
Abstract
Background:
Gap junctions are membrane structures composed of connexins (Cx) that allow diffusion of small molecules between cells. They are involved in tissue homeostasis, and various organ dysfunctions have been associated with gap junction defects. To verify their possible involvement in thyroid pathologies, the expression of connexin43 (Cx43), the major Cx in the human thyroid, was evaluated in a variety of diseases including cancer.
Methods:
There were 122 samples from various thyroid pathologies that were collected to analyze the presence of Cx43 by immunofluorescence. Through confocal microscopy, different patterns of Cx43 localization were identified as normal (membrane) or abnormal (cytoplasmic or lack of detection). The analysis of Cx43 expression was further performed by quantitative reverse transcriptase–polymerase chain reaction and immunohistochemistry in a subset of 25 papillary carcinomas and compared with nontumoral thyroid tissues.
Results:
The presence of Cx43 was commonly altered in thyroid cancer, as abnormal Cx43 staining was detected in 94.1% of cancer, 47.4% of adenomas, 45.7% of multinodular goiter, 16.7% of Graves' disease, and 25% of thyroiditis. In papillary carcinoma samples, the deregulation of Cx43 expression was mostly the consequence of a decrease of Cx43 mRNA (68% of cases) when compared with normal tissue. When Cx43 mRNA was not downregulated (32% of cases), both loss of membrane staining and aberrant cytoplasmic distribution of the protein were observed.
Conclusions:
These results show that aberrations of Cx43 expression are associated with thyroid papillary carcinoma.
Introduction
A few studies have addressed the roles of gap junctions in the human thyroid. An approach to understand their role is considering the in situ expression and localization of Cx in thyroid diseases. Therefore, since gap junctions are present in the thyroid and localized at cell–cell contact areas of follicular cells, we explored whether thyroid pathologies are associated with disturbances of Cx expression. To do so, we analyzed the expression of the major Cx expressed in human thyroid, connexin43 (Cx43), in a large panel of thyroidectomy samples representing various unrelated pathologies from thyroiditis to cancer. Since confocal analysis commonly identified abnormal patterns of Cx43 expression (cytoplasmic localization or lack of detection) in papillary carcinomas, the analysis of Cx43 expression was further pursued by quantitative real-time reverse transcriptase (RT)–polymerase chain reaction (PCR) and immunohistochemistry in an extra subset of 25 tumor samples and compared with their corresponding normal tissues.
Materials and Methods
There were 122 samples of thyroid tissue collected from patients undergoing thyroidectomy between 2001 and 2005 (Service de Chirurgie Viscérale, CHU de Poitiers, France). Since Cx43 was the only Cx type detected in situ, a blind study was routinely conducted independently by two persons to analyze Cx43 expression by immunofluorescence. The definitive diagnosis was retrospectively collected. Exclusion criteria were lack of patient's authorization.
Since Cx43 expression appeared to be commonly altered in thyroid cancer, the analysis of its expression was further pursued by quantitative real-time RT-PCR and immunohistochemistry in an extra subset of papillary carcinoma samples from 25 patients treated between 1998 and 2002 (Service de Chirurgie Viscérale, CHU de Poitiers). All samples (25 tumoral and 16 corresponding normal tissues) were obtained from the Regional Tumor Bank (CHU de Poitiers) in accordance with French regulations and the declaration of Helsinki. Clinical data concerning the patients were retrospectively collected from hospital files.
Preparation of the samples
For immunofluorescence study, the pathologist received the 122 surgical specimens immediately after surgery. Tissue samples were selected from each patient, snap frozen in liquid nitrogen, and stored at −80°C until analysis. For each sample, series of 5μm-thick slices of the tissue were prepared on a glass slide and fixed in pure acetone (10 minutes). The preparations for immunofluorescence were then dried before being processed. A haematoxylin-eosin stain was done for diagnosis.
The tumor specimens used for quantitative real-time RT-PCR and immunohistochemistry were snap frozen after surgery in liquid nitrogen and stored at −80°C in the Regional Tumor Bank until our study.
Indirect immunofluorescence
The preparations were re-hydrated in a bath of phosphate-buffered saline (PBS) without Ca2+ and Mg2+ (pH 7.2; 10 minutes) and incubated for 45 minutes in a blocking solution (3% bovine serum albumin in PBS) for saturating nonspecific sites. After blocking, slides of each preparation were incubated (1 hour) with rabbit polyclonal (Zymed, San Francisco, CA) or mouse monoclonal (Chemicon Int., Temecula, CA) antibodies directed against Cx43 diluted (1:200) in PBS. After washing in PBS, preparations were incubated (1 hour) with corresponding secondary antibodies (Sigma-Aldrich, L'Isle d'Abeau Chesnes, France) coupled either to rhodamine (anti-rabbit secondary antibodies) or to fluoresceine (anti-mouse secondary antibodies). In order to distinguish cells, nuclei were labelled by adding TO-PRO3 (Interchim, Montluçon, France) to the secondary antibody solution (diluted 1:1000). After washing in PBS, preparations were mounted under a coverslip in Vectashield (Interchim) to protect them against photobleaching. After sealing, preparations were kept in darkness (+4°C) before being observed by confocal microscopy (FV-1000 station installed on inverted microscope IX-81; Olympus, Tokyo, Japan).
