
Abstract
Background:
The role of viruses as environmental triggers for Hashimoto's thyroiditis (HT) is controversial. Thyroid epithelial cells express a variety of molecules involved in antiviral responses. This study combined histological, immunological, and virological tests to describe changes in tissue from patients with newly diagnosed and untreated HT. To study the early events, patients with positive thyroid peroxidase antibodies (TPO-Ab) and normal thyroid function were also included. This stage was defined as “prethyroiditis.”
Methods:
Thyroid tissue was collected from 47 patients with high titers of TPO-Ab and from 24 controls. Seventeen patients had prethyroiditis, 17 had subclinical hypothyroidism, and 13 had overt hypothyroidism. The interferon (IFN)-α/β-inducible myxovirus resistance protein 1 (myxovirus resistance protein A; MxA) was used as a surrogate marker for type I IFN expression. Inflammation, expression of MxA, and the presence of the enteroviralcapsid protein (VP1) were characterized by immunohistochemistry. The presence of enterovirus (EV) RNA was examined by in situ hybridization.
Results:
The density of CD4+ T cells was increased in all three patient groups, while CD8+ T cells were increased only in patients with overt hypothyroidism. The density of plasma cells increased as the disease progressed. The density of plasmacytoid dendritic cells and the expression of MxA were significantly increased in all patient groups compared with controls (p<0.01). EV RNA was present in 11% of HT patients, but in none of the control subjects, whereas the enteroviral protein was detected in 19% and 16%, respectively.
Conclusion:
The inflammatory reaction in the thyroid gland is a very early event in the pathogenesis of HT. The increased expression of MxA in the inflamed tissue suggests that type I IFN plays a role in disease development. Whether this is virus-dependent needs to be explored in further studies.
Introduction
Several reports indicate that viruses, such as Epstein–Barr virus (6), parvovirus B19 (7), human T lymphotrophic virus-1 (8), and herpes virus (9), might be involved in the pathogenesis of HT. However, firm evidence is still lacking. Enteroviruses (EVs) are strong candidates as they can induce low-grade persistent infection. Such low-grade viral infections may be difficult to detect using standard laboratory assays, even in tissue samples (10). EV infection has been associated with other autoimmune diseases, such as type 1 diabetes and cardiomyopathies (11 –16). The timing of sampling may be important, as viruses triggering autoimmunity may only be present temporarily in tissues because they are cleared by the immune system. Chronic inflammatory reactions will eventually destroy the endocrine tissue, which is replaced by an inflamed and fibrotic tissue. It is, therefore, important to identify both viruses and footprints of viruses at the early stage of the disease.
Thyroid epithelial cells express a variety of molecules involved in antiviral responses (17). Plasmacytoid dendritic cells (PDCs) produce large amounts of type I interferon (IFN) when triggered by viruses (18), and the density of PDCs correlates with the high number of cells expressing the IFN-α/β-inducible myxovirus resistance protein 1 (MxA) in various disorders (19). It has been demonstrated that type I IFN induce the synthesis of MxA (20). Since IFN are difficult to measure (21), we used in situ detection of MxA as a surrogate marker for the production of type I IFN.
To study early events, we also included a preclinical stage called “prethyroiditis,” which we define as the presence of autoantibodies in patients with normal thyroid function (thyrotropin [TSH] and free thyroxine [FT4] within the reference ranges). This is similar to the term “prediabetes” frequently used in the study of type 1 diabetes.
Few reports have combined both virological and immunological parameters to examine the clinically defined stages of HT, particularly the preclinical stage. Therefore, in this study, we performed a detailed examination of the inflammatory reactions and markers of viral infection, including the expression of the IFN-inducible MxA protein and the presence of EVs in thyroid glands showing different stages of HT.
