
Abstract
Background:
Rat and mouse have been widely used to estimate the radiation risk and tumorigenic effects of radiation with extrapolating the findings to humans. RET/PTC is a characteristic genetic alteration frequently found in radiation-induced thyroid cancer in human populations. Recently, nuclear architecture and spatial proximity between recombinogenic genes have been implicated as important factors in the generation of RET/PTC and other chromosomal rearrangements in human cells. However, it is unknown whether the nuclear architecture in rodent thyroid cells is similar to that of human thyroid cells. The aim of this study was to test whether the proximity effects that are observed between loci involved in RET/PTC rearrangements in humans are conserved across different species.
Methods:
Using 3D fixation, fluorescence in situ hybridization, and confocal microscopy, we compared the distance between genes involved in RET/PTC rearrangement in normal thyroid cells from humans, mice, and rats.
Results:
While in humans, RET, NCOA4, and H4 are all located on the same chromosome (10q), in rodents these genes are located on separate chromosomes. In mouse, RET is located on chromosome 6F1, NCOA4 on 14B, and H4 on 10B5.3. In rat, RET is on chromosome 4q42, NCOA4 on 16p16, and H4 (TST1) on 9q36. We further observed that in human thyroid cells, mean distance between genes involved in two most common types of RET/PTC, that is, RET and NCOA4 (partners of RET/PTC3) and RET and H4 (partners of RET/PTC1), was 1.08±0.04 and 1.24±0.05 μm, respectively. In mouse thyroid cells, these distances were 3.21±0.1 and 3.43±0.1 μm, and in rat cells the values were 3.37±0.1 and 3.87±0.1 μm (p<0.001). Moreover, we found that in contrast to human thyroid cells, in rodent cells these genes were randomly positioned with respect to each other.
Conclusions:
The differences in nuclear architecture and spatial positioning of genes involved in RET/PTC rearrangements between human and rodent thyroid cells raise a concern about suitability of animal models for assessing RET/PTC-driven thyroid carcinogenesis in humans.
Introduction
The pathogenesis of papillary thyroid carcinoma involves the activation of the mitogen-activated protein kinase (MAPK) signaling pathway. This activation can typically occur either as a result of chromosomal rearrangements involving the RET gene (14,15) or due to point mutations involving the BRAF or RAS genes (16 –20). Although rearrangements of the RET gene, known as RET/PTC, are found in ∼20% of adult sporadic papillary carcinomas, they are consistently found with a significantly higher frequency (50%–80%) in papillary carcinomas from patients with the history of radiation exposure (21 –24). In addition to RET/PTC, chromosomal rearrangements involving the NTRK1 gene, known as TRK rearrangements (25,26), and BRAF/AKAP9 rearrangement that activates the BRAF gene (27) have been identified in radiation-induced thyroid papillary carcinomas. In contrast, BRAF and RAS point mutations are found with a high prevalence in sporadic but not in radiation-induced papillary carcinomas (28 –30). Moreover, among atomic bomb survivors in Japan, the presence of RET/PTC in papillary carcinomas had direct correlation with the dose of radiation, whereas the inverse correlation was found between the dose to the thyroid and BRAF point mutations (31,32). The link between radiation exposure and chromosomal rearrangements, particularly RET/PTC, has been further confirmed in the experimental setting. Exposure of cultured cells and human fetal thyroid tissue xenografts in SCID mice to ionizing radiation have resulted in the generation of RET/PTC rearrangement, and in some studies the generation of RET/PTC was dose dependent (33 –36). All these findings indicate that gene activation by chromosomal rearrangement rather than via point mutation is a characteristic genetic signature of thyroid radiation carcinogenesis.