In parallel, a control slide was used to test any unspecific fluorescence by being only exposed to the secondary antibody coupled to the fluorochrome. The results with the two antibodies (monoclonal and polyclonal) were overlapping.
RNA extraction and cDNA preparation
Tissues from tumoral and corresponding normal tissues were homogenized using tissue lyser (Qiagen, Courtaboeuf, France). Total RNA was extracted using Qiagen RNeasy Mini Kit (Qiagen). The procedure was followed according to the manufacturer's instructions with minor modifications: for exclusion of contaminating genomic DNA, the spin-column membranes were treated with DNase (Qiagen) for 15 minutes before elution. DNase treated total RNA (3 μg) was transcribed into cDNA using Superscript TM II RNase H and random hexamers (Invitrogen, Cergy Pontoise, France).
Quantitative real-time RT-PCR
We assessed Cx43 mRNA amount to the level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA in 41 thyroid tissue samples (25 pathological parts and 16 corresponding normal parts of same tumors when available). This was done using quantitative real-time RT-PCR in the ABIPRISM 7000 Sequence Detection System (Applied Biosystem, Foster City, CA). The amplification of Cx43 and GAPDH cDNAs was carried out in 25 μL reaction volume consisting of 1 × Taqman® Universal PCR Master Mix and 1.25 μL of Taqman® Gene Expression Assay Reagents (Cx43: Hs 00748445 s1, GAPDH: 999999905 m1) containing 900 nM primers and 200 nM probe (Applied Biosystems).
The reaction was performed as follows: 50°C for 2 minutes, 95°C for 10 minutes, followed by 40 cycles at 95°C for 15s, and at 60°C for 1 minute. Each sample was tested in duplicate, and a negative control (water) was included in every plate. The computed tomography value was defined as the cycle number (Ct) at which the fluorescence crossed the threshold. The results were expressed in ΔCT (CtGAPDH − CtCx gene) or in 2−ΔCt × 1000 for the amount of the target normalized and relative to the endogenous reference.
Immunohistochemistry of thyroid papillary samples
Immunohistochemistry was performed on 15 papillary carcinomas and their adjacent normal tissue. Surgical specimens were fixed in formalin, embedded in paraffin, and cut (5 μm). The sections were deposited on a glass slide, deparaffinized, rehydrated, and heated at 100°C for 2 minutes in 10 mM citrate buffer (pH 6.0) for antigen retrieval. Endogeneous peroxidases were blocked with 1% hydrogen peroxide (Sigma-Aldrich). Preparations were then incubated (1 hour) with a mouse monoclonal antibody directed against Cx43 (Sigma-Aldrich) diluted in PBS (1:2000). The immunostaining was performed with a standard immunocytochemical protocol based on an indirect streptavidin-biotin method with biotinylated anti-mouse IgG secondary antibody, streptavidin peroxidase, and diaminobenzidin substrate (Dako Real™ Detection System; Dako, Trappes, France). Preparations were counterstained with hematoxylin.
Data and statistical analyses
Patterns of staining were collected by numbering the samples in a data processor. The definitive diagnosis was communicated at the end of confocal analysis by the pathologist. This diagnosis permitted to group the samples according to the pathology and to compare the immunofluorescence data between themselves. A statistical analysis was done by comparing normal and abnormal expressions among thyroid pathology groups, using Chi square and Fisher's exact test. To do so, a conservative correction for multiple testing (8 chi-square analyses) was made according to the Bonferroni method (23), resulting in an adjusted alpha-level of 0.00625 (= 0.05/8). SAS version 9.1 (SAS Institute, Cary, NC) was used for statistical analysis.
For Cx43 mRNA, the data distribution of normalized expression levels of tissue samples is non-Gaussian. So, nonparametric tests were applied for data analysis, and the Wilcoxon test was used for comparing tumor tissues (T) with their normal surrounding counterparts (S). Upregulation or downregulation of Cx mRNAs in thyroid tumors was defined as follows: T/S = 2−ΔCt tumor/2−ΔCt normal tissue. A normal interval of the T/S ratio was defined for Cx43 mRNA. This normal interval was defined by the 95% confidence interval of mean for normal tissues and was 0.77–1.259 for Cx43. p-Value < 0.05 was considered statistically significant.
Results
Cx expression in human normal thyroid
The in situ immunofluorescence approach was unable to detect gap junctions made of Cx26 and Cx32 in normal human thyroid tissue (data not shown). Only Cx43 was detected at cell-cell contacts of the follicular epithelium (Fig. 1A, B), suggesting that only Cx43 is detected at the protein level in the normal human thyroid.
