
Abstract
Background:
Organogenesis of the thyroid gland requires the Pax8 protein. Absence or reduction of Pax8 results in congenital hypothyroidism in animal models and humans, respectively. This study aims at elucidating the regulatory mechanism leading to the expression of Pax8 in thyroid cells.
Methods:
The murine Pax8 gene promoter was functionally dissected by mutagenesis and transfection in the thyroid cell line FRTL-5. Nuclear factors important for thyroid-specific gene expression were identified by DNA-binding assays.
Results:
We show that Pax8 binds to and controls the expression of its own promoter. Furthermore, we identify a novel, thyroid-specific, DNA-binding activity (denominated nTTF [for novel Thyroid Transcription Factor]) that recognizes a specific region of the Pax8 promoter.
Conclusions:
The Pax8 promoter appears to be autoregulated, a feature that might be responsible for the haploinsufficiency displayed by this gene.
Introduction
Materials and Methods
Plasmids
Plasmids containing the 14, 14.1, 14.2, and 14.3 promoter sequences were amplified from mouse genome with forward primer containing a KpnI site and reverse primer containing a BglII site. Fragments obtained were cleaved with KpnI and BglII restriction enzymes and inserted into pGL3 basic vector (Promega) containing the luciferase (LUC) reporter gene. The primers used to amplify the promoter sequences are described in Table 1.
Plasmids containing promoter deletions or point mutations (D1: −269/+100, D2: −169/+100, D3: −99/+100, D4: −46/+100, D5: del-169/−100, 14.2A7m, 14.2B3m, 14.2C4m and 14.2C5m) were obtained using the overlap extension method (19). The primers used to make promoter and point mutations are listed in Table 2. Construction of CMV-Pax8 was previously described (20).
Cells
FRTL-5 cells were grown in Coon's modified F-12 medium (Euroclone) supplemented with 5% (v/v) calf serum and a six-hormone mixture (6H), as described (21). HeLa cells were grown in Dulbecco's modified Eagle's medium (Euroclone) supplemented with 10% (v/v) fetal calf serum (Hyclone). MDCK (22), SK-N-BE (23), and R1 mouse embryonic stem cells (24) were grown as indicated in the corresponding references.
Promoter activity and DNA-binding assays and western blots
Transfections, luciferase assays, cells extracts, and gel mobility shift assays were performed as described (18). For supershift assay, the antibodies used were anti-Sp1 (sc-59; Santa Cruz), anti-Sp3 (sc-644; Santa Cruz), and anti-FLAG (F3040; SIGMA).
Western blots were performed as previously described (25). Rabbit polyclonal antibody against Pax8 was used at 0.5 μg/mL (26). Immune complexes were detected by enhanced chemiluminescence as instructed by the manufacturer (Immuno-Star Western Kit; Bio-Rad).
RNA extraction, cDNA synthesis, and real-time polymerase chain reaction
Total RNA was prepared using TRIZOL Reagent (Invitrogen) according to the manufacturer's directions. Total RNA (1 μg) was retrotranscribed using the iScript cDNA Synthesis kit (Bio-Rad). Real-time polymerase chain reaction (PCR) analysis was performed using an iCycler-iQ real-time detection system and SYBR green chemistry (Bio-Rad). Reactions were carried out in triplicate in three independent experiments. The specific primers sets used for this analysis were for amplification of β-actin (F: 5′-GGC AAT GAG CGG TTC CGA TG-3′; R: 5′-ATG GTG GTG CCA CCA GAC AG-3′) and Pax8 (F: 5′-CAG CTA TGC CTC TTC CGC TAT T-3′; R: 5′-TGT GGC TGT AGG CAT TGC C-3′). For each gene, values are means±SD of three independent experiments, normalized by the expression of β-actin, and expressed as a percentage of the value measured in parental cells.
Chromatin immunoprecipitation
Cross-linked chromatin preparation and the transcription rate measurement were performed as previously described (27). For chromatin immunoprecipitation (ChIP) with 2.5 μg of anti-Pax8 antibody or anti-RNA polymerase II antibody (Santa Cruz; sc-899) were used.
The following oligonucleotides were used for detection of immunoprecipitated chromatin fragments: Ch14.2Frat AGG GGA AGA GAA GGG TTG CAG and Ch14.2Rrat AGG GGA GGA TTT TGC AAG GGG.
