
Abstract
Background:
Adult hypothyroidism is a highly prevalent condition that impairs processes, such as learning and memory. Even though tetra-iodothyronine (T4) treatment can overcome the hypothyroidism in the majority of cases, it cannot fully recover the patient's learning capacity and memory. In this work, we analyzed the cellular and molecular changes in the adult brain occurring with the development of experimental hypothyroidism.
Methods:
Adult male Sprague-Dawley rats were treated with 6-propyl-2-thiouracil (PTU) for 20 days to induce hypothyroidism. Neuronal and astrocyte apoptosis were analyzed in the hippocampus of control and hypothyroid adult rats by confocal microscopy. The content of brain-derived neurotrophic factor (BDNF) was analyzed using enzyme-linked immunosorbent assay (ELISA) and in situ hybridization. The glutamatergic synapse and the postsynaptic density (PSD) were analyzed by electron microscopy. The content of PSD proteins like tyrosine receptor kinase B (TrkB), p75, and N-methyl-
Results
: We observed that the hippocampus of hypothyroid adult rats displayed increased apoptosis levels in neurons and astrocyte and reactive gliosis compared with controls. Moreover, we found that the amount of BDNF mRNA was higher in the hippocampus of hypothyroid rats and the content of TrkB, the receptor for BDNF, was reduced at the PSD of the CA3 region of hypothyroid rats, compared with controls. We also observed that the glutamatergic synapses from the stratum radiatum of CA3 from hypothyroid rats, contained thinner PSDs than control rats. This observation was in agreement with a reduced content of NMDAr subunits at the PSD in hypothyroid animals.
Conclusions:
Our data suggest that adult hypothyroidism affects the hippocampus by a mechanism that alters the composition of PSD, reduces neuronal and astrocyte survival, and alters the content of the signaling neurotrophic factors, such as BDNF.
Introduction
Materials and Methods
Induction of hypothyroidism in adult rats
Male Sprague-Dawley rats, weighing 250–280 g, were treated with 0.05% of 6-propyl-2-thiouracil (PTU; Sigma) in drinking water for 20 days to induce hypothyroidism, as described previously (17). A control group of rats received drinking water without PTU. Serum samples from both PTU-treated and normal rats were obtained to analyze levels of thyroid hormones T3, T4, and TSH. Serum levels of total T3 (tT3) and free T4 (fT4) were measured by chemioluminescence in the laboratory of Institute of Advanced Medical Studies (IEMA), Santiago, Chile (18). Radioimmunoassay was used to measure TSH at the Carlos Chagas Filho Institute of Biophysics of Federal University of Rio de Janeiro, Brazil. The levels of thyroid hormones found in our rat serum were similar to those previously reported in the literature (19). All animal work was performed according to the Guide for Care and Use of Laboratory Animals (National Institute of Health), the approval of the Andrés Bello University Bioethics Committee, and the institutional guidelines were overseen by a veterinarian.
Immunofluorescence and apoptosis assays
Coronal brain sections of 30 μm thickness were cut between 2.16 and 4.56 mm from Bregma by cryostate. These sections were fixed and permeabilized in 70% ethanol and then incubated in terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) reaction mixture at 37°C for one hour (Roche). The tissue sections were washed and incubated in blocking solution at room temperature. These tissue sections were then incubated overnight with the primary antibody mix: monoclonal anti–glial fibrillary acidic protein (GFAP) (1:1000; Sigma) and polyclonal anti–microtubule-associated protein (MAP)-2 (1:300; Sigma). Fixed sections were washed, incubated with Cy3-conjugated anti-rabbit immunoglobulin G (IgG) (Sigma; 1:300) and Cy5-conjugated anti-mouse antibody (1:300; Invitrogen) for one hour at room temperature, followed by a one hour phosphate-buffered saline (PBS) wash. Tissue sections were mounted using Prolong Gold® (Molecular Probes) and examined by confocal microscopy (Leica AOBS). To identify the apoptotic neurons or astrocytes, a double immunofluorescence staining for each cell type was performed as described (TUNEL and anti-MAP-2 and TUNEL and anti-GFAP respectively) (20,21). Specificity was confirmed by replacing the primary antibody with the appropriate isotype-matched control reagent, anti-IgG2A, or the IgG fraction of normal rabbit serum (Santa Cruz Biotechnology). Quantification of neuronal and astrocyte apoptosis was performed by counting the total number of nucleus and the number of TUNEL positive nucleus in GFAP and MAP-2 positive cells in the field. For each condition, four rat brains were analyzed and three slices were studied for each brain. Data were expressed as the percentage of apoptotic neurons or astrocytes in the field respectively.
In situ hybridization detection of BDNF mRNA
Brain tissue was fixed by transcardiac perfusion of 3% paraformaldehyde in 0.1 M PBS pH 7.4. After fixation, brains were removed and postfixed at 4°C during 2 hours in the same fixative and then cryoprotected in 25% sucrose solution in 0.1 M PBS at least during 48 hours at 4°C. In situ hybridization studies using a non-isotopic riboprobe were performed as previously described (22), but modified for mounted sections. Briefly, 16 μm thick-sections from experimental and control rats were cut on a Leica cryostat and selected sections from the hippocampus and parietal cortex regions were mounted on poly-
BDNF quantification
The content of BDNF was analyzed in 7 mg of the total telencephalon homogenate. The homogenate was centrifuged at 20,000 g in a MIKRO 22R Hettich Centrifuge at 4°C for 30 minutes. The supernatant was discarded and the pellet was resuspended in a solution containing 137 mM NaCl, 20 mM Tris-HCl (pH 8.0), 1% nonidet P-40 (NP40), 10% glycerol, 1 mM of phenylmethylsulfonyl fluoride, 10 μg/mL aprotinine, 1 μg/mL leupeptine, and 0.5 mM sodium vanadate. The samples were sonicated with three pulses for 15 seconds at intensity 4, with 5-s intervals in a Misonix Sonicator Ultrasonic processor XL. After sonication, samples were centrifuged at 16,000 g for 30 minutes at 4°C in a MIKRO 22R Hettich Centrifuge. Aliquots of 100 μL of the supernatants were mixed with 40 μL of a solution containing 137 mM NaCl, 2.68 mM KCl, 1.47 mM KH2PO4, 8.1 mM Na2HPO4 (pH 7.35), 0.9 mM CaCl2, and 0.5 mM MgCl2. Then, 20 μL of HCl 1 M was added, incubated for 15 minutes and neutralized with 20 μL of 1 N of NaOH. The samples were kept at −80°C until use. The content of BDNF in these extracts was analyzed by BDNF Emax® Immunoassay System (Promega) as described previously (25). Briefly, a 96-well plate was coated with anti-BDNF monoclonal antibody (20 ng/well) and incubated at 4°C overnight. Then, the plate was washed with a buffer containing 150 mM NaCl, 0.05% Tween 20, and 20 mM Tris-HCl (pH 7.6) and blocked with 200 μL of 1× blocking solution, for one hour. Plates were then washed with 150 mM NaCl, 20 mM Tris-HCl, and Tween® 20 0.05% buffer (TBST), and 100 μL of sample was added in triplicate to each well and incubated for 2 hours at 4°C. Then, the plate was washed five times using TBST buffer and 100 μL of anti-human BDNF polyclonal antibody (diluted to 1:500) was added to each well and incubated for 2 hours. The plate was again washed with TBST buffer and incubated with 100 μL of 1:200 dilution of anti-IgG horseradish peroxidase (HRP) conjugate for one hour at room temperature. The plate was washed five times with TBST buffer and antibody binding was evidenced by adding 100 μL Promega 3,3′,5,5′-tetramethylbenzidine solution until color development was evident. This reaction was stopped with 100 μL of 1 N HCl. The absorbance was measured at 450 nm in a plate reader (Nunc-Immuno™). The BDNF concentration was calculated using a BDNF standard and expressed as picogram of BDNF/mg of protein.