Materials and Methods
Forty-seven patients were enrolled in the study. The inclusion criteria were based on the presence of autoantibodies in the serum. Serum TPO antibodies (TPO-Ab), Tg-Ab, TSH receptor antibodies (TRAb), TSH, FT4, and free triiodothyronine were measured before biopsy. All recruited patients (with elevated serum TPO-Ab and/or Tg-Ab) were divided into three groups: (i) 17 patients (36%) with prethyroiditis with normal thyroid function (normal FT4 and TSH ≤3.6); (ii) 17 (36%) patients with newly diagnosed untreated subclinical hypothyroidism (3.6≤TSH≤10 and normal FT4); and (iii) 13 (28%) patients with overt hypothyroidism (TSH>10). Only 4/47 (9%) patients were treated with LT4 for longer than 3 months. The control group comprised 24 patients with no history of autoimmune thyroiditis and antibody titers below reference ranges, who underwent neck surgery for other reasons (primary hyperparathyroidism or a suspected thyroid nodule). TRAb were measured using a competitive immunoassay (B.R.A.H.M.S.; Giagnostic GMBH), TPO-Ab and Tg-Ab a sequential immunoluminometric assay (Immulite 2500; Siemens Medical Solutions Diagnostics).
Age, gender, thyroid function, and therapy at the time of tissue sampling were all documented (Tables 1 and 2). The disease duration was estimated from the first time patients were referred for assessment of pathological blood tests. Thyroid tissue was collected by core needle biopsy. All tissue samples were obtained by a single experienced surgeon (T.P.) using a Magnum® reusable biopsy device and a 16G needle (Bard Medical, Bard Peripheral Vascular, Inc.). All biopsies were collected and stored under strict sterile conditions to minimize the risk of contamination. The specimens were fixed in 10% neutral-buffered formalin and paraffin embedded within 24 hours. Sections of formalin-fixed, paraffin-embedded (FFPE) tissue samples were routinely cut into 3-μm thick slices and mounted on slides.
ANOVA, analysis of variance; SD, standard deviation.
TSH, thyrotropin; FT4, free thyroxine; FT3, free triiodothyronine; TPO-Ab, thyroid peroxidase antibody; Tg-Ab, thyroglobulin autoantibody; TRAb, TSH receptor antibody.
Histopathological examination
The histological examination was performed on tissue sections stained with hematoxylin and eosin to determine the degree of inflammation and fibrosis (% affected area). All sections were examined by an expert pathologist (F.L.J.) to determine the percent of area with inflammation, fibrosis, and intact thyroid tissue. The pathologist was blinded for clinical and laboratory data.
Immunohistochemical examination
To characterize the inflammatory reaction, immunoenzymatic staining of FFPE tissue sections was performed using monoclonal antibodies to the following antigens: sequential staining (22) for CD4 (helper T cells, Clone 4B12; NeoMarkers, dilution 1:100) and CD8 (cytotoxic T cells, Clone C8/144B; Dako, dilution 1:20), and single staining for CD138 (plasma cells, Clone B-A38; AbD Serotec, dilution 1:100). The presence of well-organized lymphoid follicles was recorded, but the cells within these formations were not included in the cell counts.
PDCs were identified by their high expression of CD123 (IL-3R α-chain, Clone 9F5; BD Pharmingen, dilution 1:200). To ensure that the CD123+ cells were PDCs, additional costaining experiments with CD45RA were performed (23,24).
Immunohistochemical detection of IFN in FFPE sections is difficult, because many different isomers exist. MxA staining was used to detect in situ production of IFN-α. MxA+ cells were detected with anti-MxA (IgG2a, clone M143, courtesy of Dr. O. Haller, Freiburg, Germany, dilution 1:100) (25). MxA was expressed on leucocytes, endothelial cells, and epithelial cells. To perform semiquantitative comparisons of the two latter cell types, the extent and staining intensity of thyroid epithelial cells and endothelial cells were evaluated using an arbitrary scale (follicle epithelial cells: negative to ++++, vessels: negative to +++).
All stained cells within a single tissue unit (defined by the superimposition of an ocular grid representing an area of 0.058 mm2) were counted at a magnification of×400. The mean tissue area examined was 0.5 mm2. All cell counts and grading of inflammation were performed in a blinded fashion.
EV analysis
Thyroid tissue was examined for the presence of virus capsid protein 1 (VP1) by immunohistochemistry (IHC) and for the presence of EV RNA by in situ hybridization (ISH).
