
Abstract
Background:
MicroRNAs (miRNAs) are involved in the pathogenesis of human cancers, including medullary thyroid carcinoma (MTC). The aim of this study was to test the hypothesis that different miRNA profiles are related to RET status and prognosis in patients with hereditary MTC (hMTC) and sporadic MTC (sMTC).
Methods:
We analyzed the expression of nine miRNAs (miR-21, miR-127, miR-154, miR-224, miR-323, miR-370, miR-9*, miR-183, and miR-375) by quantitative real-time–polymerase chain reaction in 34 cases of sMTC, 6 cases of hMTC, and 2 cases of C-cell hyperplasia (CCH). We also analyzed the immunohistochemical expression of PDCD4, an miR-21 gene target. sMTC (n=34) was genotyped for somatic RET and RAS mutations. Disease status was defined on the basis of the concentration of serum calcitonin at the latest follow-up and other parameters as indicated in the results.
Results:
MTC and CCH were both characterized by a significant overexpression of the whole set of miRNAs (the increase being 4.2-fold for miR-21, 6.7-fold for miR-127, 8.8-fold for miR-154, 6.6-fold for miR-224, 5.8-fold for miR-323, 6.1-fold for miR-370, 13-fold for miR-9*, 6.7-fold for miR-183, and 10.1 for miR-375, p<0.0001). PDCD4 expression was significantly downregulated in MTC samples, consistent with miR-21 upregulation. Significantly lower miR-127 levels were observed in sMTC carrying somatic RET mutations in comparison to sMTC carrying a wild-type RET. In sMTC and familial MTC, the miR-224 upregulation correlated with the absence of node metastases, lower stages at diagnosis, and with biochemical cure during follow-up.
Conclusions:
miRNAs are significantly dysregulated in MTC, and this dysregulation is probably an early event in C-cell carcinogenesis. miR-224 upregulation could represent a prognostic biomarker associated with a better outcome in MTC patients.
Introduction
Age at diagnosis and stage of disease at the time of initial treatment currently represent the main prognostic indicators for MTC (2,3). The tumor's variable behavior in some cases strongly suggests, however, that cancer progression and survival may also be influenced by other molecular and pathological features (4,5). Among others, distinctive germinal RET mutations in the inherited forms and somatic RET mutations in sporadic cases represent the most important molecular markers for an adequate prognostic stratification of patients with MTC (6,7). In fact, RET status significantly influences (i) the mean age of tumor onset and the tumor's aggressiveness in familial cases, and (ii) the probability of achieving a biochemical cure and overall patient survival in sporadic cases (8 –10).
MicroRNAs (miRNAs) are short, single-stranded, noncoding RNAs that, by binding to the 3′-untranslated region of target messenger RNAs (mRNAs), are able to posttranscriptionally modulate gene expression, playing a crucial part in various biological processes (11,12). Dysregulation in miRNA expression has recently been implicated in the pathogenesis of various types of human cancers, including thyroid tumors (12 –14). However, the biological role of miRNA dysregulation in thyroid carcinogenesis remains poorly understood, since involved miRNA gene targets have been only partially recognized (15). Systematic miRNA evaluation in thyroid cancers (always supported by advanced histology and the clinical characterization of the specimens) will not only add to our knowledge of their basic biology, but could also pave the way to risk-stratified patient management programs and innovative therapies (12,13).
In this regard, there are only about two studies that evaluated miRNA expression in MTC. Nikiforova et al. investigated miRNA expression profiling in two fine-needle aspiration biopsy (FNAB) samples from MTC, finding a particular cluster of upregulated miRNAs (16). More recently, Abraham et al. studied a series of patients with MTC using miRNA microarray analysis and reported a signature of three miRNAs (miR-183, miR-375, and miR-9*) capable of distinguishing hMTC and sMTC cases (17).
Expanding the available information, the aims of the present study were to confirm miRNA dysregulation in a large series of patients with hMTC and sMTC, to determine whether there is a relationship between miRNA profiles and the outcome in MTC, and to determine whether there is a relationship between miRNA signatures and RET status in sMTC.