TrkB immunohistochemistry
Rats were perfused via the left ventricle using peristaltic pump (WP, Peri-Star), first with PBS, then with a solution containing 4% of paraformaldehyde (Merck), 0.3% of picric acid (Sigma), and 0.1% of glutaraldehyde (Sigma) in PBS pH 7.4. Brains were removed and incubated in the same fixation solution with 10% sucrose (Sigma), and then 30% sucrose solution. Floating coronal brains slices of 30 μm thick were obtained using a cryostat (Leica, CM 1510). Floating sections were incubated with 0.5% hydrogen peroxidase, then with the blocking solution (3% of normal goat serum [Sigma] in PBS, pH 7.4). The floating sections were first incubated with 0.05 μg/mL of polyclonal anti-TrkB (SC-12, Santa Cruz Biotechnology, Inc.), then with 1.5 μg/mL of biotinylated goat anti-rabbit (BA-1000, Vector Laboratories), and with avidin-biotin peroxidase enzyme conjugated (ABC Kit, Vector Laboratories) at a dilution 1:250. The specific binding of the anti-TrkB antibody was revealed after incubating the floating sections with 0.05% (w/v) 3,3′-diaminobenzidine, 0.01% H2O2, and 0.25% nickel chloride in PBS. The tissues sections were analyzed in a Leica DM 2000 microscope. Three brain slices were analyzed per rat and three rats were used in this study.
Isolation of PSDs
PTU-treated and control male rats were decapitated with a guillotine and their brains were isolated. The telencephalon was dissected, cut, and homogenized on ice in 8 mL of homogenization buffer (0.32 M sucrose, 0.5 mM EDTA, and 5 mM Tris pH 7.4), using 12 strokes with a 40 mL Tissue Grind Potter with Teflon Pestle (Thomas Scientific). One milliliter of each brain homogenate was saved to analyze the total amount of protein. Then, the telencephalon homogenate was centrifuged at 1000 g for 10 minutes at 4°C, using a Sorvall centrifuge 5B-plus with a SS-34 rotor. The supernatant (s1) was saved and the pellet (P1) was washed with homogenization buffer and manually homogenized on ice using Tissue Grind Potter. Then, this fraction was centrifuged at 1000 g for 10 minutes at 4°C. The second pellet obtained (P2) was discarded and the supernatant (s2) was mixed with s1. s1+s2 were centrifuged at 12,000 g in a Sorvall 5B-plus, with a SS-34 rotor, for 20 minutes at 4°C. The third pellet (P3) was saved and rinsed with solution A (0.32 M sucrose, 5 mM Tris-HCl pH 8.1, 0.5 mMethylene glycol tetraacetic acid, and 1 mM dithiotreitol) and manually homogenized with a 17 mL Tissue Grind Potter with Teflon Pestle (Thomas Scientific). The homogenized P3 was loaded on a discontinuous sucrose step gradient (0.32, 1, 1.2 M sucrose in 5 mM Tris-HCl pH 8.1) and centrifuged at 150,000 g for 2 hours at 4°C, using a AH629 Sorvall rotor. The synaptosome 1 fraction (Syn1) was isolated from 1 to 1.2 M fraction of the sucrose gradient, diluted 10 times with lysis buffer (5 mM Tris-HCl pH 8.1 and 0.5 ethylene glycol tetraacetic acid) and lysed by gently mixing with magnetic agitation on ice for 30 minutes. Then the lysed Syn1 fraction was centrifuged at 33,000 g for 30 minutes in a Sorvall 5B plus rotor SS34. The pellet obtained after this centrifugation step (P4) was resuspended in 3 mL of solution A and manually homogenized. Then P4 was loaded on a discontinuous sucrose step gradient (0.32, 1, and 1.2 M sucrose in 5 mM Tris-HCl pH 8.1) and centrifuged at 250,000 g for 2 hours at 4°C, using the AH629 Sorvall. The fraction of synaptosomes 2 (S2) was obtained from 1 to 1.2 M fractions. S2 was saved and diluted with eight volumes of 0.32 M sucrose, 0.025 mM CaCl2, 1% Triton X-100, 2 mM dithiothreitol, and 10 mM Tris-HCl pH 8.1. Then, the S2 was gently mixed in this buffer and centrifuged at 33,000 g (Sorvall RC5B plus rotor SS34) for 30 minutes at 4°C. The fifth pellet (P5) is the PSD fraction. This fraction was washed with 50 mM HEPES pH 7.4 and centrifuged at 250,000 g (Beckman Optima TLX, rotor TLA110) for 10 minutes at 4°C. The sixth pellet was resuspended in 50 mM HEPES pH 7.4, frozen in liquid nitrogen, and stored at −80°C until use.
Immunoblot for postsynaptic proteins
Protein concentration in the PSD fraction, and total telencephalon homogenate samples were determined by the bicinchoninic acid method, as described previously (26). The PSD and total homogenate samples were diluted 1:1 with loading buffer and heated at 100°C for 5 minutes. Then, 20 and 40 μg of protein, respectively, were loaded on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), resolved by electrophoresis, and electrotransferred to a nitrocellulose membrane (Thermo Scientific). The nitrocellulose membranes were incubated one at a time with (i) 1/1000 dilution of anti-NR1 antibody (Chemicon, catalog no. AB1516) for 1 hour; (ii) 1/1000 dilution of anti-NR2A/B antibody (Chemicon, catalog no. AB1548W) for 1.5 hours at room temperature; (iii) 1/1000 dilution of anti-P75 antibody (Upstate, catalog no. 07-476) for 1 hour at room temperature; (iv) 1/1000 dilution of anti-PSD-95 antibody (Chemicon, catalog no. MAB377) for 16 hours at 4°C; (v) 1/1000 dilution of anti-TrkB antibody (Transduction, catalog no. 610102) for 16 hours at 4°C; and (vi) 1/1000 dilution of anti-β-tubulin antibody (Sigma, catalog no. T8660) for 16 hours at 4°C. A HRP-conjugated goat anti-rabbit IgG (Calbiochem, catalog no. 401215) or HRP-conjugated goat anti-mouse IgG (Calbiochem, catalog no. 401315) was used as secondary antibody. Both of these secondary antibodies were diluted to 1/1500 and incubated with the nitrocellulose for one hour at room temperature. The immunoblots were revealed using the enhanced chemioluminescence Western blot detection system (ECL, Amersham).