Estimations of radiation risk in humans have been largely based on information obtained from atomic bomb survivors and patients treated with radiotherapy, all of whom were exposed to high doses of radiation. In contrast, the estimation of risk associated with low-dose radiation has been predominantly based on animal models. Experimental data from animals have been the basis for dose recommendations, dose–rate effectiveness factors, and radiation quality factors. Several studies have been published that extrapolate data on mortality and life shortening from animals to humans (37 –40). As far as specific tissue responses are concerned, carcinogenic effects of radiation exposure have been observed in mice and rats, and based on these studies extrapolation of animal data to humans has been extended also to the tumorigenic effect of radiation (41 –46). The thyroid carcinogenic effects of high-dose X-ray exposures (≥10 Gy) were observed in mice and rats (41 –45), and one study utilized lower doses of radiation (0.94–10.6 Gy) to induce thyroid carcinomas in rats (46). Surprisingly, whereas in humans exposure to radiation leads predominantly to papillary carcinomas, none of the radiation-induced thyroid tumors in animals were of the papillary phenotype. One of the explanations for such a discrepancy was that rodents develop only follicular-pattern thyroid tumors and those could be considered as an equivalent of papillary carcinoma in humans. However, more recently, it was shown that transgenic mice expressing RET/PTC and other oncogenes actually did develop thyroid tumors with papillary architecture and nuclear features similar to papillary carcinomas in humans (47). Therefore, another explanation is required for the lack of papillary carcinoma phenotype in rodent models of radiation-induced thyroid cancer.
Recently, nuclear architecture and spatial proximity between recombinogenic genes have been implicated as important factors in the generation of chromosomal rearrangements in human cells. Studies on gene positioning in interphase nuclei of the thyroid and of other cell types have firmly established the presence of close spatial positioning of genes involved in various types of chromosomal rearrangements. In thyroid cells, this has been shown for genes involved in two most common types of RET/PTC, that is, RET/PTC1 (RET and H4 genes) (48) and RET/PTC3 (RET and NCOA4) (49) as well as for the genes involved in TRK rearrangement (50). Spatial proximity has also been demonstrated between the TMPRSS2 and ERG genes, which frequently rearranges in prostate cancer (51,52), as well as between several gene pairs involved in recurrent rearrangements in hematopoietic malignancies (53,54).