Patients and Methods
Patients
A consecutive series of 34 patients with sMTC (21 men and 13 women, median age 60, range 22–81 years) and of 6 patients with hMTC (3 men and 3 women, median age 40, range 32–59 years) collected from 2006 to 2009 with a median follow-up of 48 months (range 12–226 months) were included in the study (Table 1). Two germinal RET mutation carriers (2 men), who underwent prophylactic thyroidectomy with a final histology of C-cell hyperplasia (CCH), were also considered; the scattered foci of hyperplastic C-cells were macro-dissected and the material was retained for molecular analysis. Due to the technical difficulties in obtaining normal C-cell samples to compare with MTC, eight nontumor thyroid tissue samples (N; removed from the contralateral lobe of patients with micropapillary thyroid carcinoma) were also included in the series.
F, female; M, male; NA, not available; Ct, calcitonin.
For all patients serum Ct level were available at diagnosis and they were considered biochemically cured if they had a Ct value of <10 pg/mL one year after primary surgery and/or at the most recent follow-up.
All patients involved in this study gave their written informed consent and the study was approved by the local ethics committee.
RET/RAS analysis
RET exons 5, 8, 10, 11, 13, 14, 15, and 16 mutations, and H-, N-, and K-RAS exons 2 and 3 were assessed by direct sequencing, as reported elsewhere (10,18).
miRNA selection
Recent expression profiling studies identified miRNA signatures associated with MTC (16,17). We consequently explored the diagnostic performance of selected miRNAs in a series of sporadic and familial cases: (i) three miRNAs reportedly of prognostic value in a similar series of sMTC/hMTC (i.e., miR-9*, miR-183, and miR-375) (17); (ii) two further miRNAs (i.e., miR-323 and miR-370) found the most overexpressed in two cytological specimens of MTC (16); (iii) miR-224, because it is dysregulated in both follicular and parafollicular C-cell-derived carcinomas (16); (iv) two miRNAs (i.e., miR-127 and miR-154) found dysregulated in two cytological specimens of MTC (16), and reportedly also dysregulated in the presence of the RET/PCT1 rearrangement in thyroid cancer cell lines (19); and (v) miR-21, an important thyroid oncomir (13).
miRNA quantitative real-time polymerase chain reaction
Total RNA was extracted using the TRIzol reagent lysis buffer (Invitrogen, Carlsbad, CA) from fresh snap-frozen samples of 40 MTC, 2 CCH, and 8 nontumor thyroid samples, according to the manufacturer's protocol. The NCode™ miRNA quantitative real-time–polymerase chain reaction (qRT-PCR) method (Invitrogen) was used on a LightCycler 480 Real Time PCR System (Roche, Milan, Italy) to detect and quantify mature hsa-miR-9* (primer: 5′-cgc ata aag cta gat aac cga aag t-3′), hsa-miR-21 (primer: 5′-cgg tag ctt atc aga ctg atg ttg a-3′), hsa-miR-127 (primer: 5′-tcg gat ccg tct gag ctt ggc t-3′), hsa-miR-154 (primer: 5′-gtt atc cgt gtt gcc ttc g-3′), has-miR-183 (primer: 5′-ggt atg gca ctg gta gaa ttc act-3′), hsa-miR-224 (primer: 5′-gca agt cac tag tgg ttc cgt t-3′), hsa-miR-323 (primer: 5′-cac att aca cgg tcg acc tct-3′), and hsa-miR-370 (primer: 5′-ctg ggg tgg aac ctg gt-3′), and has-miR-375 (primer: 5′-cgg ctc gcg tga aaa a-3′), according to the manufacturer's instructions. Normalization was performed with the small nuclear RNA U6B (RNU6B; primer 5′-acg caa att cgt gaa gcg tt-3′). All real-time reactions, including no-template controls, were run in duplicate. Data were analyzed using the comparative cycle threshold (CT) method.