Immunoprecipitation
The NR2A subunit of the NMDA receptor was immunoprecipitated from synaptosome 2 fraction (S2) obtained from PTU-treated and control rats, to analyze the composition of NMDA receptor in this fraction. The S2 isolated was denatured and solubilized as previously described (27). Briefly, S2 was centrifuged at 100,000 g for 30 minutes at 4°C (Beckman TLX, rotor TLA110). The pellet was resuspended and solubilized at 37°C for one hour in radioimmunoprecipitation assay buffer containing 50 mM Tris pH 9.0, 150 mM NaCl, 1% bovine serum albumin, 1 mM EDTA, 1% NP40, 1% sodium deoxycholate, 0.1% SDS, 1 mM phenylmethylsulfonyl fluoride, 3.3 μg/mL leupeptin, 0.1 mM sodium fluoride, 0.5 mM sodium o-vanadate, and 0.5 mg/mL aprotinin. Then samples were centrifuged at 100,000 g for 60 minutes at 4°C (Beckman TLX, rotor TLA110). The supernatant of S2 (detergent soluble portion of synaptic membranes) was dialyzed overnight at 4°C in 0.32 M sucrose, 0.05 mM Tris pH 7.5, 10 μM EDTA, 0.1% Triton X-100. In parallel, 5 μg of an antibody against the NR2A subunit (Zymed, catalog no. 32-0600) was preincubated with 50 μL of protein G-sepharose beads (GE Healthcare, catalog no. 17-0618-01) in 0.5 mL of 0.05 M Tris-HCl pH 7.5; 10 μM EDTA; and 0.1% Triton X-100. The dialyzed samples were centrifuged at 100,000 g for one hour at 4°C (Beckman TLX, rotor TLA110). The supernatants were incubated with protein G-sepharose beads for 60 minutes at 4°C with agitation. After this treatment, the supernatants were centrifuged at 6000 g for 5 minutes, and half of the supernatant volume was added to protein G-sepharose-antibody complex (General Electric) and incubated for 5 hours at 4°C with agitation. The beads were washed twice in buffer that contained 0.05 M Tris-HCl pH 7.5, 10 μM EDTA, and 0.1% Triton X-100, and twice with the same buffer plus 0.5 M LiCl. The beads and samples of supernatant were boiled for 5 minutes in loading buffer (two-fold concentration). Finally, the samples were analyzed by Western blotting. The content of each protein was quantified by a densitometric analysis of each detection band. Then these values were corrected by the loading factor. The loading factor was obtained from the densitometric analysis of the tubulin band.
Transmission electron microscopy
Rats were perfused via left ventricle with 0.1 M PBS pH 7.4 followed with 2% of glutaraldehyde and 4% of paraformaldehyde in PBS. The hippocampi were dissected from the telencephalum and incubated by immersion with the same fixative solution. Tissue blocks were postfixed in 3% glutaraldehyde in 0.1 M sodium cacodylate buffer pH 7.2 for 6 hours, followed by fixation in 1% of osmium tetroxide. Then the hippocampi were stained with 2% of uranyl acetate aqueous followed by dehydration with ascendant acetone solution and subsequently were pre-embedded in epon:acetone 1:1 and then with pure epon. Semithin sections of 1 μm were stained with 1% toluidine blue. Thin sections of 80 nm hippocampi were stained with 4% of acetate.
PSDs were purified as described in the PSD isolation protocol. The PSD pellets were collected and fixed in PBS with 2.5% of glutaraldehyde. Then, the PSD were centrifuged at 250,000 g for 12 minutes (Beckman TLX, TLA110 rotor). The pellet was postfixed with 1% osmium tetroxide. After this the samples were stained with 2% of uranyl acetate aqueous. PSD were dehydrated in ascendant alcohol and embedded in EPON812 resin. PSD sections between 0.6 and 0.7 nm thicknesses were cut using ultramicrotome (Sorvall, model MT2). The CA3 hippocampi and the PSDs fractions were observed and microphotographed with Zeiss EM-109 transmission electron microscope Philips Tecnai 12 Bio Twin at 80 kV at the Unidad de Microscopía Electrónica of the Pontificia Universidad Católica de Chile.
Statistical analyses
Unpaired two-tailed t-tests were performed on the experimental data shown here. The Sigma Plot9 plus sigma Stat 3.5 program Systat Software, Inc. was used for the analyses. The results were considered to be significantly different when p<0.05.
Results
Hypothyroidism induction in adult rats
After 20 days of PTU treatment, serum samples were collected from PTU-treated and control rats to analyze the serum levels of tT3, fT4, and TSH (Fig. 1). PTU-treated rats showed significantly reduced serum levels of fT4 (control: 1.4±0.4 ng/mL; PTU-treated rats: 0.4±0.2 ng/mL, n=3, p<0.05; Fig. 1A), tT3 (control: 92.9±1.9 ng/dL; PTU-treated rats: 51.8±3.9 ng/dL, n=3, p<0.05; Fig. 1B) and significantly increased TSH serum levels (control: 2.5±1.3 ng/dL and PTU-treated rats: 23.8±5.9 ng/dL, n=3, p<0.05) compared with normal rats (Fig. 1C). Based on the clinical definition of hypothyroidism, the serum levels of tT3, fT4, and TSH indicated that the PTU treatment induced a hypothyroid condition in these animals.