The rationale for using animal models to simulate carcinogenic effect in humans presumes that animals and humans have similar tissue morphology and physiology and share common mechanisms for DNA repair, cell proliferation and differentiation, and gene expression (55,56). However, no data are available to date that establish whether nuclear architecture of rodent cells is similar to that of human cells, particularly with respect to spatial proximity between recombinogenic chromosomal loci involved in carcinogenic rearrangements. In this study, we analyzed the distances between genes involved in the most common types of RET/PTC rearrangement, that is, RET, NCOA4, and H4, in normal thyroid cells from humans, mice, and rats to test whether the proximity effects between the potentially recombinogenic genes exist across species.
Materials and Methods
Human, mouse, and rat thyroid tissues
Normal thyroid tissues from adult individuals were harvested fresh immediately after surgery for benign thyroid disease using the University of Pittsburgh Institutional Review Board approval. Normal rodent thyroid tissues were obtained from C57Blk6 mice and Sprague-Dawley rats. Tissues were obtained from animals sacrificed for conditions not related to normal thyroid structure and function. Histological examination of the removed tissues was performed in frozen sections stained with hematoxylin and eosin to confirm that they were normal thyroid tissues (Fig. 1)
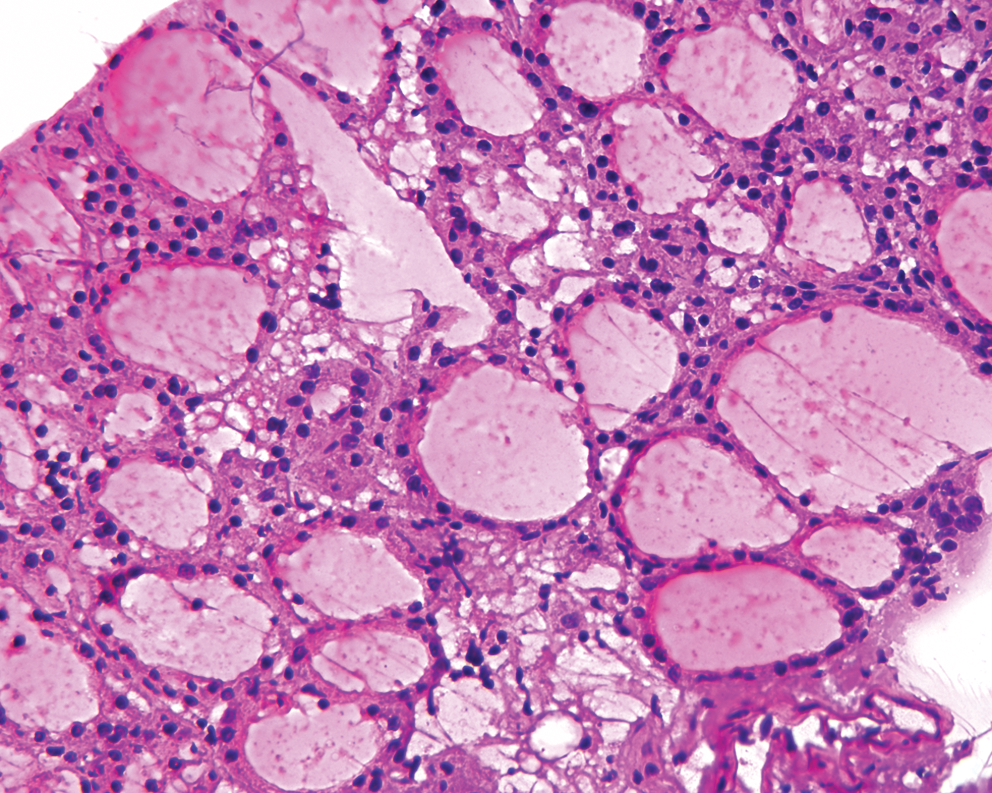
Frozen section image showing normal histology of the mouse thyroid gland used to obtain thyroid cells for the study. Color images available online at
Preparation of nuclei for 3D fluorescence in situ hybridization
Touch preparations of thyroid tissues from human, mouse, and rat were fixed in 3.7% paraformaldehyde (PFA) with repeated freeze–thaw cycles in liquid nitrogen to preserve their 3D nuclear architecture (57,58). Briefly, touch preparation on positively charged slides were fixed in 3.7% PFA followed by treatment with 0.1% Triton X-100 and incubation in 20% glycerol. Slides were subjected to four rounds of freeze–thaw treatment in liquid nitrogen followed by washes in phosphate-buffered saline (PBS), 0.1 M HCl, and 2×saline-sodium citrate buffer (SSC). Slides were stored in 50% formamide/2×SSC at 4°C until use. Care was taken to prevent slide drying at any point in the preparation process.
DNA probes and fluorescence in situ hybridization
BAC clones RP11-351D16 (Hs RET), RP11-481A12 (Hs NCOA4), RP11-435G3 and RP11-369L1 (Hs H4), RP24-303J2 (Mm RET), RP23-469G7 (Mm NCOA4), RP23-223L12 (Mm H4), RP23-56I20 (Mm p53), CH230-264F14 (Rn RET), CH230-46G21 (Rn NCOA4), CH230-510G6 (Rn H4 [TST1]), and RP32-558D10 (Rn p53) were obtained from BAC/PAC Resources, Children's Hospital, Oakland. BAC clone RP11-481A12 containing the human NCOA4 gene was subcloned into fosmid vector after cutting with restriction enzymes (Epicentre). A mixture of subcloned probes (SC10, SC19) containing 70 kb of the NCOA4 gene and its flanking regions was used as a probe for NCOA4. The probes were labeled by nick translation using Spectrum Green-dUTP, Spectrum Orange-dUTP, or Spectrum Red-dUTP (Abbott). Stored slides were treated with Collagenase H, washed in PBS, and postfixed in 10% formalin. Fluorescently labeled probe was applied and the slides were hybridized overnight at 37°C. Posthybridization washes were carried out in 50% formamide/2×SSC and 2×SSC and slides were counterstained with DAPI.