miR-21 in situ hybridization
Tissue samples from 5 sMTC and 5 normal thyroid tissues were considered for the in situ hybridization (ISH) study. ISH was performed using the GenPoint™ Catalyzed Signal Amplification System (DakoCytomation, Carpinteria, CA) according to the manufacturer's protocol. Briefly, slides were incubated at 60°C for 30 minutes and deparaffinized, as described elsewhere (20,21). Sections were treated with Proteinase K (DakoCytomation) for 30 minutes at room temperature, rinsed several times with dH2O, and immersed in 95% ethanol for 10 seconds before air-drying. The slides were prehybridized at 49°C–56°C for 1 hour with mRNA ISH buffer (Ambion, Carlsbad, CA) before incubation overnight at 49°C–56°C in buffer containing the 5′-biotin labeled hsa-miR-21 miRCURY™ LNA detection probe (Exiqon, Woburn, MA) or the scrambled negative control probe (U6; Exiqon) at a final concentration of 200 nM. The slides were washed in both Tris-buffered saline Tween-20 (TBST) and GenPoint™ stringent wash solution (54°C for 30 minutes), then exposed to H2O2 blocking solution (DakoCytomation) for 20 minutes, and then further blocked in a blocking buffer (DakoCytomation) for 30 minutes before they were exposed to primary streptavidin- horseradish peroxidase (HRP) antibody, biotinyl tyramide, secondary streptavidin-HRP antibody, and 3,3′-diaminobenzidine (DAB) chromogen solutions, according to the manufacturer's protocol. The slides were then briefly counterstained with hematoxylin and rinsed with TBST and water before mounting.
PDCD4 immunohistochemistry
From the original 40 cases of MTC, a total of 10 primary sMCT and their corresponding nontumor thyroid tissue samples were retained for immunohistochemistry. Immunohistochemical staining was done automatically (Ventana Benchmark XT System, Touchstone, AZ) for PDCD4 (catalog No. HPA001032; Atlas Antibodies, Stockholm, Sweden; 1:100), as explained elsewhere (22). Sections were lightly counterstained with hematoxylin. Appropriate positive and negative controls were run concurrently. In the cancer samples, positivity for stromal and inflammatory cells served as an internal control. Nuclear PDCD4 staining was scored on a three-tiered scale (score 0, no nuclear staining; score 1, ≥1 ≤50% positive nuclear staining; score 2, >50% positive nuclear staining) (22).
Statistical analysis
All statistical analyses were performed using the MedCal software (rel. 11.6.0). The t-test was used to assess the differences in miRNA expression levels between MTC, HCC, and normal thyroids, or in different subgroups classified by RET status and patient's outcome. Mann–Whitney test for nonparametric data was used to correlate Ct values with pathological data and the outcome of patients. A p<0.05 was considered statistically significant.
Results
miRNAs are dysregulated in MTC
By comparison with normal thyroid tissues, all hMTC and sMTC showed a significant upregulation of the nine miRNAs (p<0.0001, by t-test), with nearly no overlap between the normal and pathological specimens. In particular, the upregulation was 4.2-fold for miR-21, 6.7-fold for miR-127, 8.8-fold for miR-154, 6.7-fold for miR-224, 6-fold for miR-323, 6.3-fold for miR-370, 6.7-fold for miR-183, 10.1 for miR-375, and 13-fold for miR-9* (Fig. 1). In the two cases of CCH, the whole set of nine miRNAs were upregulated similarly to that observed in MTC, by comparison with nontumor thyroid tissues (data not shown). No difference was detected in miRNA expression between familial and hMTC cases.