Immunohistochemical detection of VP1 was performed using a commercially available monoclonal anti-EV antibody (clone 5-D8/1; DakoCytomation, Denmark A/S, 1:300), an immunostainer (Ventana BencMark LT; Ventana Medical Systems, Inc.), and the UltraView™ Universal detection system, as previously described (26). EV-infected and mock-infected green monkey kidney (GMK) cells were used as controls in all separate staining batches.
ISH is based on a single EV-specific probe targeting the highly conversed group common sequence within the 5′ noncoding region of the EV genome. The probe was 3′ end-labeled with digoxigenin (DIG) using the DIG oligonucleotide tailing kit (Roche Diagnostics Ltd.). Ten picomoles of the probe were used for one labeling reaction. Tissue samples from EV-infected mice and infected GMK cells were used as positive controls in all separate staining batches (26).
Both assays cover most of EV types, and according to GenBank data, ISH detects all known EVs. Both methods have been described earlier (26 –29). Grading the positivity for the EV protein or genome was performed by two expert researchers (S.T., H.H.). Both were blinded to all the clinical data and the diagnoses of the patients and controls. Results for signals were semiquantified into the following categories: negative (no positive cells), few cells (less than 10 virus-positive cells per section in ×400 magnification), and clear positive (10 or more positive cells per section).
This clinical study was approved by the governmental Regional Ethics Committee for Medical and Health Research, South-East region, Norway. This study was conducted in accordance with the guidelines in The Declaration of Helsinki (30). Written informed consent was obtained from all participants.
Statistical analysis
Results are presented as the mean and standard deviation, or number and percentage. The one-way analysis of variance test was used for comparisons between groups. The Mann–Whitney U test was used for nonparametric data. A significance level of 5% (p<0.05) was applied when comparing patients and controls. All statistical analyses were performed using SPSS software (SPSS, Inc.) and GraphPad Prism 4 (GraphPad).
Results
Thyroid tissue was collected from 47 patients with elevated serum TPO-Ab and/or Tg-Ab and from 24 control subjects. The clinical and demographic characteristics of each group according to the clinical stage are presented in Table 2. The study patients were slightly younger than the control group, but there were no significant differences regarding age between the three patient groups. Considering that the prevalence of hypothyroidism increases with age, our groups were still quite comparable. Furthermore, there were no sex differences.
Histopathological examination
The histological examination was performed on tissue sections stained with hematoxylin and eosin to determine the degree of inflammation and fibrosis (Fig. 1). We observed a significantly larger area containing inflammatory cells in thyroid tissue of patients at all stages compared with the controls, but fibrosis was particularly apparent in tissues from patients with overt hypothyroidism. A progressive loss of the endocrine tissue correlated with increased inflammation and fibrosis. In subclinical prethyroiditis samples, inflammation and fibrosis already comprised ∼30% of the glandular tissue. We observed that the thyroid follicles became smaller in diameter in all patient groups (including at the early stage) compared with those in controls (Fig. 2A, B). The mean follicular area was 17 μm2 in the controls, 7.8 μm2 in the subclinical prethyroiditis samples, 7 μm2 in the subclinical hypothyroidism samples, and 5.9 μm2 in the overt hypothyroidism samples (p<0.01 in each patient group when compared to controls). No statistically differences were found between the three patient groups. Lymphoid follicles with germinal centers were observed, especially in tissue from patients with subclinical hypothyroidism. These organized lymphoid structures were present in 4/17 patients with prethyroiditis, 6/17 with subclinical hypothyroidism, and 2/11 with overt hypothyroidism (Fig. 2B, C, G).