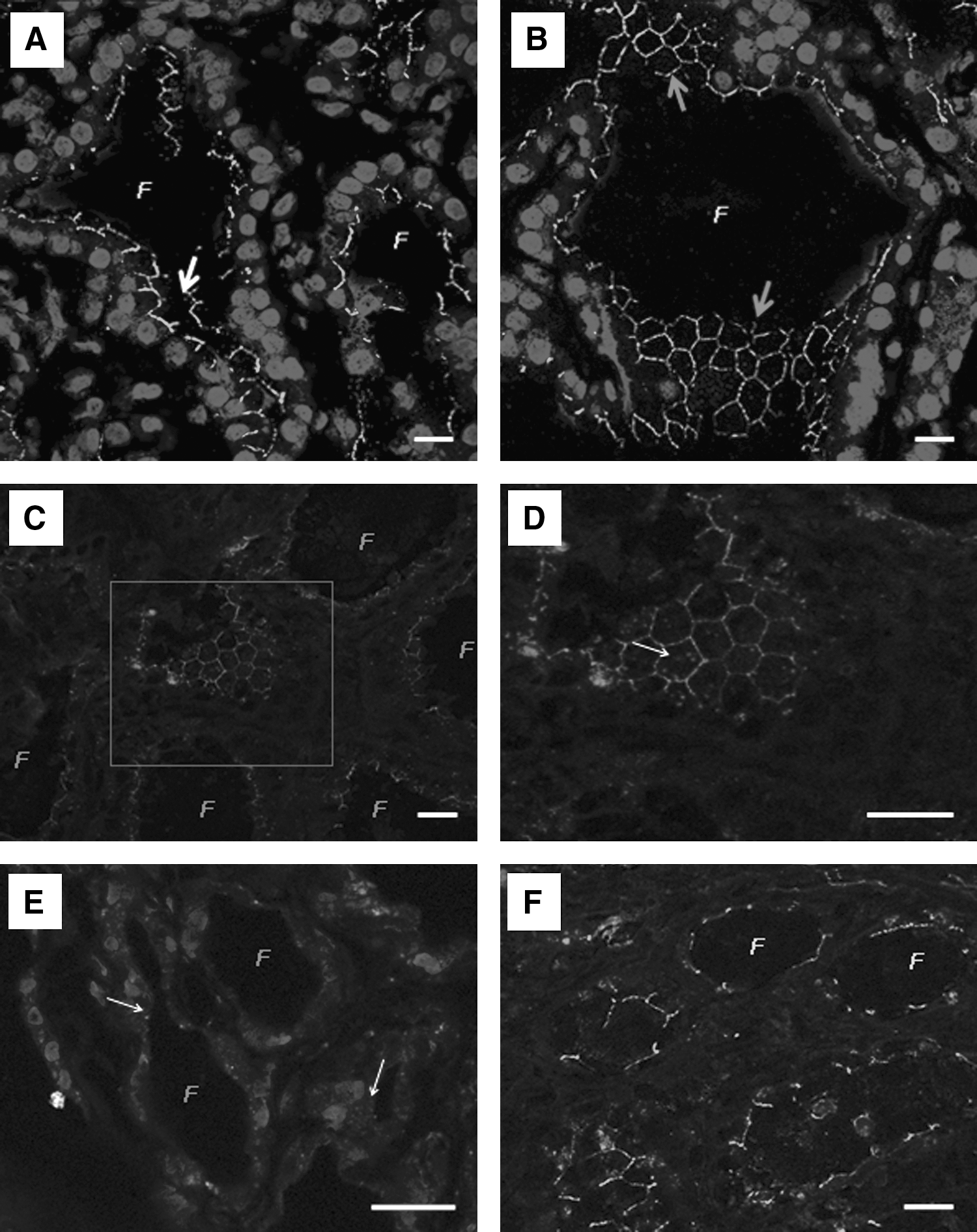
Expression and localization of Cx43 in normal thyroid, Graves' disease, and thyroiditis.
Depending on sections, thin segments of staining or alveolar pattern reflecting the edges of follicular cells were detected (Fig. 1A, B), leading us to conclude that Cx43 is mostly localized at the plasma membrane of follicular cells. We interpreted this homogeneous and regular staining as “normal,” and any other pattern (cytoplasmic or absent) seen in the pathologies was considered abnormal (Table 1).
The numbers of cases exhibiting each of the various patterns of Cx43 staining are indicated. These patterns were considered as “normal” when Cx43 is localized exclusively in the membrane (regular and irregular) and abnormal when Cx43 is aberrantly expressed in the cytoplasm or not detected. The percentage of samples exhibiting either “normal” Cx43 membrane staining or abnormal staining is indicated for every kind of pathology. The p-value has been obtained by comparing each pathology group (thyroiditis, Graves' disease, multinodular goiters, and adenomas) versus the papillary carcinoma group. There was no significant difference comparing thyroiditis or Graves' disease against adenomas.
Weak staining.
Percentages of staining and p-value for the different types of adenomas (follicular and oncocytic) are combined.
Neither percentage nor p-value is considered for follicular cancer, as only one case was diagnosed.
Cx43, connexin43.
Cx43 expression in Graves' disease and thyroiditis
By considering Graves' disease and thyroiditis samples separately (Table 1), it appeared that the proportion of regular Cx43 staining in the membrane was higher (66.7%) among the cases of Graves' disease (Table 1; Fig. 1C, D). Only two cases (16.7%) exhibited an irregular staining but still in the plasma membrane. Another two cases of Graves' disease (16.6%) showed either no or a cytoplasmic (Fig. 1E) staining pattern.