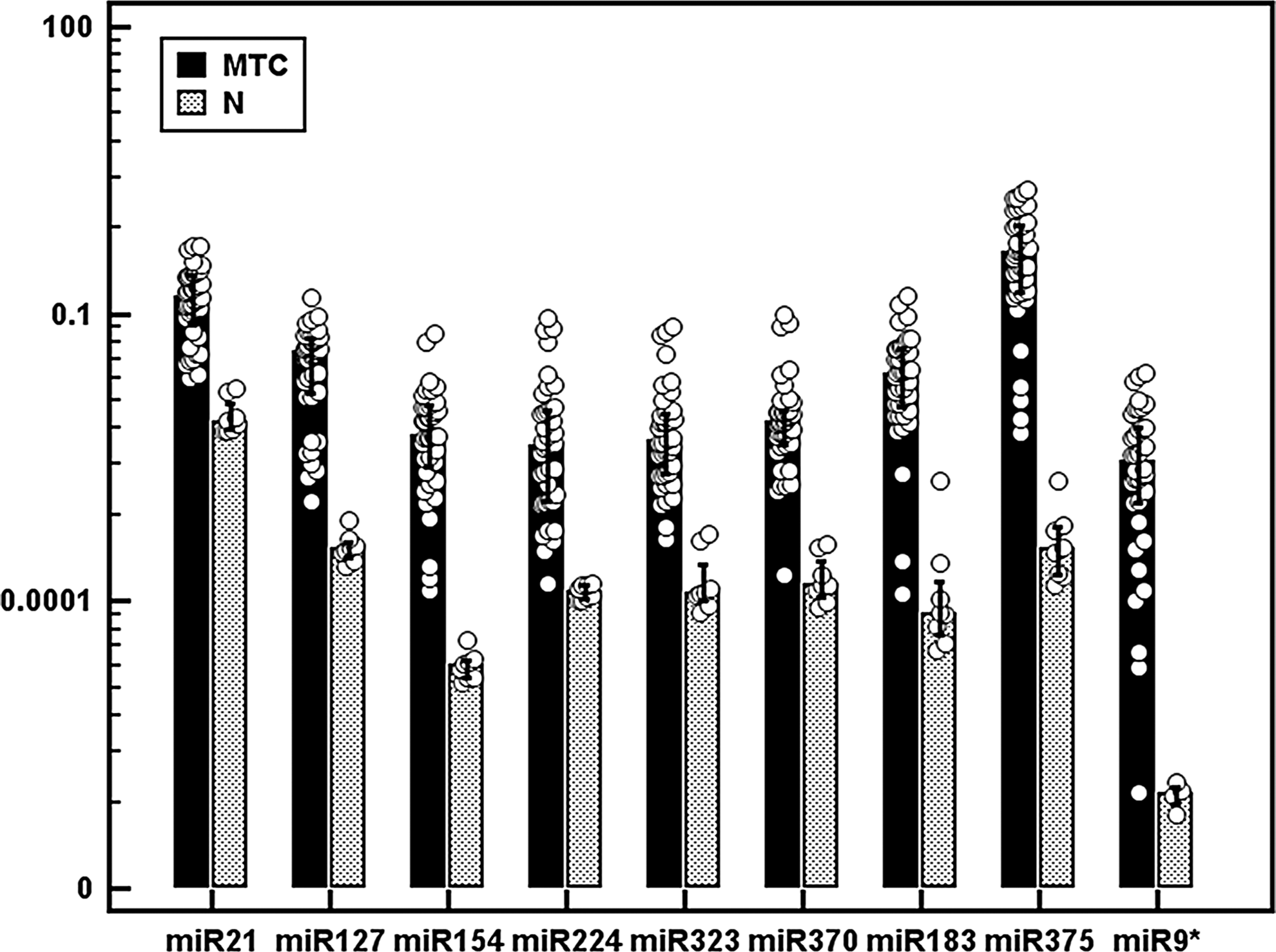
Expression levels of miR-21, miR-127, miR-154, miR-224, miR-323, miR-370, miR-183, miR-375, and miR-9* in medullary thyroid carcinoma (MTC) samples and nontumor (N) thyroid tissues by quantitative real-time–polymerase chain reaction (qRT-PCR) analysis. Each circle represents an individual sample; each bar represents the mean value of the different microRNAs (miRNAs), and vertical lines represent 95% confidence interval (CI); p<0.0001, by t-test for the nine miRNAs.