Results
The minimal promoter of the Pax8 gene displays cell-type-specific activity
CNS-14, a conserved DNA sequence spanning from nucleotides −369 to +680 relative to the Pax8 gene transcriptional start site (the translation initiator codon beginning at +469), shows transcriptional activity in the thyroid cell line FRTL-5, while it is essentially inactive in HeLa cells. Such a robust thyroid-specific transcriptional activity is retained by the subregion 14.2, spanning the positions −369 to +100 (Fig. 1A). Derivatives of 14.2 (Fig. 1B) were constructed and transfected into FRTL-5 cells. The transcriptional activity of various 14.2 deletions indicates that critical elements required for a full transcriptional activity of the Pax8 promoter are located over the sequences from −369 to −100 (note the difference between 14.2 and D3). Of special relevance is the region from −169 to −99, whose deletion results in a severe reduction of transcription (compare D2 with D3 and 14.2 with D5).

Structure and activity of the Pax8 promoter.
Binding sites for nuclear factors in the Pax8 promoter
The progressive decrease of the transcriptional activity observed in the 14.2 deletions is consistent with the hypothesis that a number of cis-regulatory elements relevant for most of promoter activity are localized in the interval −369/−100. To identify the trans-acting factors recognizing this region, we decided to carry out an exhaustive analysis of the entire sequence by EMSA assays. We thus designed 19 oligonucleotides, covering the region. Each radiolabeled oligonucleotide was incubated with either FRTL-5 or HeLa nuclear protein extracts and the formation of complexes analyzed by gel electrophoresis.