On the other hand, 50% of thyroiditis cases exhibited a normal staining at cell–cell contacts of the follicular epithelium (Fig. 1F). Such a staining was not so regular in the membrane of the other half of samples and presented cytoplasmic spots for some of them (25%). Interestingly, the normal staining of Cx43 corresponded to the samples of sub-acute thyroiditis, whereas less regular pattern of Cx43 characterized the samples of chronic lymphocytic thyroiditis.
Cx43 expression in multinodular goiters
Among the thyroid pathologies we studied, these 46 cases exhibited the highest heterogeneity of Cx43 expression/localization (Table 1). A normal membrane staining was observed in 20 cases (43.5%), whereas an irregular membrane staining affected 5 cases (10.9%). Nine cases (19.6%) were characterized by weak staining of Cx43 that was not clearly localized in plasma membrane or cytoplasm. More aberrant Cx43 cytoplasmic localization or no detection were observed in 2 cases (4.4%) and 10 cases (21.7%), respectively (Fig. 2A, B). Interestingly, this disparity of Cx43 staining patterns tends to support the heterogeneity of this disease.
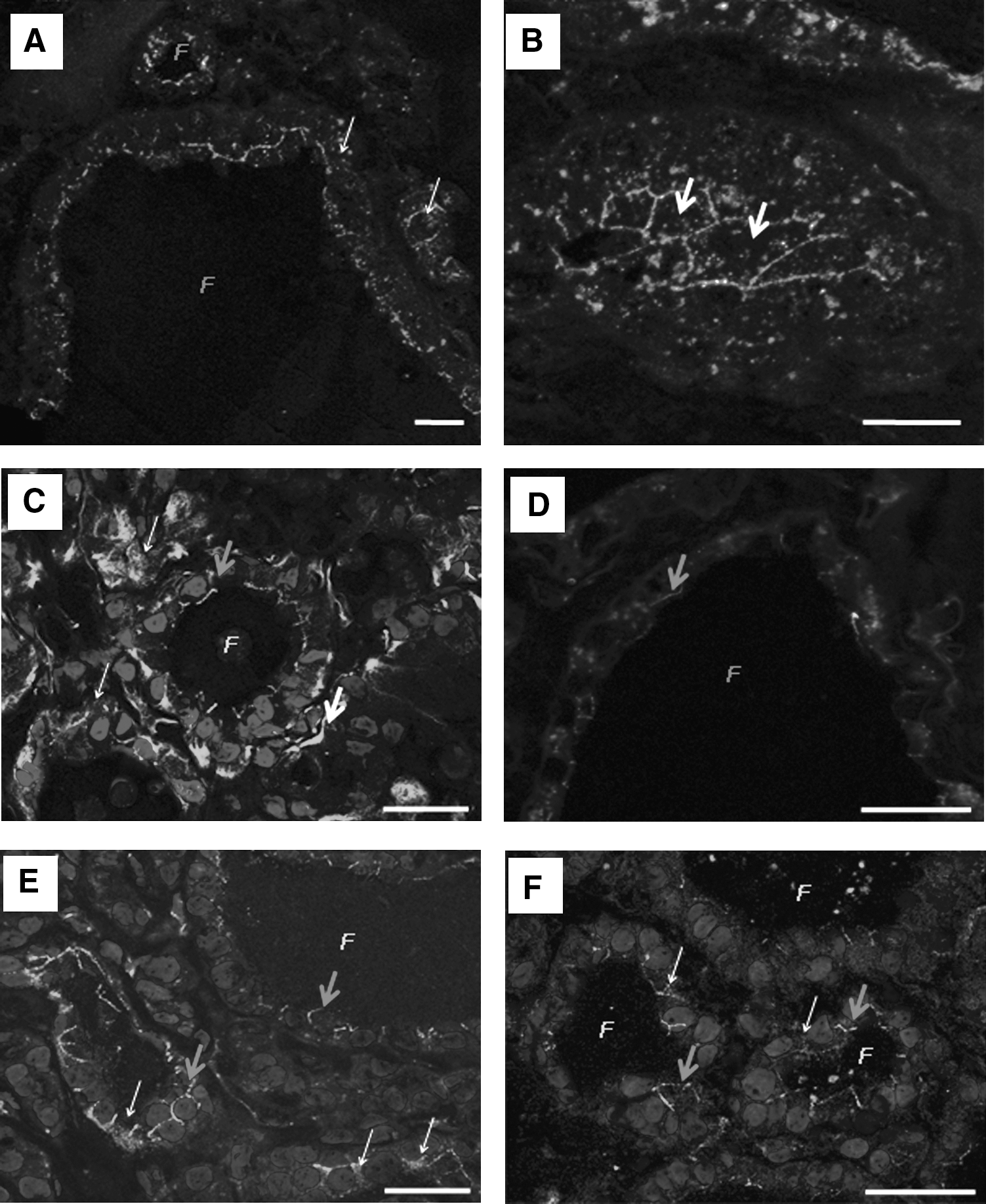
Examples of Cx43 detection in multinodular goiters and adenomas.