To further confirm our qRT-PCR findings, miR-21 expression was investigated in five sMTC and five normal thyroid samples by ISH analysis. miR-21 expression was identifiable as a granular brown cytoplasmic staining constantly observable in cancerous C-cells. miR-21 was significantly overexpressed in MTC by comparison with adjacent normal thyroid tissue (Fig. 2A–C).
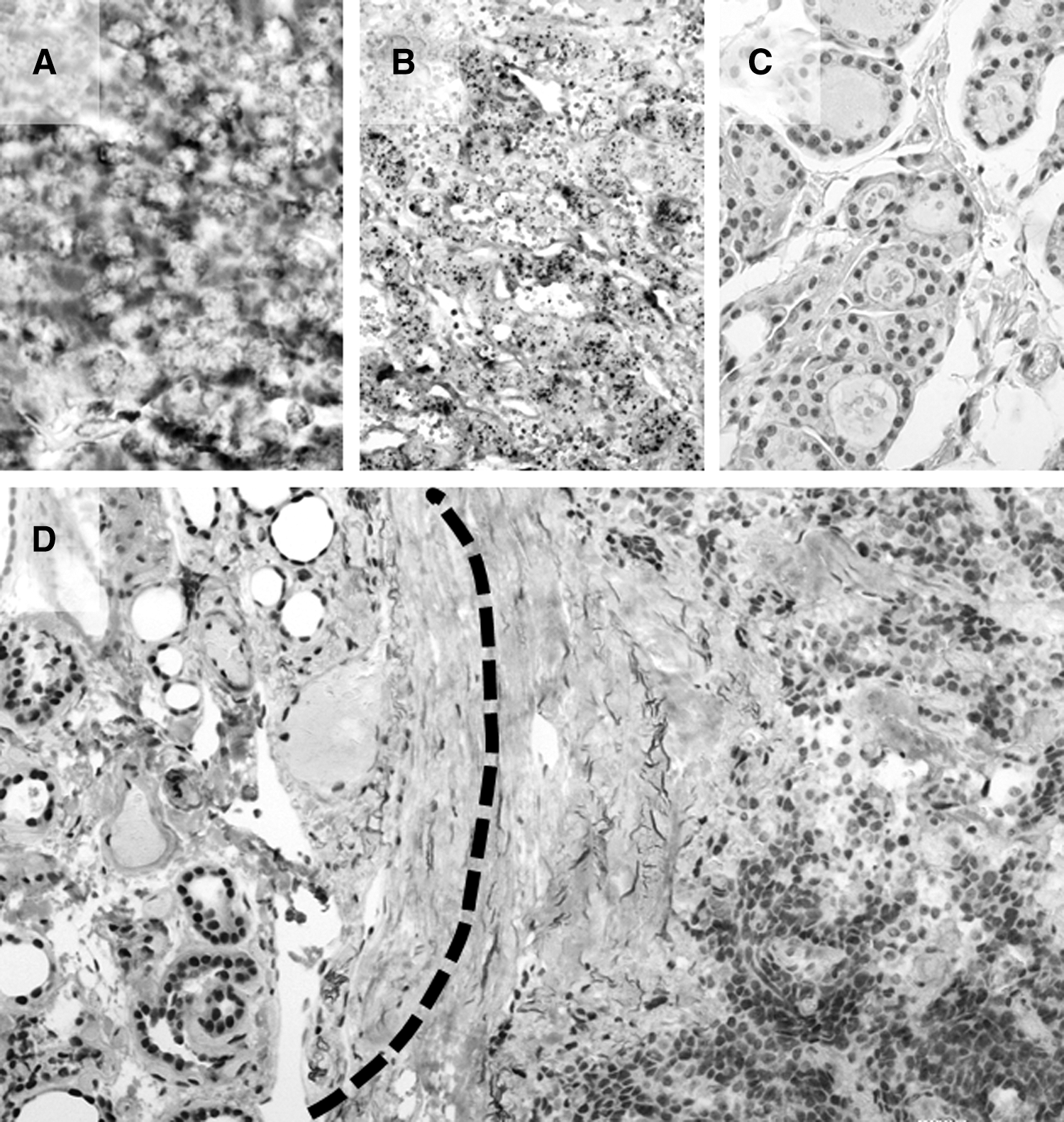
miR-21 overexpression in MTC inversely correlates with PDCD4 nuclear loss. miR-21 was significantly upregulated on in situ hybridization analysis in MTC samples
One of the mechanisms by which miR-21 can exert its oncogenic function is by directly targeting the 3′-UTR mRNA of the oncosuppressor gene PDCD4 and regulating its expression. PDCD4 expression was therefore investigated in a series of 10 MTC and their corresponding normal thyroid samples. Normal thyrocytes persistently showed a strong nuclear PDCD4 expression coexisting with a moderate-to-strong cytoplasmic PDCD4 expression (all normal samples were scored as 2). sMTC samples showed a significant nuclear PDCD4 protein downregulation (score 0=5 cases, score 1=2, score 2=3), retaining weak-to-moderate cytoplasmic staining (Fig. 2D).