Confocal microscopy, 3D reconstruction, and distance measurements
Microscopy was performed using a Leica SP5 TCS 4D confocal laser scanning fluorescence microscope. On average, 50 nuclei from two different donors were scanned for humans, mouse, and rat and the digital images were reconstructed using Volocity software (Perkin Elmer). Hybridization signals were subjected to uniform thresholding to demarcate the loci signals. Based on the thresholding, the centroid of the probe signal was automatically calculated and represented as x, y, and z coordinates. The 3D distance between probe pairs were mathematically calculated. Suppose that (xi
, yi
, zi
) are coordinates for the ith locus in an arbitrary coordinate system, i=1, 2, 3, 4, 5. The physical distance between any two loci i and j is then:
In humans, since RET, NCOA4, and H4 are located on the same chromosome, appropriate grouping of signals for each chromosome was readily possible. However, since in mouse and rat these genes are located on separate chromosomes, the closest distance between the two possible distance combinations for each signal pair was considered for analysis.
Statistical analysis
Comparison of distances between probe pairs was performed using the two-tailed Student's t-test for means after confirming normality of the data. The difference between two values was considered significant when the p-value was less than 0.05. Cumulative frequency distributions function (CDF) of absolute distances for specific probe pairs were built by plotting the values on the X-axis and on the Y-axis the proportions of all data points that were smaller than this measurement. The CDFs for the different genes were compared by the two-sample Kolmogorov–Smirnov test, which is a nonparametric test for continuous data and is used to determine whether two distributions are similar or not (59).
Results
Unique spatial positioning of RET, NCOA4, and H4 in human thyroid cell nuclei when compared with rat and mouse thyrocytes
Since data from rat and mouse models of thyroid radiation tumorigenesis have been transposed to humans, we first analyzed the distances between the genes involved in RET/PTC1 and RET/PTC3 rearrangements in normal human, rat, and mouse thyroid cells. In humans, the RET, NCOA4, and H4 genes are all located on chromosome 10. However, in mouse and rat cells, these genes are positioned on different chromosomes. In mouse, RET is located on chromosome 6F1, NCOA4 on 14B, and H4 on 10B5.3. In rat, RET is on chromosome 4q42, NCOA4 on 16p16, and H4 (TST1) on 9q36. We measured 3D distance between these three recombinogenic genes in thyroid cell nuclei from each of these species hybridized with probes for RET, NCOA4, and H4 (Fig. 2). In normal human thyroid nuclei, the mean distances between RET-NCOA4 and RET-H4 were 1.08±0.04 and 1.24±0.05 μm, respectively. In mouse cells, the values for the distances were 3.21±0.1 and 3.43±0.1 μm, respectively. In rat cells, these values were 3.37±0.1 and 3.87±0.1 μm, respectively. The mean distances for both probe pairs in human cells were significant different when compared with mouse and rat cells (p<0.001).