We observed formation of protein–DNA complexes on oligonucleotides A2, A5, A7, B3, C4, and C5 (Fig. 2A). No detectable complex was observed on the remaining oligonucleotides (data not shown). Proteins recognizing oligonucleotides A2, A5, B3, and C5 are not thyroid specific, since they are present both in FRTL-5 and HeLa extracts. Since the B3 and C5 regions show a consensus for binding of both Sp1 and Sp3, we used supershift analysis with specific antibodies to demonstrate that indeed Sp1 and Sp3 participate to those complexes (Fig. 2B), presumably with other proteins that probably account for the different mobility of the complexes formed by the two oligonucleotides. However, since no difference was detected between HeLa and FRTL-5 cellular extracts, we did not further characterize these complexes. In contrast, oligonucleotides A7 and C4 form a main complex only with FRTL-5 extracts. We reasoned that these DNA-binding activities might be related to the thyroid-specific expression displayed by the 14.2 sequence and they were characterized further.
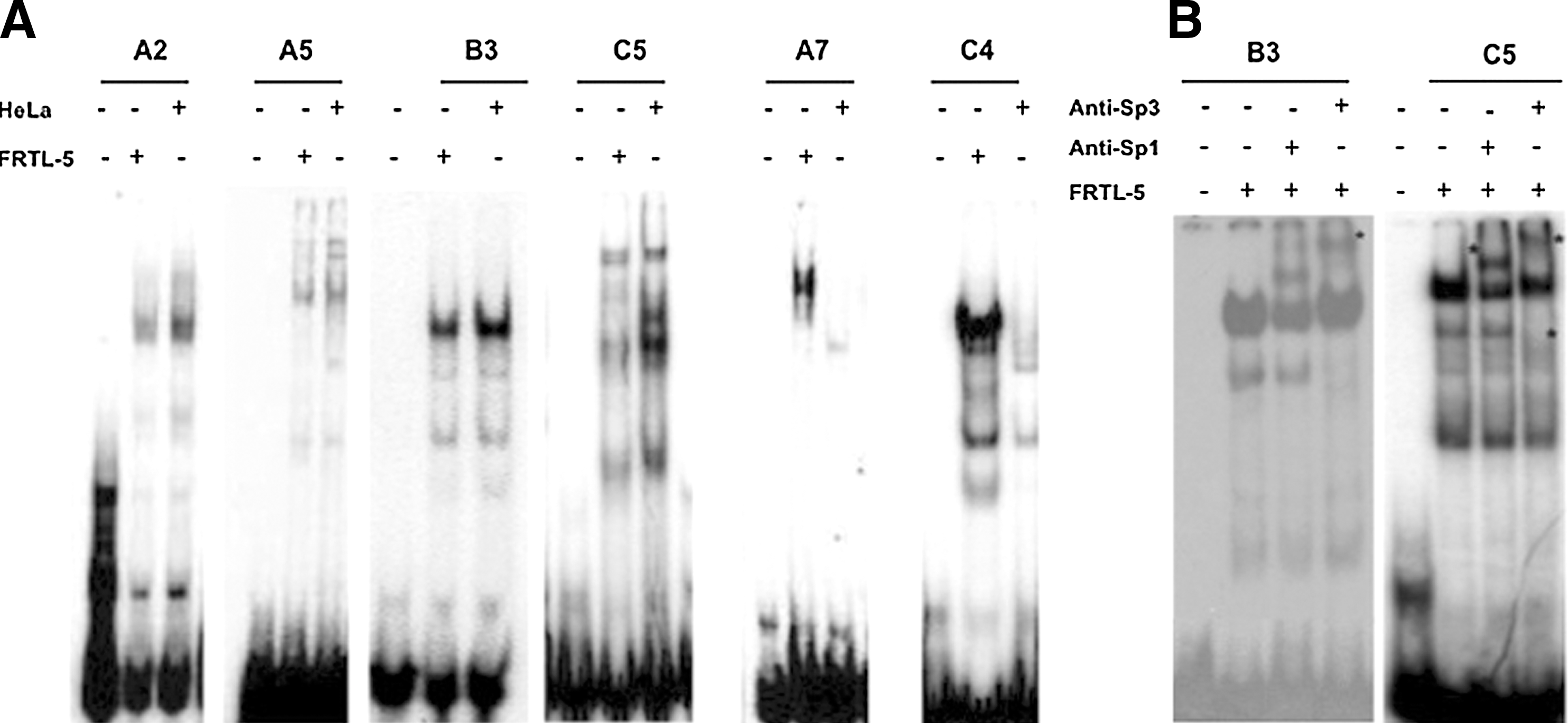
Binding sites for nuclear factors in the 14.2 sequence.
Pax8 and an unknown thyroid-specific protein bind to the Pax8 promoter
In the sequence of oligonucleotide C4 region, we identified a consensus recognition sequence for Pax8, strongly conserved in the Tg and TPO promoters (Fig. 3A). To test whether the Pax8 protein actually binds to the C4 sequence, radiolabeled oligo-C4 was challenged with a purified, bacterially produced, Pax8-paired domain (bPD-PAX8). As shown in Figure 3B, bPD-PAX8 binds to the sequence C4. To further analyze the C4/Pax8 complex, the C4 oligonucleotide was challenged with protein extract from HeLa cells transfected with the Pax8-FLAG vector. As Figure 3C shows, the C4 oligonucleotide forms a complex only with the HeLa cells extract prepared from cells transfected with the Pax8-FLAG vector. Furthermore, this protein complex is efficiently supershifted with the anti-FLAG antibody. Taken together, these data demonstrate that Pax8 recognizes the C4 sequence.