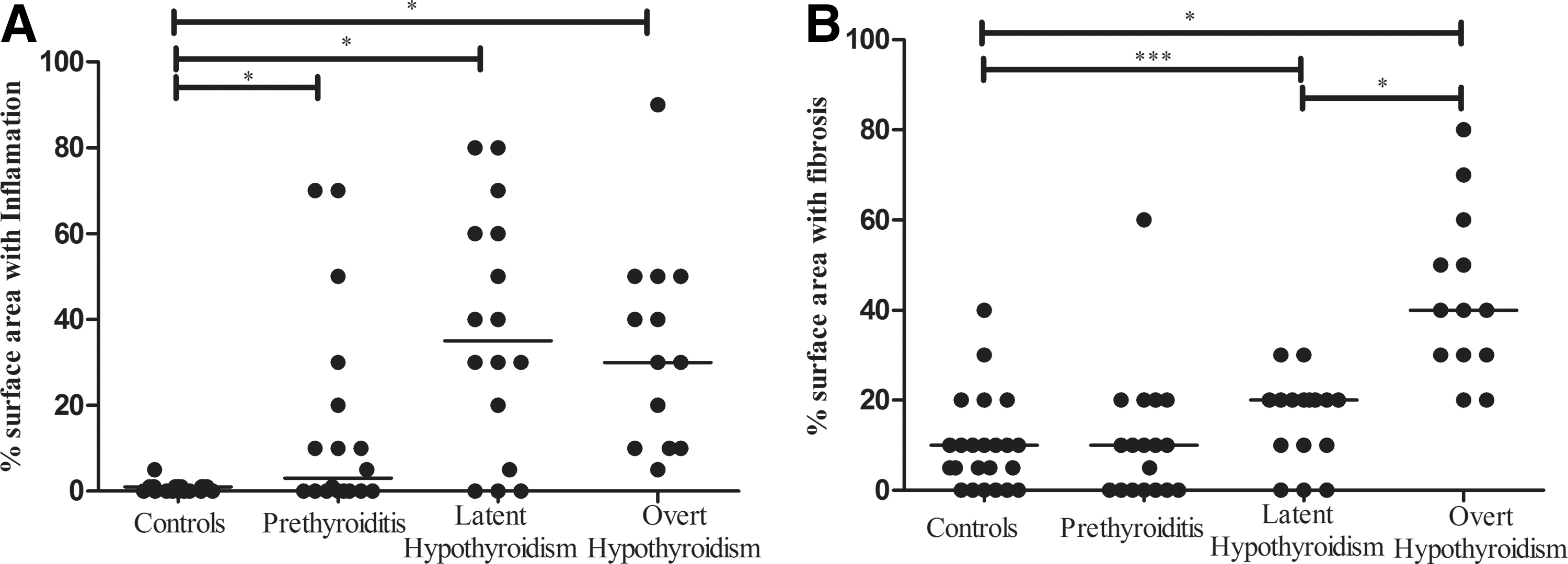
Degree of affected tissue in controls and the three subgroups of Hashimoto's thyroiditis (HT) patients.
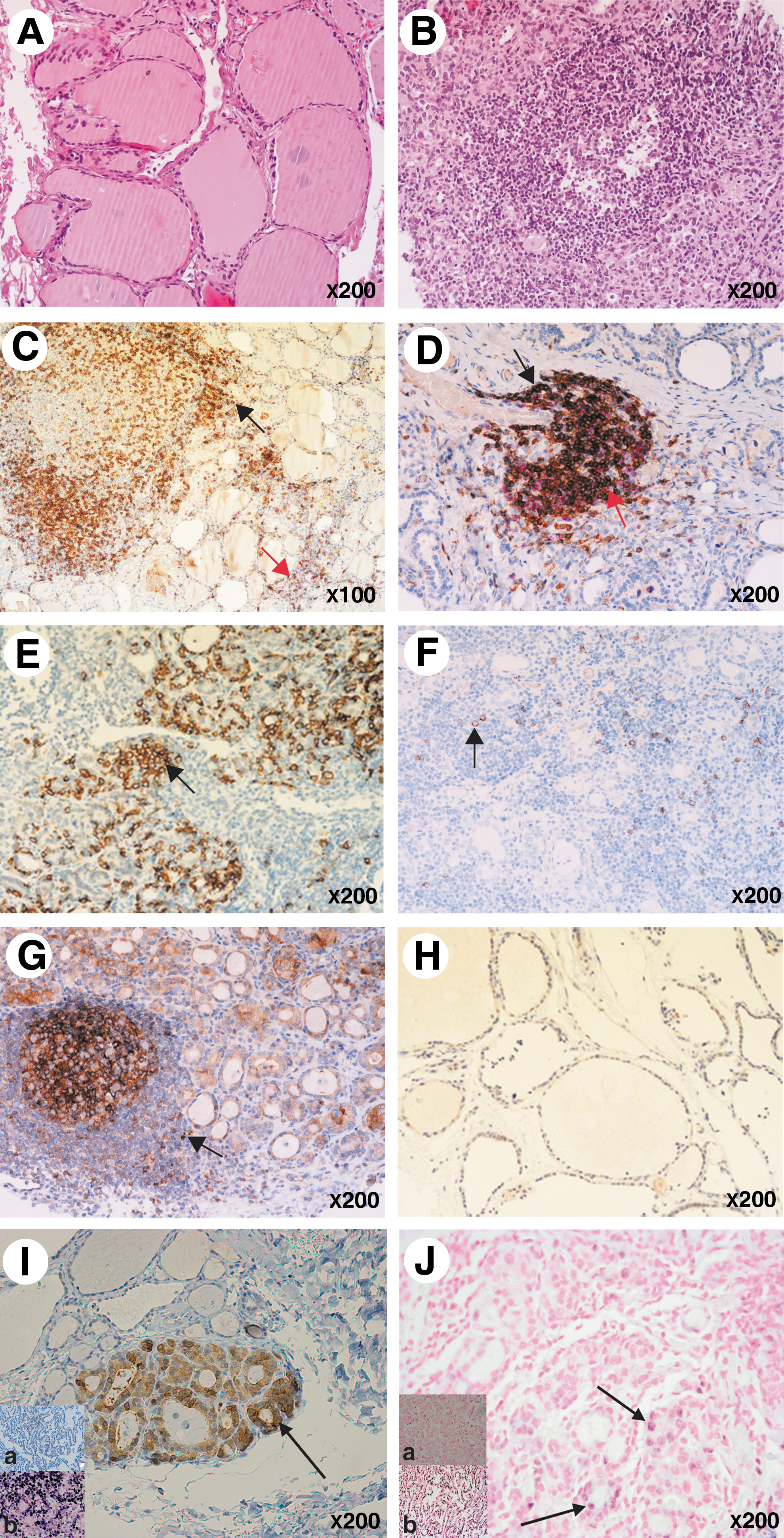
Microscopy of thyroid tissue sections. Representative examples are shown.
Immunohistochemical examination
Double staining for CD4 and CD8 showed that the number of CD4+ T cells was significantly higher in all patient groups compared with controls. Importantly, high numbers of CD4+ T cells were also observed in patients with prethyroiditis and subclinical hypothyroidism (Figs. 2C and 3A). By contrast, significantly increased numbers of CD8+ T cells were only detected in patients with overt hypothyroidism (Figs. 2D and 3B). The number of plasma cells (CD138+) paralleled that of CD4+ T cells, with numbers increasing as the disease progressed (Figs. 2E and 3C). The density of plasma cells correlated with the level of TPO-Ab (R 2=0.2; p=0.002).