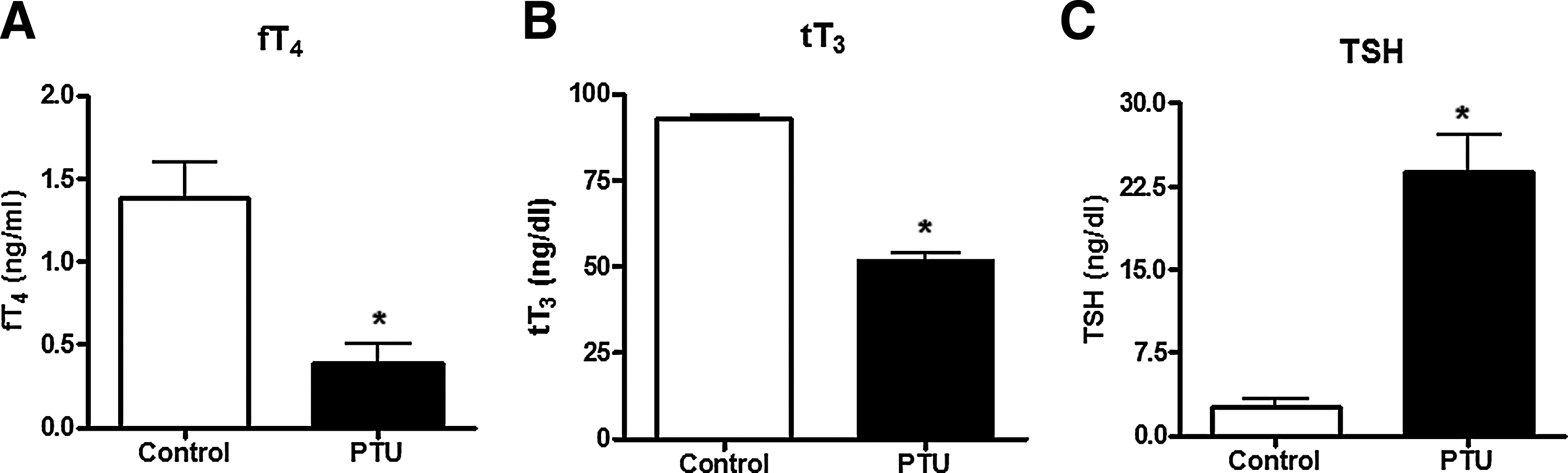
PTU treatment induces hypothyroidism in adult rats. Serum levels of fT4, tT3, and TSH were determined in adult rats non-treated (control) or treated with PTU for 20 days (see “Materials and Methods”).
Hypothyroidism promotes apoptosis and gliosis in the adult rat brain
To evaluate whether induction of hypothyroidism in adult rats alters the survival of neurons and astrocytes in the hippocampus, tissue sections were obtained from control and hypothyroid animals and stained for the following marker: TUNEL to determine apoptosis, the neuronal marker MAP-2, the glial marker GFAP to determine gliosis, and a nuclear staining (DAPI). Then, quantification of these parameters was performed by confocal microscopy. As shown in Figure 2A and B, a low level of neuronal and astrocyte apoptosis was observed in the hippocampus of normal adult rats, however, the hippocampus obtained from hypothyroid adult rats showed a significant increased level of apoptosis in neurons and astrocytes. Moreover, we observed that astrocyte apoptosis was higher than neuronal apoptosis and that both were significantly different in hypothyroid rats compared with control rats (Fig. 2B). Negative controls with non-immune sera, IgG1 control or no enzyme for TUNEL staining were included in all experiments (Fig. 2A).
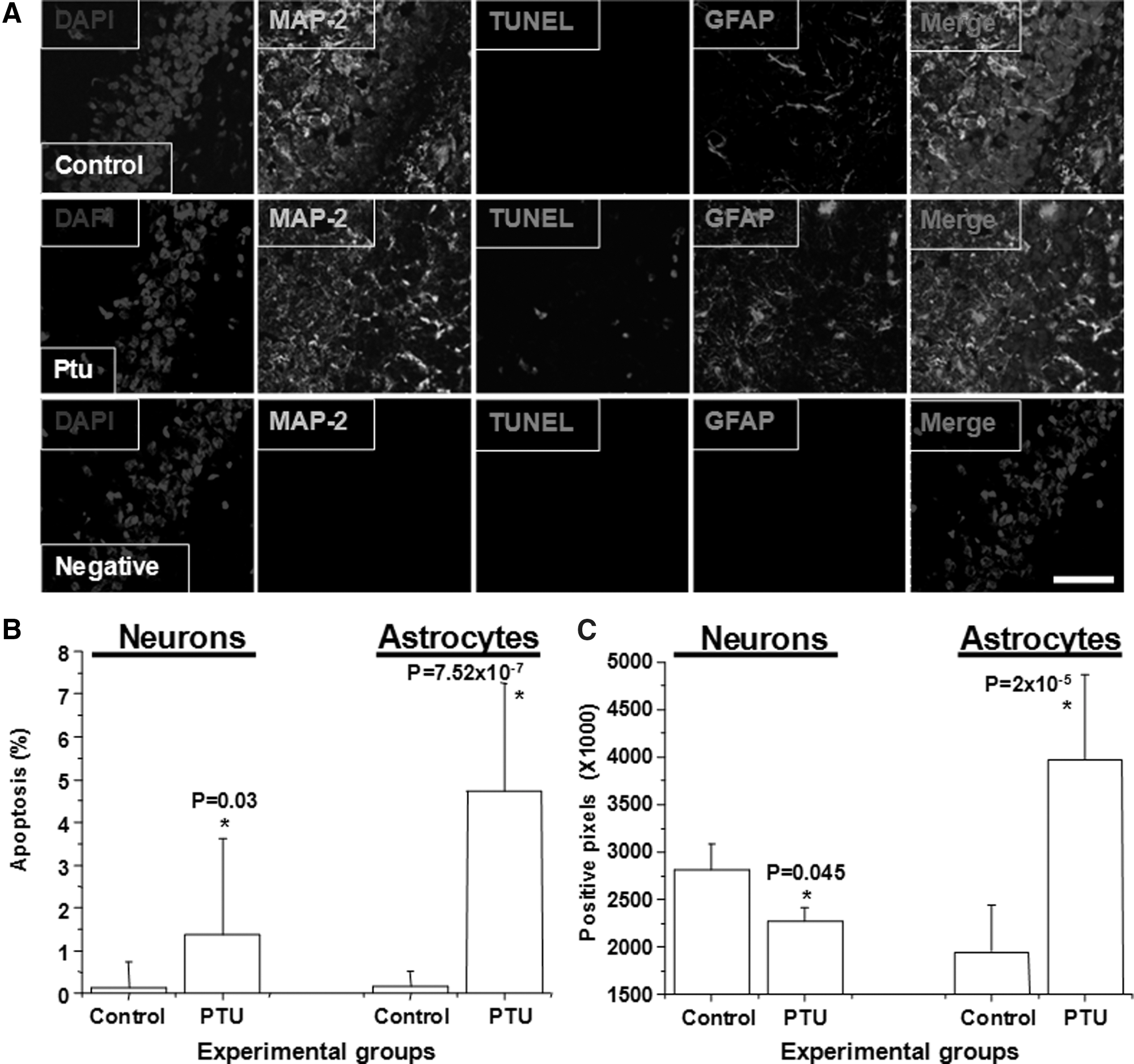
The hippocampus of hypothyroid adult rat present gliosis and apoptosis. Adult rats were treated or not with 0.05% PTU for 20 days. Then the brain was recovered, fixed as described in “Materials and Methods” and analyzed for the presence of specific proteins by confocal microscopy.