Correlation with RET/RAS status and patient's pathological findings and outcome
We then evaluated the differences in miRNA expression levels in relation to (i) the presence of somatic RET/RAS mutations in sMTC, and (ii) pathological findings and the biochemical cure rate in all MTC.
RET somatic mutations were found in 50% of the cases of sMTC (17/34): the majority of the mutations were located in exon 16 at codon 918 (M918T in 8/17; 47%); mutations in exon 10 were found in three patients (18%; C611R, C618S, and C618Y); mutations in exon 11 in three (18%; 2 C630R and C634W); two patients had a 12 base pair (bp) deletion starting from codon 898 in exon 15; one patient carried a new point mutation in exon 8 (A513G, data not shown). RAS somatic mutations were present in three patients with sMTC (3/34; 9%) carrying no simultaneous RET mutation (G12R of K-RAS in one patient, Q61L of N-RAS in another patient, and M72I of H-RAS in another one).
From the whole set of nine miRNAs, only miR-127 featured a significant link with RET status; in particular, sMTC carrying somatic RET mutations showed a lower upregulation of this miRNA than those with a wild-type RET (p=0.02, by t-test). No relationships with RAS somatic mutation were found.
At the end of follow-up, 47% (19/40) of patients were biochemically disease-free judging from their latest Ct value (Table 1). As expected, initial high serum Ct levels correlated positively with the presence of nodal metastases, higher stages at diagnosis, and a “nonbiochemical-free” status during the follow-up (p=0.001, p=0.001, and p=0.002, respectively).
High miR-224 levels significantly correlated with the absence of lymph node metastases, lower stages at diagnosis, and to a biochemically free status at the end of the follow-up (p=0.01, p<0.01, and p=0.01, respectively, by t-test) (Fig. 3). No other marker was associated to pathological and/or clinical findings.
