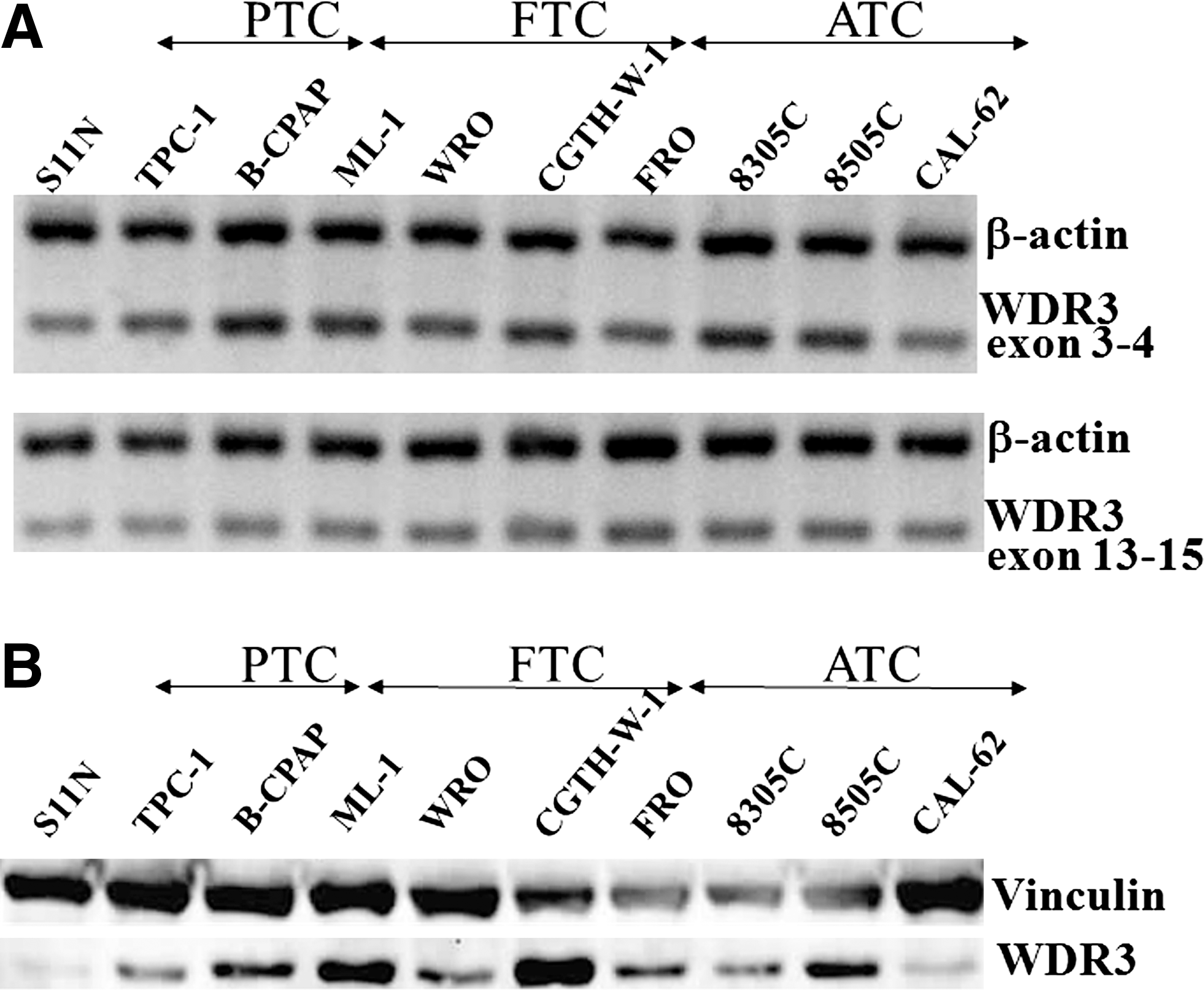
Expression of WDR3 in human thyroid cell lines. (
Discussion
This study was based on our previous finding that suggested the involvement of chromosome 1p12 as a susceptibility region for thyroid cancer (5). Here, we have identified the WDR3 gene, which maps in this region, as a susceptibility locus for thyroid cancer in a Spanish population. Additionally, we have observed altered expression of the WDR3 gene in thyroid cancer cell lines.
The case–control association studies have shown similar results in each of the two phases of the study, and an increased risk to thyroid cancer was revealed by haplotype analysis. In the second phase of the study, the sample size was enlarged to 402 patients and 479 control subjects to ensure enough power for the study, and from the four inferred haplotypes the CAT haplotype showed an OR of 1.85 (95% CI = 0.97–3.55, p = 0.063) (see Table 3). The OR for this haplotype was increased (OR = 2.00, 95% CI = 1.05–3.92, p < 0.044) when only PTC patients were taken into account. Further, a different distribution of haplotypes related with age of diagnosis (p = 0.0192) was observed in the patient group. These results would suggest that WDR3 could play a role in the histological differentiation of thyroid cancer, and in its etiology related to age.
Examination of WDR3 expression in thyroid cancer cell lines revealed that the WDR3 is upregulated in eight of the nine thyroid cancer cell lines used. These results support our association studies indicating that the WDR3 gene can be considered a susceptibility factor for thyroid cancer in the studied population. To our knowledge, this is one of the first studies describing a genetic risk factor for thyroid cancer. Recently, the FOXE1 gene has also been reported to confer thyroid cancer susceptibility, both in sporadic PTC and radiation-related PTC (24,25).
Altogether, our findings will require further work to uncover the biological significance of WDR3. Although the specific function of WDR3 is unknown, this gene is a member of the WD-repeats genes that are involved in cell cycle progression and signal transduction (12). A role of WDR3 in ribosome biogenesis has just been reported (26). The authors of this report proposed that increased expression of WDR3 will promote cancer cell proliferation though the disruption of ribosome synthesis pathway. Previous reports have also shown that expression of different WD-repeat proteins was altered in some types of cancer (19,21). Overall, these studies suggest that WD-repeat genes could contribute to cancer development.
In conclusion, here we provide evidence indicating that WDR3 is a susceptibility gene for thyroid cancer. The investigation was done in a relative large group of patients from a Spanish population, but other populations should be investigated to determine the implication of WDR3 in thyroid cancer risk of the general population. This study also shows that WDR3 is upregulated in thyroid cancer cells, which could have consequences in prevention and treatment of thyroid cancer. Therefore, additional studies to understand the possible role of WDR3 in the etiology of thyroid tumorigenesis are required.
Footnotes
Acknowledgments
The authors thank all the subjects participated in this study; the members of the Nuclear Medicine Service, Hospital Vall d'Hebron (Barcelona), and the Endocrinology Unit of the Hospital Josep Trueta (Girona) for providing patient blood samples; and Dr. R. Melillo and Dr. M. Santoro, University of Naples (Italy), Dr. R. Ciampi, University of Pisa (Italy), and Dr. Ludwig Hieber, German Research Center for Environmental Health (Germany), for sending some of the thyroid cancer cell lines. Gisselle Pérez, Aida Baida, Eddy González-Flores, and Cristian Valiente actively participated in collecting and preparing the samples for genotype analysis. This work was partially funded by the Spanish Ministry of Education and Science (project SAF2007-6338) and the Generalitat de Catalunya (CIRIT, 2009SGR-725). A. Akdi and E.-M. Giménez were supported by a predoctoral fellowship from the Universitat Autònoma de Barcelona.
Disclosure Statement
The authors declare that no competing financial interests exist.