Further, using quantification of positive pixel for each channel, we also observed a reduction in MAP-2 staining in the hippocampus of hypothyroid rats, compared with normal animals (Fig. 2C, neurons). Because the reduction of MAP-2 was higher than the total number of neuronal apoptosis, we suggest that MAP-2 reduction is not only due to neuronal cell death, but also due to changes in length, diameter, and number of neuronal processes. In contrast to the reduction of MAP-2, we observed an increased staining for GFAP in the hippocampus of hypothyroid rats, compared with normal animals (Fig. 2C, astrocytes), suggesting that gliosis occurs in hypothyroid animals.
Hypothyroid adult brains show altered contents of BDNF and TrkB
It has been reported that BDNF could play opposite effects in neuronal survival and synapsis potentiation depending in part on which type of receptor is expressed in the target cell (28). TrkB and p75 are the specific and non specific receptors for BDNF, respectively (28). Therefore, we measured both the mRNA and protein expression level of BDNF and its receptors TrkB and p75 in the brain of PTU-treated and control rats. The BDNF mRNA level was studied by an in situ hybridization assay, using the exon V as the hybridization target, which is the common exon among the four different forms of BDNF mRNAs. The hybridization signals for BDNF exon V mRNA showed a significant increase in layers II, III, and V of the parietal cortex, dentate gyrus, CA1, and CA3 hippocampal regions of PTU-treated animals, compared with the expression levels shown by control animals (Fig. 3A, B). In agreement with this result, we determined by enzyme linked immunosorbent assay (ELISA) that BDNF peptide level was increased fivefold in telencephalon derived from PTU-treated rats, compared with control animals (PTU-treated: 252.16±78.43 pg/mg protein, n=5 and controls: 47.49±18.58 pg/mg protein with n=7; Fig. 3C).
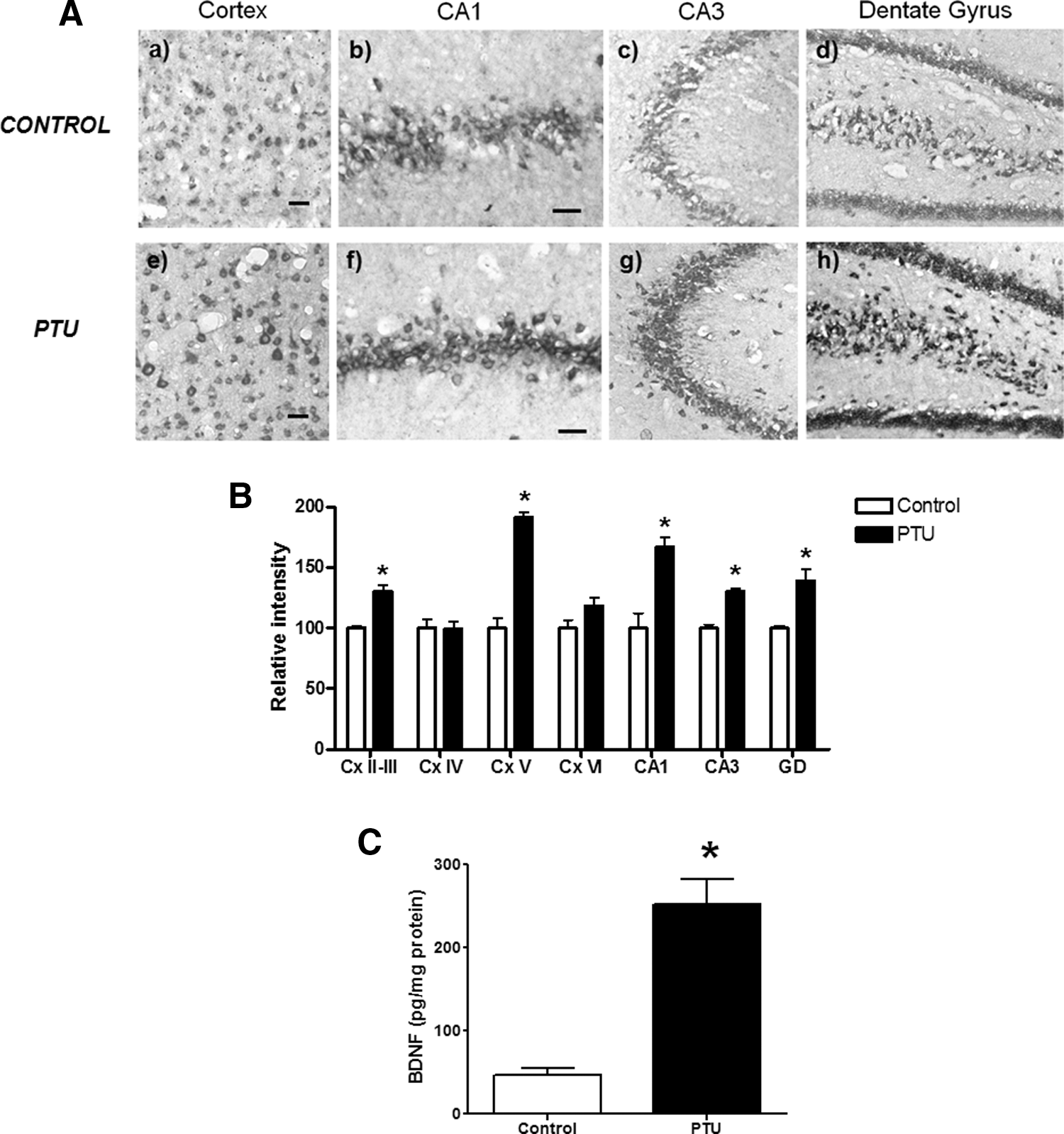
Increased expression of BDNF exon V in the hippocampus and parietal cortex of hypothyroid adult rats.
We also analyzed, by Western blot, the content of the BDNF receptors, TrkB and p75, in the total brain homogenate and in the PSD fractions. As shown in Figure 4A and B, we observed a reduction in TrkB content in the PSD fraction of PTU-treated rats, compared with controls (68±2.5% of TrkB content in PTU-treated rats relative to controls p<0.01, n=3). Conversely, we observed higher levels of TrkB in total telencephalon homogenate from PTU-treated rats, compared with controls (131±15% of TrkB content in PTU-treated rats relative to controls p<0.01, n=5) (Fig. 4A, B). Importantly, the content of p75 receptor in the PSD and total homogenate was similar in control and PTU-treated rats (Fig. 4A, B). Finally, we analyzed the TrkB content by immunohistochemistry in the same brain areas where BDNF was analyzed (Fig. 4C). We observed a reduced stain for this protein in the dendrites of neurons at the layer V of the neocortex, CA1, CA2, CA3, and dentate gyrus.