Cx43 expression in adenomas
Adenomas were the most frequent pathology after goiters (n = 38). They were diagnosed as oncocytic (n = 10) or follicular (n = 28) adenomas (Table 1). For oncocytic adenomas, the localization/expression of Cx43 was distributed as follows: three cases (30%) exhibited a regular membrane staining, three cases showed irregular membrane staining, two cases (20%) had cytoplasmic and membrane staining (Fig. 2C), and two cases showed no staining.
The same Cx43 localization/expression patterns were found in follicular adenomas but at different ratios: 10 cases (35.7%) exhibited a weak regular membrane staining, 4 cases (14.3%) had irregular membrane staining (Fig. 2D), 10 cases (35.7%) showed cytoplasmic and irregular membrane staining (Fig. 2E, F), and 4 adenomas (14.3%) showed no staining. Even if there is a high percentage of abnormal staining in this group, it is not specific for this pathology.
Cx43 expression in differentiated thyroid carcinomas
There was only one case of follicular carcinoma in our series, which presented absence of staining. The remaining cases were papillary thyroid cancers (n = 17), and most of them showed an abnormal staining: five cases (29.4%) had both cytoplasmic and membrane staining (Fig. 3A), seven cases (41.2%) exhibited no staining (Figs. 3B and 4), four cases (23.5%) had only cytoplasmic staining (Fig. 3C), and one case (5.9%) had normal but weak regular membrane staining (Fig. 3D).
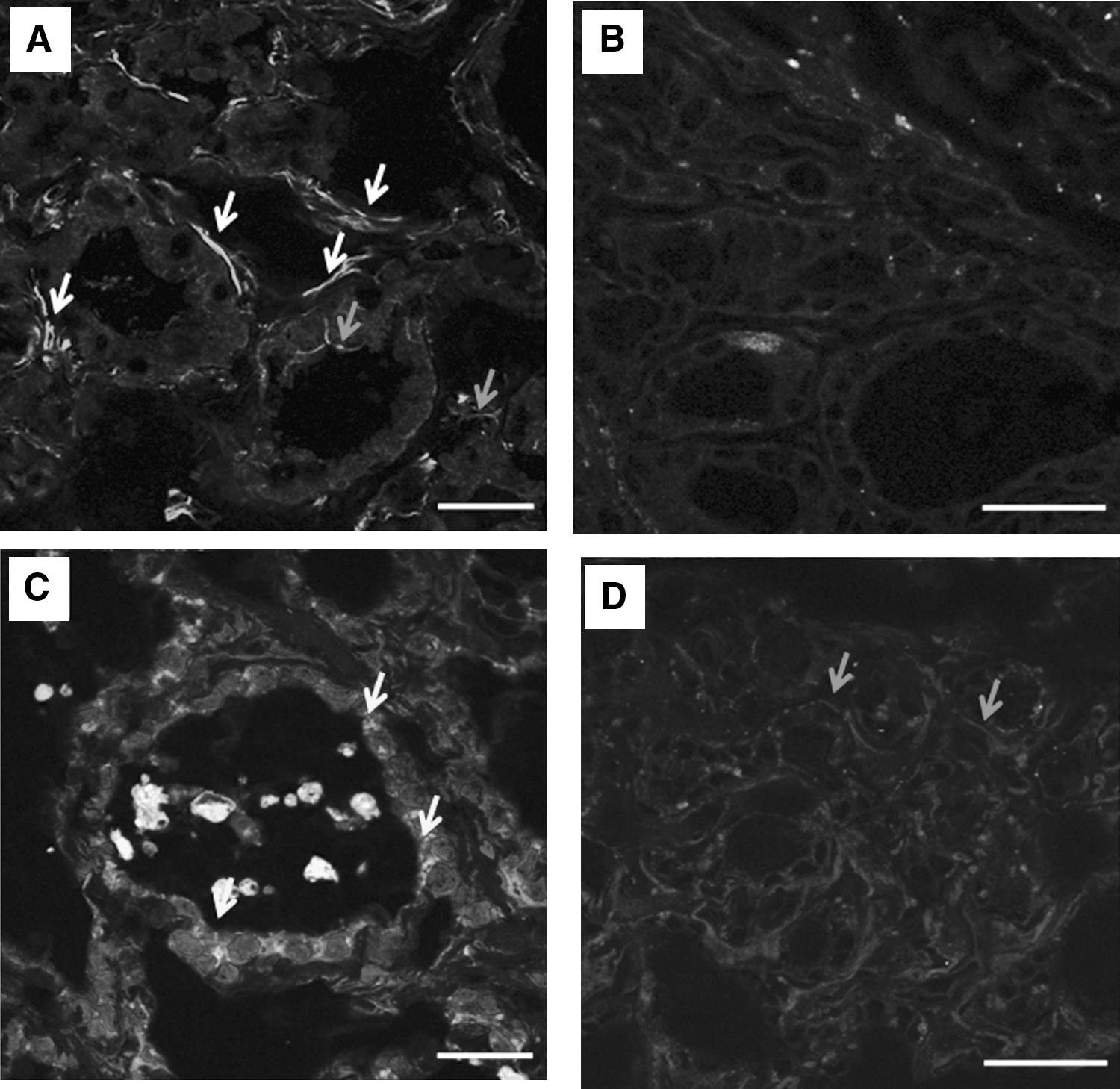
Cx43 detection in papillary carcinomas.