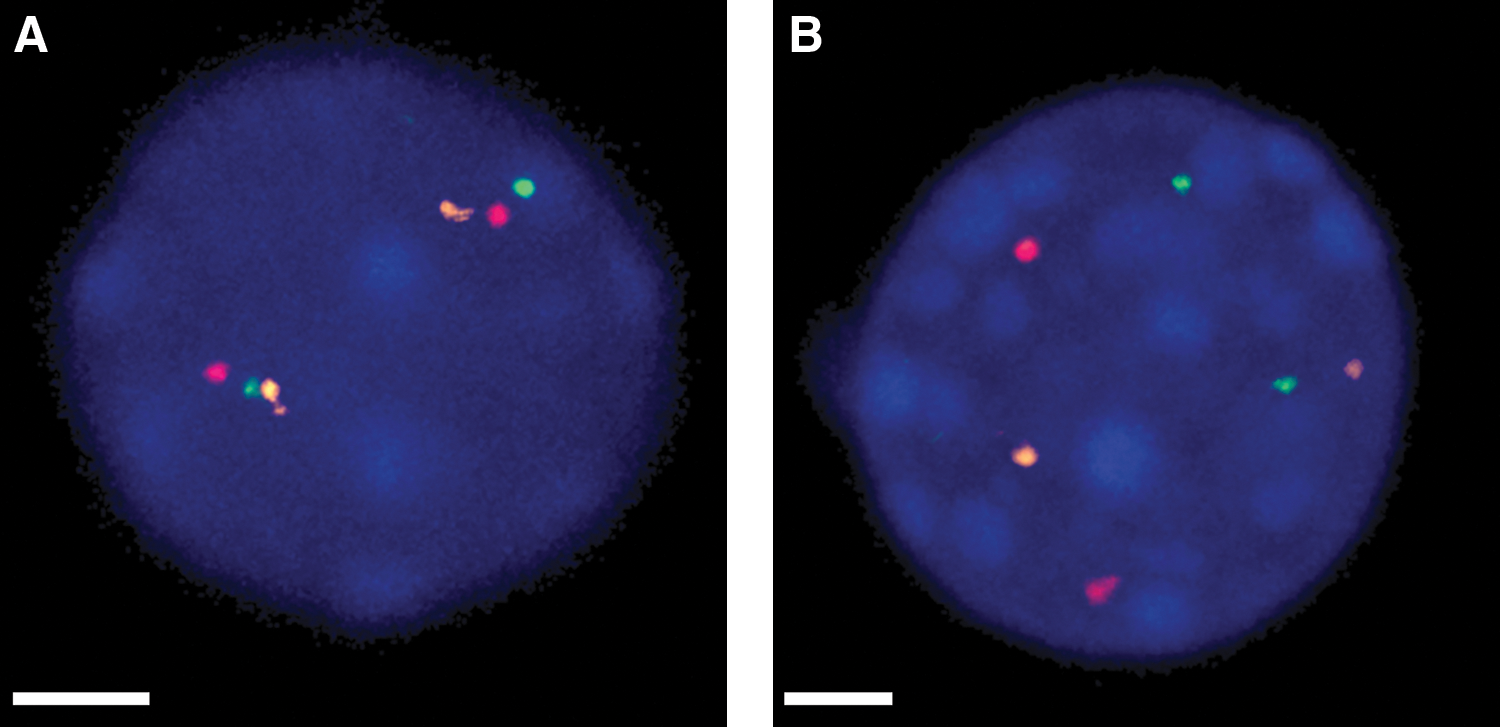
Extended focus (2D) view of normal thyroid cell nuclei showing location of the RET (green), NCOA4 (red), and H4 (orange) probes.
Although there is strong evidence for the requirement for close spatial positioning between chromosomal loci participating in recombination, exact limits of the distance that facilitates gene fusion have not been established. Therefore, we then analyzed the differences in the entire distribution of distances between recombinogenic loci in these species. This was achieved using the CDF curves, which are built taking into account the full spectrum of distances between each probe pair. The CDF curves show that, in human thyroid nuclei, RET-NCOA4 and RET-H4 distances were significantly shorter over the entire distribution of values when compared with mouse and rat cells (p<0.05; Fig. 3). The respective CDFs in cells from the two rodents were similar (p>0.05).