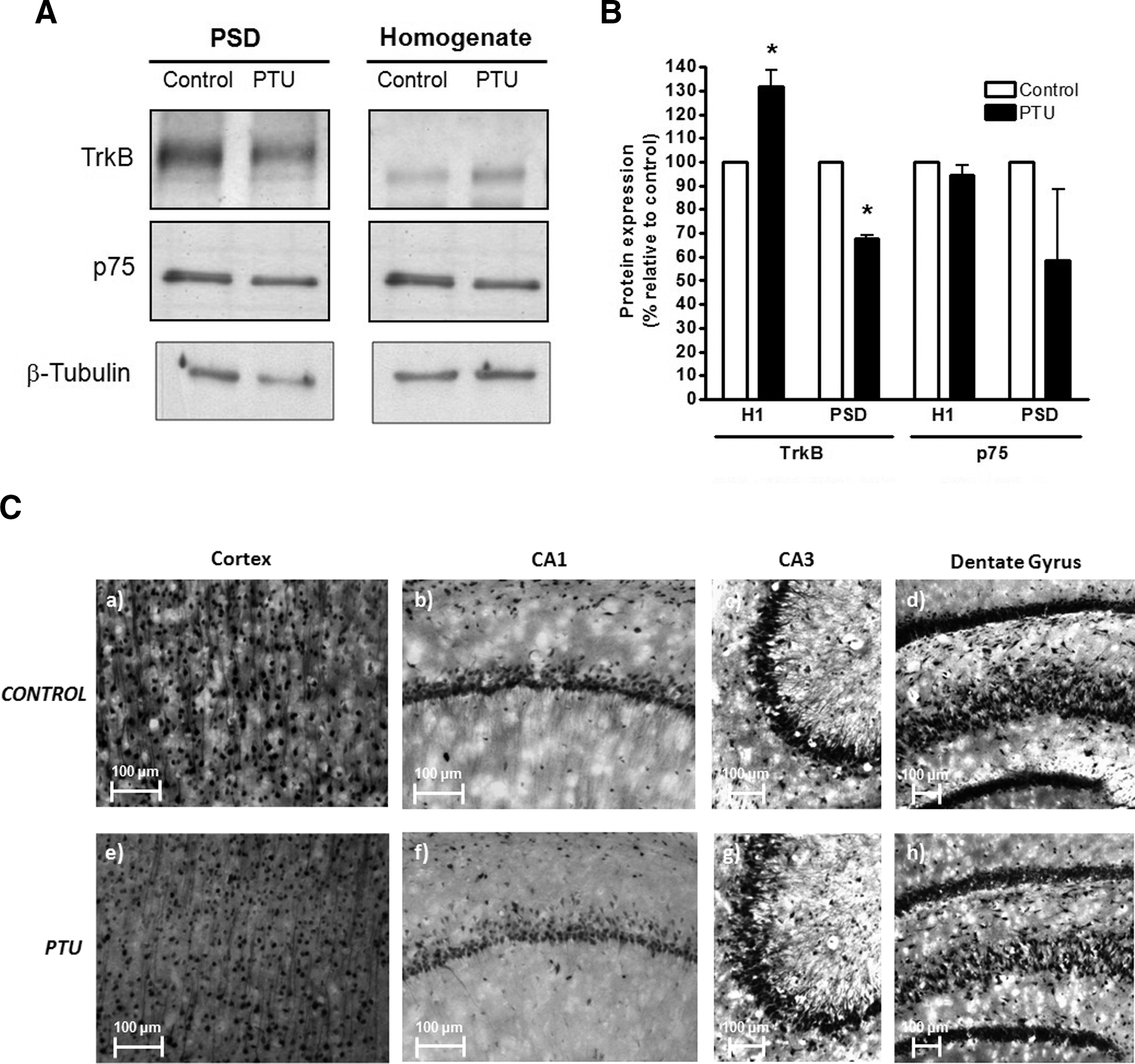
Reduction of TrkB in the PSD of adult hypothyroid telencephalon.
Hypothyroidism reduces NMDA receptor contents in PSDs
To evaluate the effect of hypothyroidism on the content of NR1 and NR2A/B subunits of the NMDAr in the PSDs of normal and PTU rats, we performed Western blot and immunoprecipitation analysis in brains of control and PTU-treated rats. As observed in Figure 5A and B, a significant reduction in the content of NR2A/B was observed in the PSDs of PTU-treated rats (NR2A/B content in the PSD: 72±7.63%, p<0.05, n=3). In contrast, we observed a slight but significant increase of this subunit in the total fraction of telencephalon homogenates from PTU-treated rats (NR2A/B content in the total homogenate: 126±13%, p<0.05, n=5).
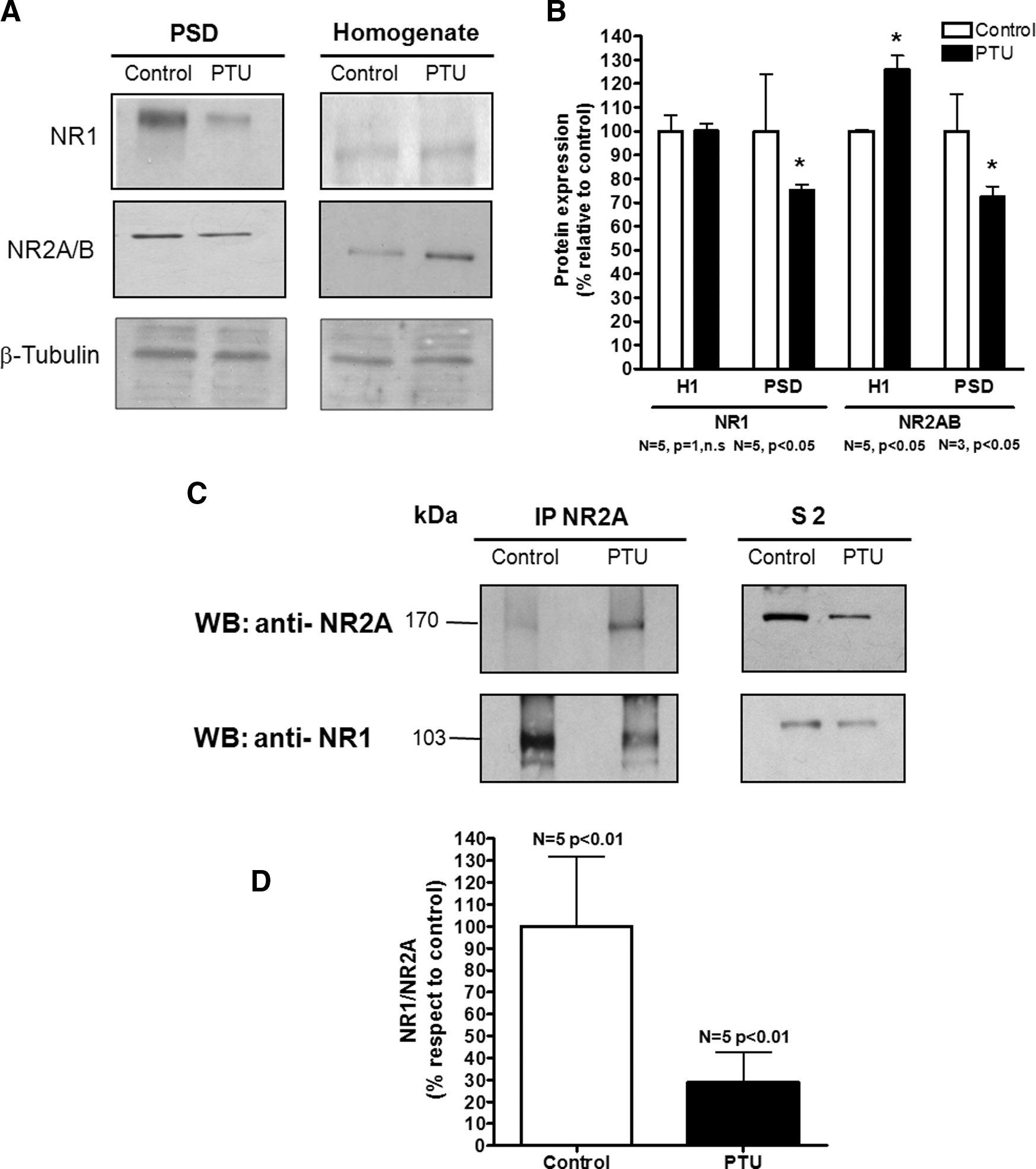
Reduction in the content of NR1 associated with NR2A in the PSD of hypothyroid adult rats.