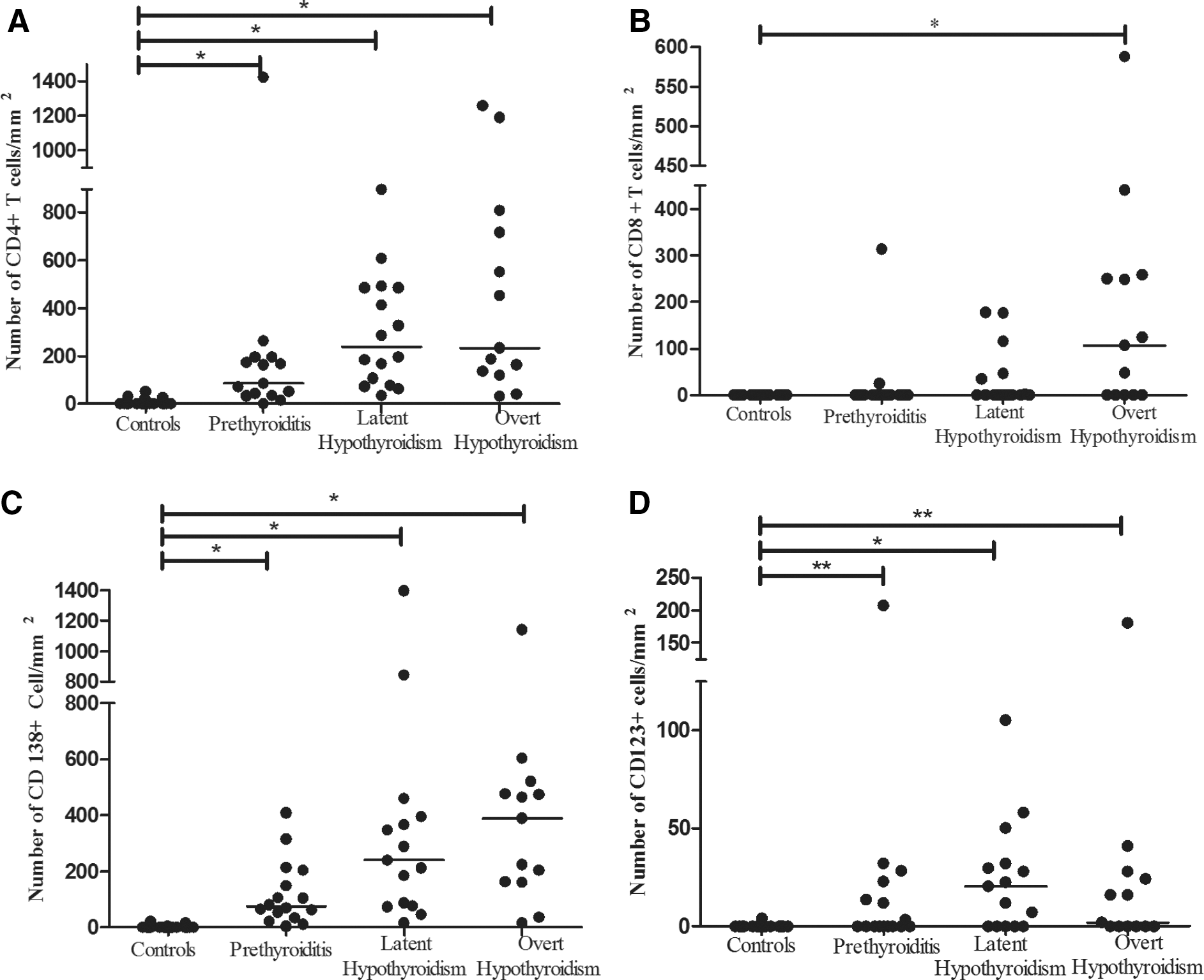
Number of cells per mm2 in formalin-fixed, paraffin-embedded sections of thyroid tissue at different stages of HT and in controls
PDCs (CD123+) were present in 24/47 (51%) of the HT thyroid tissue samples and the density was significantly higher in all patient groups compared with that in control subjects (Figs. 2F and Fig. 3D). MxA (a surrogate marker for type I IFN production) was expressed both by single cells (mainly leukocytes), epithelial cells, and endothelial cells (Fig. 2G, H). MxA-positive single cells were observed in all patient groups but not in control tissue (Figs. 2G, 4B). MxA was also expressed by follicular epithelial cells in the controls (Fig. 2H). However, a semiquantitative assessment of the extent and intensity of staining showed that MxA expression in epithelial and endothelial cells was significantly increased in the patient groups (Fig. 4A, C). Importantly, we found a positive correlation between the density of PDCs and the density of MxA+ leucocytes (R 2=0.43; p<0.001).