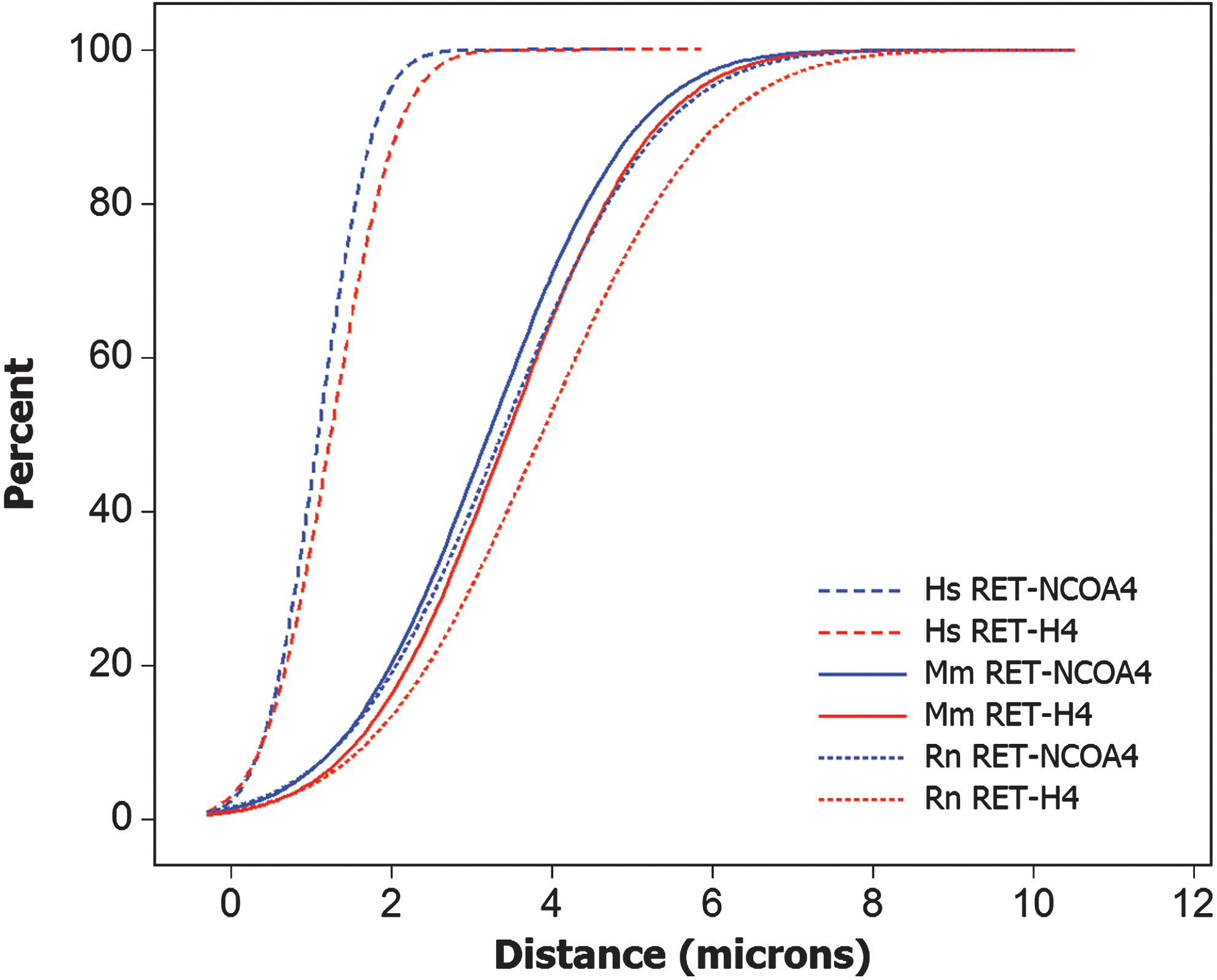
Cumulative frequency distribution function (CDF) of RET-NCOA4 and RET-H4 distances in human (Hs), mouse (Mm), and rat (Rn) thyroid cells showing significant difference in the distributions between human and rodent cells. Color images available online at
Random spatial positioning of RET/PTC partner genes in rodent cells
Once we had established that in rodents, the RET, NCOA4, and H4 gene loci were positioned on spatial separation much larger when compared with human thyroid cells, we proceeded to examine whether these gene loci were at least nonrandomly located with respect to each other, that is, were positioned on a distance shorter that the distance between nonrecombinogenic genes. In human cells, RET/PTC partner genes have been found to be located not only on small spatial separation, but also nonrandomly with respect to each other (48,49). To address this possibility in the rodent cells, we used four-color fluorescence in situ hybridization, adding a probe for the p53 gene to the probes for RET, NCOA4, or H4. The p53 gene is not known to be involved in recombination with either RET, NCOA4, or H4. Distances between the recombinogenic pairs, RET-NCOA4 and RET-H4, were compared with distances between the nonrecombinogenic probe pairs, NCOA4-H4, RET-p53, NCOA4-p53, and H4-p53, in both types of rodent cells (Table 1). The mean distances between recombinogenic gene pairs and nonrecombinogenic probe pairs were not significantly different in both types of rodent cells (p>0.05). Analysis of CDF curve plots also revealed no statistical difference for mouse (Fig. 4A) and rat (Fig. 4B) (p>0.05). These results indicate that in the interphase nuclei of rodent cells, the RET, NCOA4, and H4 gene loci are positioned randomly and on a large spatial separation with respect to each other.