The NR1 subunit was also significantly reduced in the PSDs from PTU-treated rats (75±5.5%, p<0.05, n=5), but no significant changes were observed in the content of NR1 in the total fraction of telencephalon homogenates (Fig. 5A, B). We also analyzed the association ratio of NR1 and NR2A by co-immunoprecipitation experiments (Fig. 5C, D). NR2A was immunoprecipitated from S2 fraction (see “Materials and Methods”). NR2A and NR1 contents were analyzed from the NR1 immunoprecipitated fraction by Western blot using specific antibodies directed to NR2A and NR1 subunits (Fig. 5C). The values obtained from densitometric analysis of NR1 and NR2A bands were used to determine the association ratio between both proteins (NR1/NR2A). As shown in Figure 5D, our results show that the association ratio of NR1/NR2A was reduced to 55% in S2 of PTU-treated rats compared with control rats (p<0.01, n=5).
Because a significant reduction in the content of NR1, NR2A/B, and TrkB was observed at the PSD from hypothyroid animals, we analyzed, by transmission electron microscopy, the thickness of PSD in these fractions and also from the glutamatergic synapses of the stratum radiatum of the CA3 (Fig. 6). To assess glutamatergic synapses from the CA3, we studied those synapses located in the proximal region of basal dendrites from CA3 pyramidal neurons. In this region of the stratum radiatum, mossy fibres and commissural synapses are found (Fig. 6A).
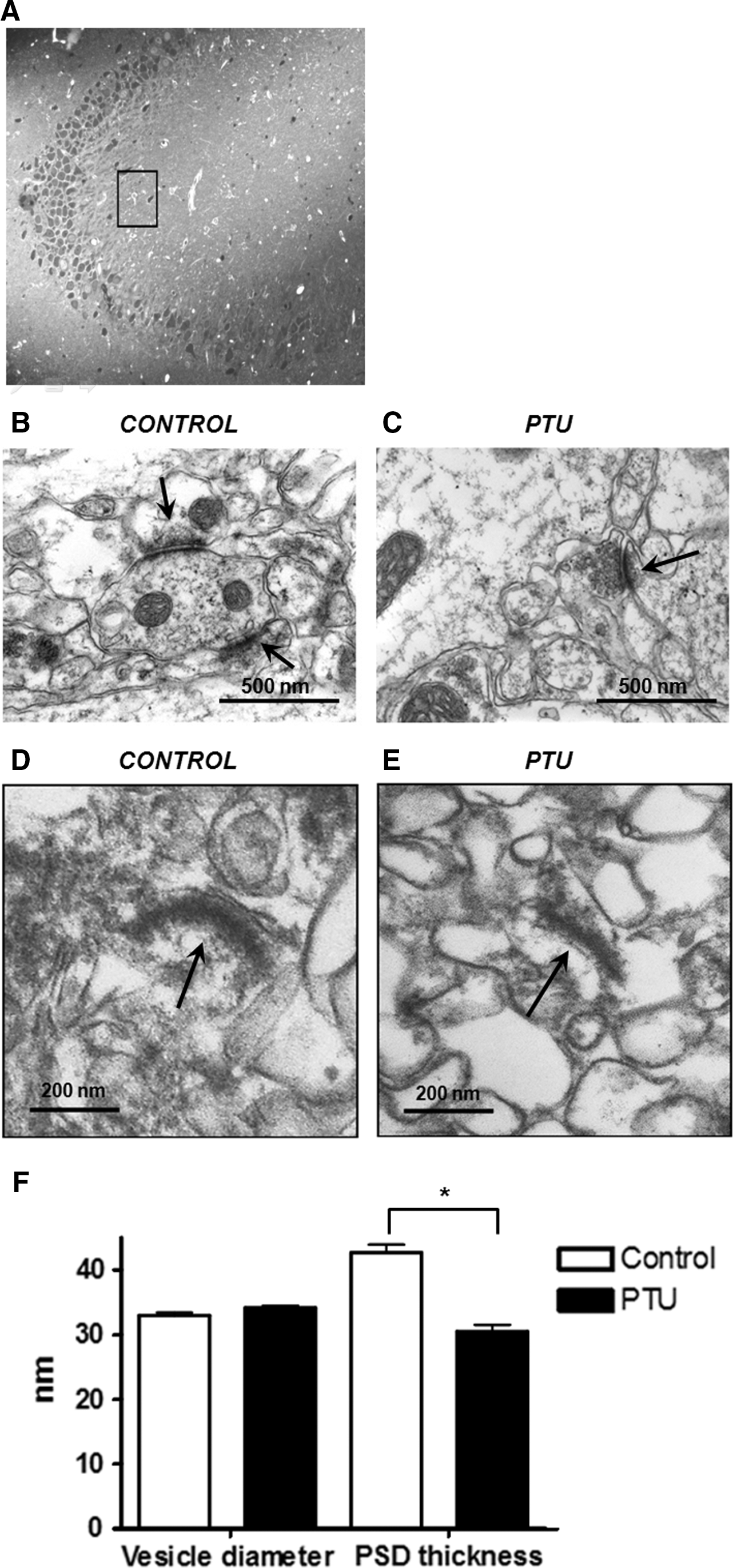
PSD thickness reduction from the CA3 region of hypothyroid adult rats.