Cx43 detection in a papillary carcinoma.
Contrary to other thyroid pathologies we analyzed, the abnormal pattern of Cx43 expression is a constant finding in differentiated thyroid cancer (Table 1). When the comparison between normal and tumor tissues was possible, the aberrant expression of Cx43 was clearly associated with the cancer phenotype (Fig. 4).
Statistical analysis of the expression patterns
Based on the different Cx43 expression patterns we analyzed, normal (membrane) and abnormal patterns were regrouped to compare the expression of Cx43 among the pathologic groups and conduct statistical analysis.
These results show that a modified Cx43 localization/expression is predominant in cancer. Therefore, the percentage of altered expression (cytoplasmic or no detection) of each group of pathology was compared versus cancer, considering this pathology as the most important in terms of diagnosis discrimination and prognostic value for the patient. From statistical analysis, a p-value was obtained for the comparison of every kind of pathology versus the carcinoma group (Table 1).
Cx43 mRNA expression in papillary carcinomas
The Cx43 mRNA expression was estimated in 25 papillary carcinomas and compared with the average Cx43 mRNA expression in 16 normal parts of the tissue used as normal control. In normal tissue, Cx43 mRNA was amplified, and the mean expression level was estimated to be 1.86 (ranging from 0.7 to 3.175). This level was found to be only 0.67 (range from 0.15 to 1.88) in papillary carcinomas. We observed an overexpression of Cx43 mRNA in 8% of the tumors (2/25 cases) and a downregulation in 68% of them (17/25 cases). Cx43 mRNA expression was found to be normal in 6 cases (24%). According to the level of Cx43 mRNA expression in tumors, the samples were divided into Group 1 (low expression; n = 17), Group 2 (normal expression; n = 6), and Group 3 (overexpression; n = 2). These results conclude that the majority of thyroid papillary carcinomas (nearly 70% of cases) are characterized by a downregulation of Cx43 transcription.
In situ detection of Cx43 protein in samples tested for Cx43 mRNA
To verify the correlation between the level of Cx43 mRNA expression and the in situ expression at the protein level, the immunohistochemistry detection of Cx43 was performed in 17 cases distributed in each of the three different groups of patients just defined (See Table 2). To do so, nine cases of Group 1 (low level of Cx43 mRNA), six cases of Group 2 (normal level of Cx43 mRNA), and two cases of Group 3 (high level of Cx43 mRNA) were studied.
Whatever their level of Cx43 mRNA, all the tumor samples exhibited an aberrant localization of the protein. When downregulated at the mRNA level (Group 1), this localization was differently altered (decreased membrane staining, cytoplasmic localization, and no detection). Such a variety of localization was probably a consequence of the high number of samples (nine). When Cx43 mRNA is normally expressed (Group 2), all samples presented a decreased staining of the protein at the membrane; among them, two samples exhibited an additional cytoplasmic staining. When overexpressed, the protein was found in the cytoplasm (group 3).
Due to the availability of tissues, only 9 out 17 samples of Cx43 mRNA-downregulated samples could be analyzed for immunohistochemistry.