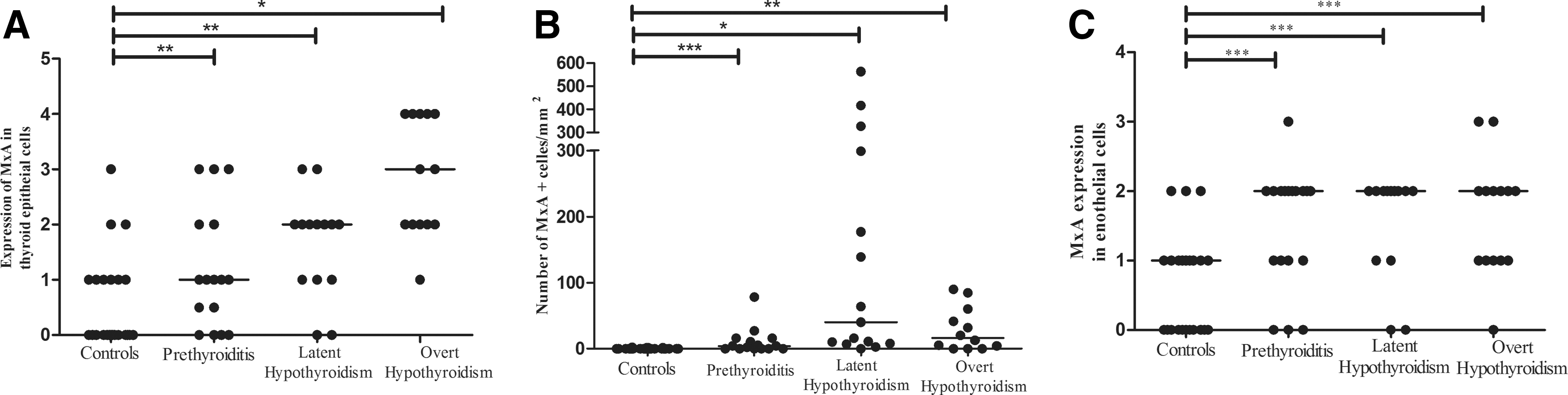
Expression of MxA in thyroid tissue from patients with HT.
EV detection
Thyroid tissue sections were analyzed for the presence of EV using two different assays. ISH identified the presence of EV RNA in thyroid tissue from 5/47 (11%) patients and in none of the control subjects (p=0.118). Virus signals were detected in thyroid epithelial cells (Fig. 2I). The presence of EV RNA was detected at all stages, including prethyroiditis. The enteroviral capsid protein, VP1, was detected by IHC in 3/17 (18%) patients with prethyroiditis (one very low positive with only few cells), in 4/17 (23%) with subclinical hypothyroidism, and in 2/13 (15%) with overt hypothyroidism. However, 4/24 (16%) of the control subjects were also positive for VP1 (two with only few cells). There were no statistically significant differences between these three groups compared with the controls (p=0.832). The virus signal (VP1) was detected in thyroid epithelial cells (Fig. 2J).
Furthermore, we could not find any significant correlation between the presence and absence of virus, and the density of PDC− or MxA+ single cells.
Discussion
This is the first study examining the thyroid tissue from patients with predominantly untreated, newly diagnosed HT, including samples taken at the very early stages of the disease (normal thyroid function tests). The histopathological examination showed pronounced tissue destruction with abundant lymphoplasmacytic inflammation and fibrosis. These tissue changes had already occurred at the prethyroiditis stage, demonstrating that tissue immunopathology is established a long time before signs of reduced glandular function become evident. The expression of MxA in affected tissue strongly suggests that type I IFN is directly involved in the pathology of HT.
Earlier studies of lymphocyte subpopulations in peripheral blood did not show any differences in CD4+ or CD8+ T-cell counts in HT compared with healthy controls (31,32). The in situ analysis of lymphocyte subpopulations in the thyroid tissue of patients with HT in the present study showed an increase in CD4+ T cells at all stages of disease compared with controls. Of particular interest, the number of CD4+ T cells was significantly increased at the prethyroiditis stage. These results are in contrast with those presented in a study by Ben-Skowronek et al. (33), although the patients in their study had a longer history of hypothyroidism and were treated with LT4. The number of CD8+ T cells was higher in the later stage of HT (overt hypothyroidism), suggesting that these cells are also involved in thyroid destruction and dysfunction. A study of NOD mice showed that both CD4+ and CD8+ T cells are required for the development of spontaneous murine thyroid disease (34). The density of plasma cells correlated weakly with the level of TPO-Ab, and these cells were present at all stages of the disease, although at significantly higher levels in overt hypothyroidism.