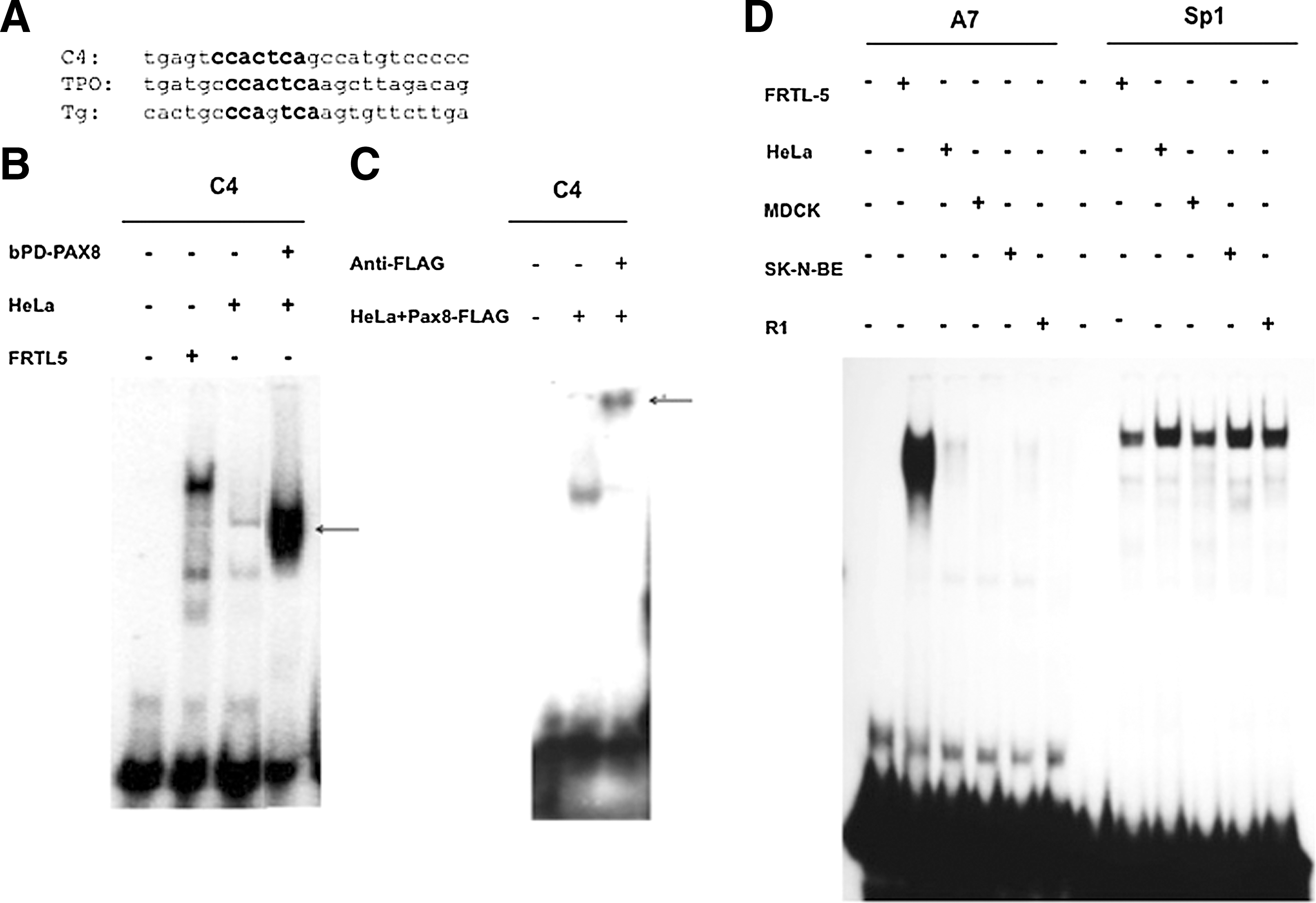
Thyroid-specific transcription factors bind to the 14.2 sequence.
To further determine the thyroid specificity of the activity recognizing the A7 sequence, we tested nuclear extracts from different cell types. As shown in Figure 3D, a complex with the A7 oligonucleotide is only observed with those of differentiated thyroid cells among the several extracts tested, even though all extracts are capable of recognizing an oligonucleotide containing the Sp1 recognition sequence. Furthermore, the A7/protein complex is not competed away with oligonucleotide binding the known thyroid-enriched TFs such as Nkx2-1/Titf1, Foxe1, Pax8, and Hhex (data not shown). We conclude that this oligonucleotide identifies a novel thyroid-specific DNA-binding protein (referred to as nTTF, a novel thyroid transcription factor).
Both Sp1/Sp3 and Pax8 are necessary for Pax8 promoter activity
The cis-elements identified in the Pax8 promoter are summarized in Figure 4A. To test the functional relevance of some of the binding sites identified, specific mutations were introduced in pGL14.2 at Sp1/Sp3 binding sites and in the C4 and A7 sites. We first demonstrated that the mutations introduced completely abolish the binding of the corresponding protein (data not shown). The activity of wild-type and mutant promoters was assessed upon transfection in FRTL-5 cells. As Figure 4B shows, the mutated construct pGL14.2A7m, which abolishes the binding of nTTF, displayed the same activity as the control pGL14.2. In contrast, both pGL14.2B3m and pGL14.2C5m, in which either of the binding sites of Sp1/Sp3 factors has been disrupted, exhibit a reduction in promoter activity to 50% of the wild type. Finally, the pGL14.2C4m construct, which abolishes Pax8 binding at that site, also shows reduced transcription activity to 50% of that displayed by the control pGL14.2. We also generated pGL14.2B3/C4/C5m combining the above-described mutations at B3, C4, and C5 sites. The combination of the three mutations led to a stronger reduction of transcription.