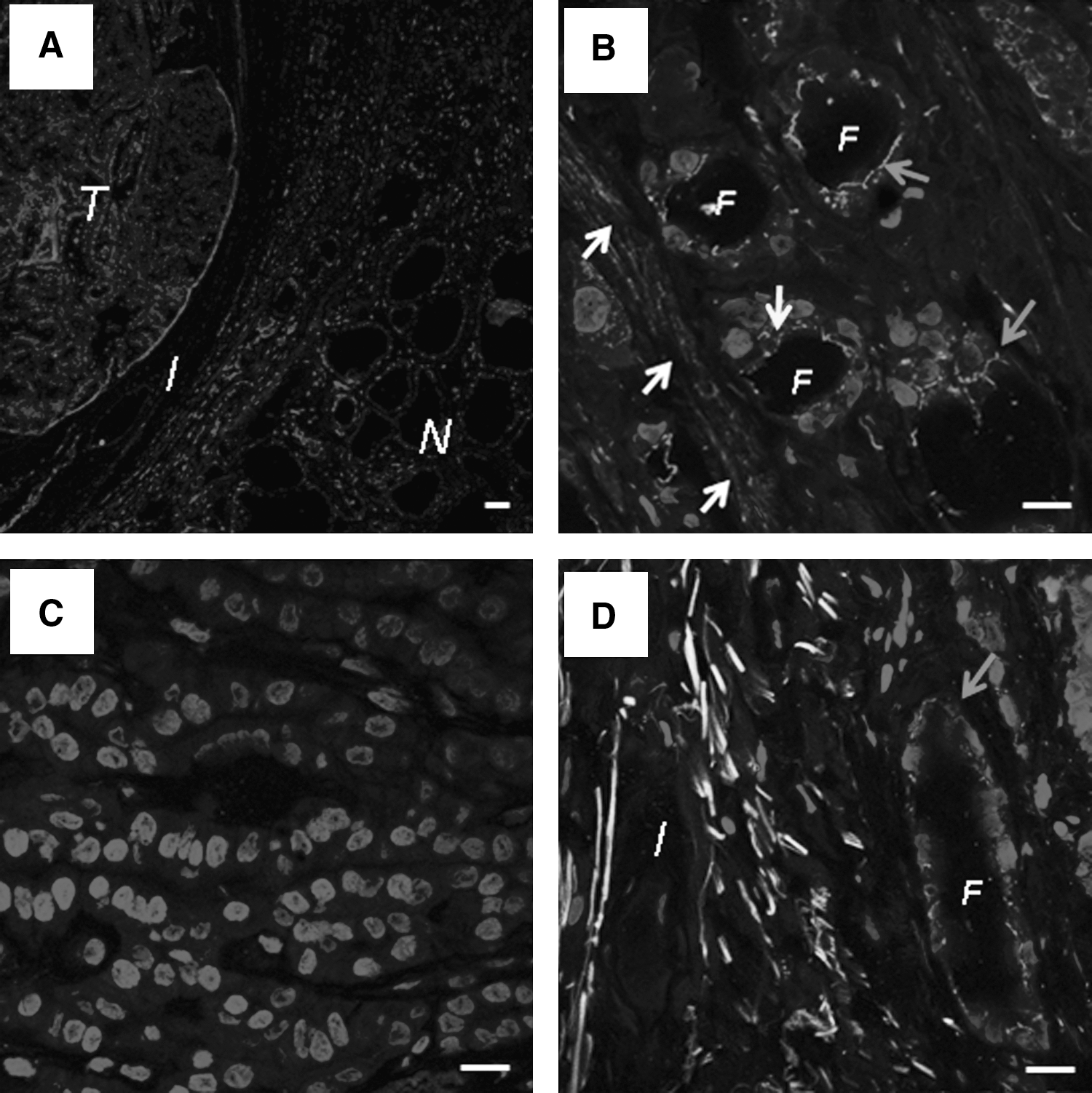
The cytoplasmic detection of Cx43 was associated with a decreased membrane staining.
First, in normal tissue, we observed that Cx43 was distributed homogeneously at the cell-cell contact areas of the follicular epithelium (Fig. 5). These results confirmed our observations made by immunofluorescence. In Group 1, the low transcriptional level of Cx43 was accompanied with either a complete lack of Cx43 detection in the plasma membrane (six of nine cases; Fig. 5A) or an irregular membrane staining (three of nine cases; Fig. 5B). It was interesting to note that the complete loss of membrane staining was associated, in four out of six cases, with an aberrant cytoplasmic distribution of Cx43. In Group 2, despite a normal transcriptional level, the Cx43 membrane staining was decreased in all six cases. In two of these cases, an aberrant cytoplasmic localization of Cx43 was observed (Fig. 5C). Finally, the two cases of Group 3, which exhibited an overexpression of Cx43 mRNA, were characterized by a cytoplasmic localization of Cx43 (Fig. 5D).

Cx43 detection in various papillary carcinomas by immunohistochemistry.
Discussion
In the thyroid, the abundance of gap junctions between follicular cells has been reported in several species (24 –28). Such an abundance and interspecies presence is in favor of an important role of gap junctions in thyroid. Some authors supposed that their presence during embryogenesis is necessary for the functional differentiation of the gland (25). Others have described that intercytoplasmic exchanges of second messengers mediated by gap junctions amplifies the hormonal response of the thyroid and regulates the secretion process of follicular cells (12,16,20). However, these data supporting that gap junctions and their structural proteins (Cx) are involved in the normal physiology of the thyroid were obtained from animal models. They only permit an indirect association of the presence of gap junctions with normal thyroid function in humans. Since a more direct association could be made by verifying whether the abundance and localization of the Cx is modified in human pathologies affecting thyroid function, we checked the in situ expression of Cx43 in various thyroid pathologies.
Our results show that only Cx43 is detected as gap-junctional plaques in nonpathological human thyroid by in situ immunofluorescence approach. After our analysis made on thyroid pathologies, we can conclude that different disturbed Cx43 patterns were observed. Some of them appeared as minor alterations such as weak and heterogeneous staining and could not be correlated to any particular pathology, and we suppose that even if such disturbances affect the efficiency of gap-junctional intercellular communication (GJIC), they probably do not completely impair it. However, this situation is drastically different when Cx43 is not detected or only present in the cytoplasm, as these patterns are not compatible with an efficient GJIC. Considering this situation, it is possible to distinguish three groups of thyroid pathologies that exhibit a clearly abnormal Cx43 behavior, with increasing prevalence starting from uncommon in inflammatory and autoimmune diseases (<25% of samples), common for goiters and adenomas (around 50%), to highly frequent for carcinomas (>90%).