One important aim of this study was to examine the presence of EV in the thyroid tissue using two different methods: ISH (viral RNA) and IHC (viral capsid protein). EV RNA was indeed detected in 11% of the patients and in none of the controls, and the viral protein was detected in 19% of the patients and 17% of the controls, with no statistically significant difference between the two groups. However, the results also showed that the thyroid tissue is replaced by inflammatory cells and fibrosis, which occurs at the early stages of the disease. Therefore, it cannot be formally excluded that triggering factors present at a very early stage of the disease are cleared from the tissue before any signs of dysfunction become apparent (35). The progression of AITD is usually slow (3,36). Maternal EV infection during pregnancy has been linked to thyroiditis in children (35). In a study on unselected postoperative thyroid tissue, Desailloud et al. detected the presence of EV by polymerase chain reaction; however, the presence of virus did not correlate with thyroiditis (37). Experimental studies have shown that EVs, such as coxsackievirus B, can replicate and persist in thyroid cells (38). We focused on detecting EVs as EV infection has been associated with other autoimmune diseases (11 –16), but other viruses might also be involved, which are not detected by these methods (39).
MxA, a surrogate marker for type I IFN, was expressed in inflamed tissue, but not in the controls. Expression of MxA is strongly associated with the type I IFN, suggesting that this inflammatory cytokine is expressed. Moreover, we also found that PDCs accumulated in the affected tissue. PDCs are the major cellular source of IFN-α, and the observed positive correlation between PDCs and MxA+ leucocytes in HT suggests that PDCs produced type I IFN at the lesion site. High numbers of PDCs, MxA+ single cells and increased expression of MxA in follicular epithelial cells in HT suggest the presence of viruses or other unknown factors, which may attract PDCs and trigger them to produce IFN-α. The production of type I IFN induces an antiviral state in surrounding cells by activating downstream molecular cascades. MxA, a key mediator of IFN-induced antiviral response, is one of these downstream molecules. Therefore, at the time of examination, viruses or other triggering factors may already have been cleared from the tissue. Experimental studies suggest that PDCs play an important role in initiating autoimmune responses and type I IFN responses to viral infection (40), and PDCs exposed to virus produce large quantities of IFN-α (41). It is also well documented that therapeutic use of IFN-α can induce AITD (42,43). Furthermore, transgenic mice overexpressing IFN-α in the thyroid develop severe inflammatory thyroiditis, leading to profound hypothyroidism (44).
Our results suggest that PDCs are directly involved in the pathogenesis of HT. These cells have the capacity to migrate into the inflamed thyroid, where they presumably produce and secrete IFN-α. This fact may protect the tissue from persistent virus infection, but the subsequent inflammation may lead to the destruction and hypofunction of the thyroid tissue.
In summary, we examined the thyroid tissue from patients at different stages of HT, including the very early stage, which we clinically define as “prethyroiditis.” The progressive replacement of the endocrine tissue by inflammation and fibrosis was observed, comprising 30% of the tissue in prethyroiditis samples. The inflammatory infiltrate comprised CD4+ T cells, CD8+ T cells, and plasma cells. Accumulating PDCs, concurrent with the expression of MxA, suggest that PDCs producing type I IFN are directly involved in HT pathogenesis. Further studies are needed to evaluate the role of viruses in the development of thyroiditis. However, it is imperative that tissue sampling takes place as early as possible to increase the chance of detecting viruses.
Footnotes
Acknowledgments
K.D.-J. is the primary investigator. We thank the staff at the Hormone Laboratory research section at Oslo University Hospital, Aker, and the Pathology Laboratory at Oslo University Hospital-RH, particularly the biomedical laboratory scientists, Douglas Pauly, Jan Schwarze, and Hogne Røed Nielsen, for their help with the IHC. We also thank Eini Eskola and Eeva Tolvanen for performing virus analysis, and all the study participants.
Disclosure Statement
H.H. is a minor shareholder (<5%) in Vactech Ltd., which develops picornavirus vaccines. None of the other authors have any potential conflicts of interest to declare.