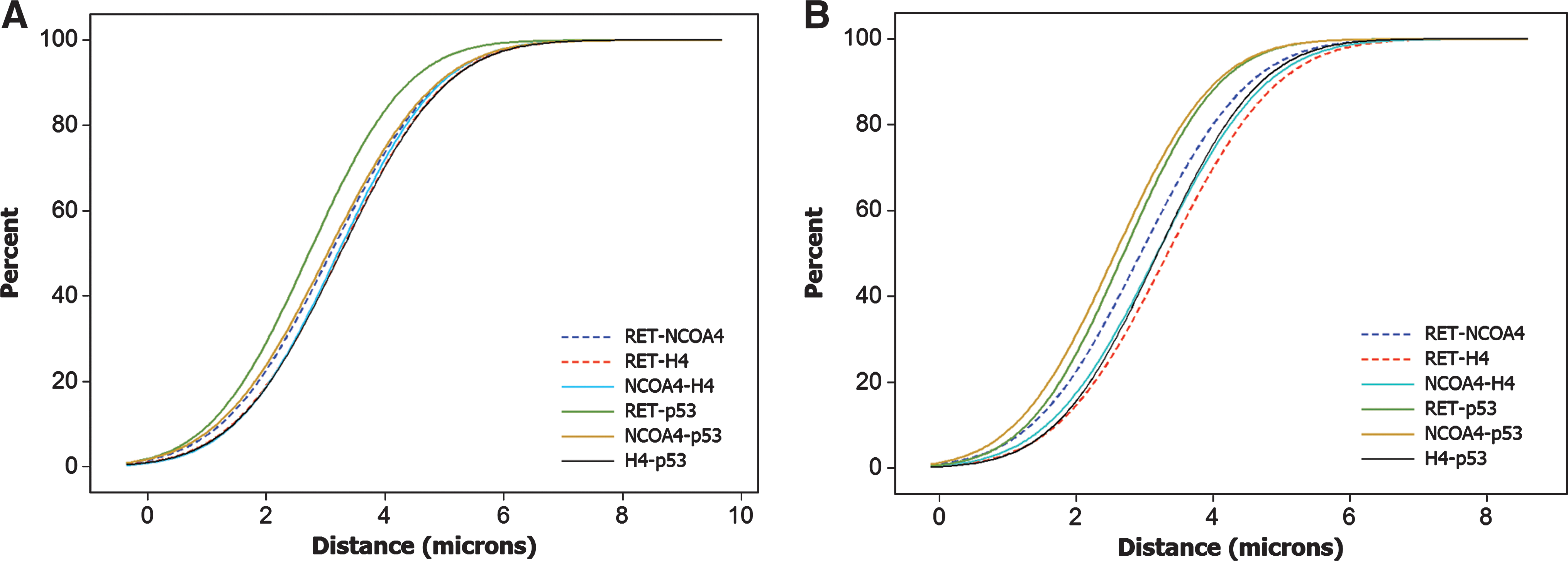
CDFs of the six different combinatorial distances measured after four-color fluorescence in situ hybridization (RET, NCOA4, H4, and p53) showing the comparison of the distances between recombinogenic (dashed lines) and nonrecombinogenic (solid lines) probe pairs in mouse
Shading indicates recombinogenic probe pairs. Standard error of the mean for all measurements ranged between 0.08 and 0.1 μm.