Glutamatergic synapses are easily identified by electron microscopy, because they are asymmetrical. Representative pictures of electron micrograph of basal glutamatergic synapses in CA3 region for control and hypothyroid rats are shown in Figure 6B and C, respectively. Representative electron micrographs of PSD fractions are shown from control (Fig. 6D) and hypothyroid rats (Fig. 6E). A quantitative analysis at defined CA3 regions demonstrated a decrease in PSD thickness in CA3 of hypothyroid rats compared with control animals (Fig. 6E). Instead, at the presynaptic regions, the diameter of neurotransmitter vesicle was comparable between hypothyroid and control rats (Fig. 6E).
Discussion
In this study we have shown evidence, at the cellular and molecular level, for the deleterious effects of hypothyroidism in adult brain regions like the hippocampus. Our findings contribute new evidence indicating that adult hypothyroidism is harmful for this organ (29). Our results showed an increase in TUNEL reaction in the hypothyroid CA3 hippocampal region, which supports the notion that there is apoptosis process in this area. Consistent with this finding is the report of Alva-Sanchez et al. (16), showing an increase in the expression of apoptotic signaling molecules such as Bax/Bcl2 and cell damage in the CA3 hippocampal region during hypothyroidism. Our finding is not only in agreement with this data, but also indicates that apoptosis in the CA3 region of hypothyroid hippocampus occurs in neurons and astrocytes. In fact, apoptosis is not restrictive to the CA3 region because we also detected increased levels of apoptosis at CA2 region (data not shown). In agreement with our data, two other studies have shown neuronal apoptosis in other hippocampal regions (e.g., dentate gyrus) (30,31).
We also found that the CA3 region of the hippocampus of hypothyroid rats displays an increase in GFAP staining, suggesting that this region has astrogliosis. This is in agreement with a study performed by Stoica et al. (32), who also reported astrogliosis in inherited tertiary hypothyroidism. The presence of both astrogliosis and apoptosis has been described in Alzheimer disease (33), Parkinson disease (34), and HIV infection (35). In all these conditions, it has been suggested that astrogliosis together with apoptosis is a sign of brain inflammation. It will be of interest to evaluate whether astrocytes are secreting chemokines or cytokines that could induce an inflammatory process in the CNS of the hypothyroid rat.
Because BDNF is a neurotrophic factor involved in neuronal protection, cell death, and synaptic plasticity, we also analyzed the brain BDNF content of hypothyroid rats (28). We detected an increase in BDNF in hypothyroid rats. Specifically, BDNF mRNA increased in all hippocampus regions and at the layers II, III, and V of the neocortex. On the other hand, the quantities of BDNF mRNA in the IV and VI neocortex layers were similar in hypothyroid and control rats. These results are not in agreement with reports that BDNF mRNA levels are similar in all brain regions of hypothyroid animals (15). We believe that these differences are due to differences in the probes used to detect BDNF mRNA. Notably, it has been shown that there are eight possible transcripts for BDNF (36) and in the in situ hybridization analysis performed in this study we used a cRNA antisense probe specific for the coding exon V; this detects all possible BDNF transcripts. We also analyzed BDNF peptide expression, because this peptide is regulated at the transcriptional level and it is also highly regulated at the transduction and post-secretion level (37). We found that BDNF peptide increased fivefold in the telencephalon of hypothyroid rats. This is consistent with the mRNA results. These results suggest that, in the hypothyroid state, there is an increase in BDNF production, probably as a response to a cellular stress.
The content of BDNF peptide was analyzed by ELISA; this assay does not discriminate between BDNF and its precursor ProBDNF. This is an important point because both molecules are co-secreted but ProBDNF has biological activity per se that could be different to that of BDNF (38). Regarding this, it has been proposed that ProBDNF has an action opposite to that of BDNF and that its action is mediated by p75 receptor (39). Instead, BDNF preferentially interacts with TrkB (37). In the present study, we found that the content of p75 was similar in hypothyroid and control rats, but the content of TrkB in PSDs was decreased in the telencephalon of hypothyroid rats. Consistent with this result is the observation of reduced stain for TrkB at the dendrites and soma of pyramidal neurons in the hippocampal regions of hypothyroid rats. Given that our results have shown that BDNF increases in the hippocampal regions of hypothyroid rats and TrkB decreases, it is possible that TrkB could be downregulated at synaptic contacts in the hippocampus of these rats. In fact, TrkB was decreased in PSDs but its content was higher in the whole telencephalon of hypothyroid rats compared with controls. Downregulation of TrkB, due to high levels of BDNF, has been reported (40). Thus, the observation of reduced amounts of TrkB, in the PSDs that contain normal amounts of p75 suggests that, in the hippocampus of hypothyroid rats, the action BDNF is mediated mainly by p75 and not by TrkB. This is relevant since the capacity to play opposite roles in neuronal survival and synapse potentiation, depending on the type of receptor that is preferentially expressed at the surface of the target cell (TrkB or p75), has been attributed to BDNF (28,41). It has also been reported that the interaction of BDNF or ProBDNF with p75 has deleterious effects on glutamatergic neurons, for example apoptosis induction and damaging of glutamatergic synapses (41). With respect to glutamatergic synapses, we found a reduction in the content of NR1 and NR2A/B NMDA subunits in the PSD fractions and in the number of NR1 subunits associated with the NR2A subunit, suggesting that PSDs in the hypothyroid state contained reduced amounts of NMDA receptors. A reduction in the number of NMDA receptors at the postsynaptic neuron has been associated with impaired glutamatergic synaptic function (42). Even though, the sizes of the vesicles were similar in hypothyroid and control glutamatergic synapses (Fig. 6D), PSDs were significantly thinner in hypothyroid rats than control rats. The reduction in the PSD thickness correlated with the drop in the content of NR1, NR2A/B, and TrkB in the PSD fraction, supporting the concept that glutamatergic synapses in this region could be impaired.
In summary, in this study we found apoptosis, gliosis, and alterations in PSDs in the hippocampus of hypothyroid rats, supporting the notion that hypothyroidism is harmful to this brain region.
Footnotes
Acknowledgments
The authors are supported by grants FONDECYT no. 1100926, FONDECYT no. 1070352, FONDECYT no. 1085281, FONDECYT no. 1100926, FONDECYT no. 3070018, FONDECYT no. 3100090, FONDECYT no. 11075060, and FONDECYT no. 1100926, Millennium Institute on Immunology and Immunotherapy (P-09-016-F), Millennium Nucleus (P07-011-F) and Proyecto Interno Universidad Andrés Bello DI-01-44-08.
Disclosure Statement
No competing financial interests exist.