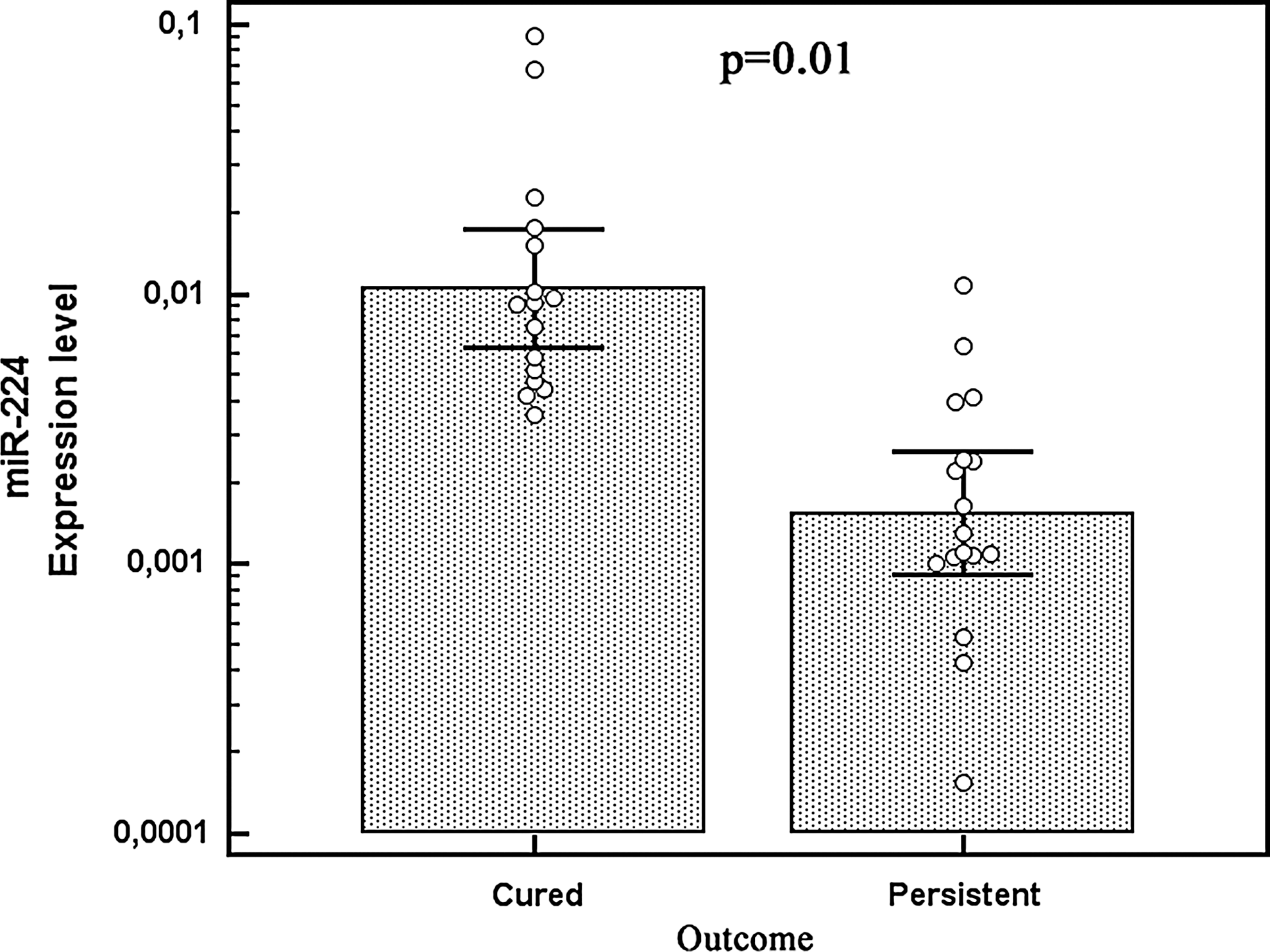
Relationship between miR-224 expression levels by qRT-PCR analysis and patient's outcome. Each circle represents an individual sample; each bar represents the mean value of the different miRNAs, vertical lines 95% CI.
Discussion
Recent studies have demonstrated that differentiated thyroid cancers are characterized by a dysregulation of different sets of miRNAs (12 –14), and distinctive miRNA expression profiles cluster significantly with a particular mutational status at somatic level, also correlating with different degrees of clinical aggressiveness (14,16).
This is at least the third report addressing miRNA analysis in hMTC and sMTC. Nikiforova and colleagues initially investigated miRNA expression profiling in two FNAB specimens obtained from MTC (16). In their seminal work, the authors found a subset of 10 specific upregulated miRNAs (16). These findings are intriguing from the diagnostic point of view because a cytological diagnosis of MTC (using FNAB) is not always feasible, particularly in the absence of substantial diagnostic clinical data (e.g., an increase in serum Ct). In this setting, miRNA expression analysis might prove an innovative tool at the pathologist's disposal to improve the diagnostic accuracy of classical cytology.