Discussion
Animal model systems have always been a vital research tool to simulate various human diseases and the effects of environmental factors on humans. This includes modeling of radiation carcinogenesis and radiation risk assessment in addition to survival and mortality. The cross-species conservation of genes involved in cell proliferation and DNA repair provide a reasonable justification for such usage. However, the appropriateness of animals to model radiation-induced carcinogenesis in humans remains questionable. Since in most cases the initiation of radiation-induced carcinogenesis requires chromosome recombination, the role of nuclear architecture and gene positioning within the nucleus has to be considered as an important prerequisite.
The results of this study indicate that the genes contributing to the RET/PTC fusion are in fact positioned in a different fashion in interphase nuclei of rat and mouse thyroid cells when compared with human cells. Specifically, in rodent cells these genes are (i) located on different chromosomes, (ii) reside on a much larger spatial separation, and (iii) located randomly with respect to each other. Since proximity between recombinogenic loci appears to play an important role in chromosomal rearrangements, these results offer an explanation for the absence of murine orthologs of human RET/PTC and also the inability to establish a radiation-induced RET/PTC model in rodents. These findings raise concerns about the appropriateness of using animals to model RET/PTC-related thyroid carcinomas, and therefore, radiation-induced carcinogenesis.
There is a general belief that animals can be used to model radiation effects and extrapolation of the experimental findings on humans should provide reasonably helpful information. Although this may be the case for noncancer effects of ionizing radiation, the animal models may have more problems as far as modeling radiation carcinogenesis is concerned. For instance, although susceptibility to radiogenic induction of tumors is seen in rats and, to lesser extent, in mice, it was obvious that the phenotype of tumors was different than in humans, that is, papillary carcinoma in humans and follicular carcinoma in rodents (60,61). Contrary to radiation-induced tumors in humans, where RET/PTC rearrangements are highly common and RAS mutations are exceedingly rare, in rats radiation-induced tumors frequently harbor KRAS mutations (62,63). The results of this study suggest that the differences in nuclear architecture and chromosome positioning in thyroid cells of these rodents does not provide a structural basis for generation of RET/PTC rearrangements. This would explain the lack of the papillary carcinoma phenotype in animals exposed to ionizing radiation. It also suggests that although rats and mice may be suitable to model thyroid follicular carcinoma, a tumor that frequently develops via RAS mutation and has also been found to have an increased incidence after radiation exposure (64), the rodent models are likely to severely underestimate the risk of papillary thyroid carcinoma induced by ionizing radiation.
Whether or not this limitation is applicable only to thyroid cancer or may also affect other types of radiogenic cancers remains unknown. Another type of such malignancy is leukemia, which also frequently harbors chromosomal rearrangements. For instance, BCR and ABL genes, despite being on separate chromosomes, have been shown to be positioned in close proximity to each other in human lymphocytes (54).
Overall, the results of this study provide additional evidence for the role of spatial proximity of the potential rearrangement sites in the generation of gene fusion in cancer, rather than supporting another possibility, that is, generation of chromosomal rearrangement as a result of selection for rare, random events that possess strong transformation potential.
In summary, the results of this study raise a significant concern about suitability of animal models to assess RET/PTC generation and other aspects of radiation-induced thyroid cancer in humans because of substantial differences in nuclear architecture between the species.
Footnotes
Acknowledgments
This work was supported by the NIH grant CA88041. The authors thank Dr. Michael Shurin (Department of Pathology, University of Pittsburgh) for providing mouse thyroid tissues and Dr. Naoki Yoshimura (Department of Urology, University of Pittsburgh) for providing rat thyroid tissues.
Disclosure Statement
The authors do not have any commercial association that might create conflict of interest with the submitted manuscript and there are no competing financial interests to disclose.