Since the in situ approach revealed that Cx43 is commonly altered in papillary carcinomas, we checked whether such an alteration was observed at the transcriptional level. To do so, we compared the amount of Cx43 mRNA between tumor and normal tissues in a series of frozen papillary carcinomas. The study showed that the majority of those tumors (68%) were characterized by a lower amount of Cx43 mRNA. However, since some tumor samples did not exhibit any lower Cx43 mRNA amount than the normal part of the thyroid (32% of the samples) but even a higher expression (8%), we made a parallel in situ analysis at the protein level on the same series of papillary carcinomas. From this analysis, it appeared that the low Cx43 mRNA expression coincides with a lack (six of nine cases) or a decrease (three of nine cases) of Cx43 in the membrane; the equal expression is related to an irregular membrane staining with a cytoplasmic accumulation of Cx43 in some samples (two of six cases); the higher expression is associated with a cytoplasmic localization (two of two cases). It is interesting to note that the cytoplasmic detection of Cx43 protein was observed in all groups and then independently of the level of Cx43 mRNA. These observations confirm those we made by immunofluorescence approach. They indicate that the irregular or lack of the membrane localization of Cx43 is a common feature of the thyroid papillary carcinomas. Moreover, the comparison between the data we obtained in situ by immunohistochemistry and those quantifying the Cx43 mRNA permits to go further in the analysis, as it suggests that, at least, two different mechanisms may be involved in the irregular or lack of localization of Cx43 in the plasma membrane. A first one seems to involve a defective addressing of Cx43 to the membrane, as it accumulates in the cytoplasm of the tumor cells whatever the Cx43 mRNA level is even if this accumulation is clear for samples overexpressing Cx43 mRNA. The second mechanism that might be involved is a decreased transcription of the Cx43 gene leading to the decreased detection of Cx43 protein.
The first general conclusion from our work is that the lack of a functional localization of Cx43 (no detection or cytoplasmic localization) and the possible consequent GJIC deficiency are more related to pathologies of the thyroid which are characterized by an uncontrolled cell proliferation (goiter, adenoma, and carcinoma). This is in agreement with a previous study describing a lack of gap junctions in human oncocytic adenomas and carcinomas (29). This is also in agreement with experimental data showing Cx as regulators of cell differentiation and proliferation. In the thyroid, most of these experimental data were obtained from rat cells showing that the expression of the major rodent Cx in that tissue, Cx32, downregulates their proliferation (9). Such a situation has also been confirmed in vivo by using Cx32-transgenic mice (22). More than a growth regulator, Cx32 is involved in the differentiation of the thyrocytes. For instance, cell proliferation is increased, whereas thyroxine release is decreased in rat cells exhibiting a mutated and nonfunctional Cx32 (20). This balance between decreased GJIC, increased cell proliferation, and decreased differentiation of thyrocytes has indeed been observed in several experimental models (20,21) as well as the reverse situation when Cx32 is expressed (9,12). Interestingly, these controls on cell differentiation and cell proliferation were specifically induced by Cx32 acting at the gene expression level (increased expression of p21 and p27) and not by Cx43 in the models used (17,30,31). This is probably different in humans, as, contrary to rodents, Cx43 is the only Cx detected in situ in the normal human thyroid at the protein level. According to our results, Cx43 might play in the human situation the role played by Cx32 in rodents even if the role of Cx32 appeared to be recently more complex than what was expected (32). This assumption is simply based on our descriptive data and has to be confirmed by accurate cell models.
According to our observations, the altered Cx43 expression is commonly related to differentiated thyroid cancer phenotype. These findings contrast with the heterogeneity of expression found in goiters and adenomas and suggest that these adenomas might evolve eventually to carcinomas consequently to the absence of optimal cell communication. If this were true, Cx43 aberrations might be a prerequisite for malignant evolution of cells, therefore having potential clinical consequences.
Our results suggest that aberrations of Cx43 pattern of expression are commonly associated with papillary thyroid carcinoma. Since such alterations are also observed in about half of the benign lesions exhibiting cell growth dysfunction (multinodular goiter and follicular/oncocytic adenomas), our data show that the frequency of abnormal Cx43 staining increases from benign to malignant lesions and might suggest it is an early event in the tumor development of thyroid. Interestingly, such a loss of Cx43 function is probably the consequence of various molecular mechanisms leading to either a decrease of Cx43 mRNA amount or a cytoplasmic delocalization of the protein. Further investigations are required to determine the molecular and cellular consequences of such Cx43 aberrations of expression in thyroid cells. In particular, it will be interesting to study whether such aberrations affecting Cx43 influence the expression of genes involved in cell proliferation such as p21 and p27. Moreover, it is possible that the involvement of Cx in thyroid cancer is more complex, as the expression of another type of the Cx family, Cx26, has been observed in association with large tumor size, lymph node metastases, and presence of intraglandular dissemination (33).
Footnotes
Acknowledgments
This work was financially supported by “La ligue contre le cancer” (Comités de Charente, Charente Maritime et Vienne; France) and “Cancéropôle Grand Ouest.” Confocal microscopy was done at the Confocal Microscopy facility of UMR-CNRS 6187.
Disclosure Statement
The authors declare that no competing financial interests exist.