More recently, Abraham et al. investigated a series of 19 MTCs by miRNA microarray analysis and found a three-miRNA signature (miR-183, miR-375, and miR-9*) capable of differentiating adequately between familial and sporadic cases. Overexpression of miR-183 and miR-375 also proved to be a molecular predictor of extensive disease at diagnosis and pointed to a worse prognosis during the follow-up in a set of 45 MTC tissues (17).
Our results partially confirm the preliminary data emerging from a large series of MTC. As in previous reports, we established that miR-127, miR-154, miR-224, miR-323, miR-370, miR-183, miR-375, and miR-9* are upregulated in MTC (showing practically no overlap with nontumor thyroid tissues); the miR-9* featured the most pronounced increase. No reliable normal control for MTC was characterized, so we planned to expand our analysis by isolating (i.e., macro-dissecting) the material from foci of CCH in two carriers of the gene mutation undergoing prophylactic thyroidectomy. We found the same miRNA profile in CCH samples too, so—even though our analysis was limited to a small number of tissue samples—we can confidently speculate that miRNA dysregulation is an early event in medullary thyroid carcinogenesis.
miR-21 is an important oncomir that is upregulated in various different human tumors and has been found involved in thyroid carcinogenesis, and in anaplastic thyroid carcinomas too (23). Recent studies have shown that miR-21 promotes cell transformation by repressing tumor suppressor genes, such as PTEN, PDCD4, RECK, and TPM1 (21). Hence, our investigation is the first to assess miR-21 expression in MTC (supported by ISH studies). We demonstrated a significant miR-21 upregulation in cancer samples and this finding was concomitant with a marked loss of nuclear PDCD4 protein.
Whereas the overexpression of miR-9*, miR-183, and miR-375 was correlated with the type of MTC, however, our data failed to show any differences in expression in sMTC versus hMTC. This was unlike the findings reported by Abraham et al. Neither did miR-183 or miR-375 upregulation correlate with TNM stages or the patients' outcomes. One possible explanation for these discrepancies was the relatively limited number of hMTC cases, this limiting the statistical power of our results. Indeed, our series was equivalent to that of the previous report, though it contained a relatively small group of familial cases (6 vs. 19) and this, from a statistical standpoint, makes it difficult to identify any differences miRNA expression between the familial and the sporadic forms. The lack of any correlation between miR-183 or miR-375 and the clinicopathological parameters needs to be confirmed in larger case studies. On the other hand, we demonstrated that miR-224 upregulation was more pronounced in MTC diagnosed in the early stage, and in cases with a good prognosis during the follow-up.
Serum Ct concentrations are the major biomarker used to determine how advanced MTC was at diagnosis (24). The study of other molecular markers (such as peculiar miRNA signatures) warrants further consideration, however, with a view to expanding our knowledge about the mechanisms underlying the development of MTC. With this in mind, we investigated whether miRNA dysregulation coincided with a particular RET status in sMTC. Our findings showed that miR-127 overexpression is more pronounced in cancer samples carrying a wild-type RET than mutated RET, suggesting an oncogenic role for miR-127 (and its gene targets) in sMTC.
In conclusion, our data need to be confirmed in larger series of MTC and functional studies are needed to further support the biological role of miRNAs in medullary thyroid carcinogenesis. This study confirms that familial MTC and sMTC are characterized by the dysregulation of a particular set of miRNAs. This is the first finding that the degree of miR-224 upregulation provides prognostic information in patients with MTC. Thus, miRNA testing may have a future role in the clinical management of MTC.
Footnotes
Acknowledgments
This work was supported by an AIRC Regional grant 2008 (assigned to M.R.) and an MIUR grant (“Progetto di Ricerca di Ateneo”—CPDA118051; assigned to C. M.). The authors thank Frances Coburn for text editing. We also thank Cristiano Lanza, Vincenza Guzzardo, and Laura Zambonin for their excellent technical support.
Disclosure Statement
The authors have no competing interests to declare.