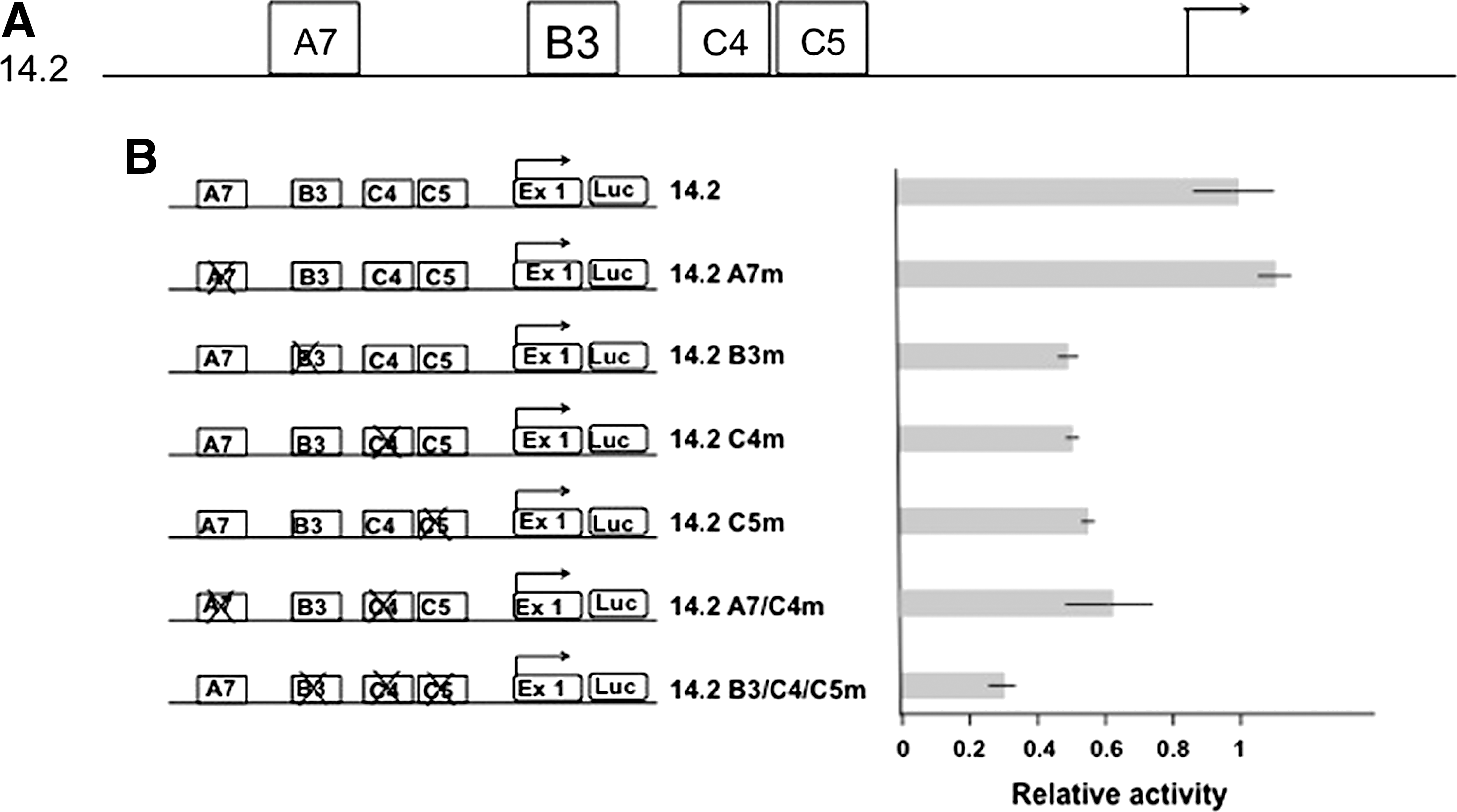
Functional analysis of the murine Pax8 promoter.
The results of our experiments indicate that the binding of both ubiquitous factors Sp1/Sp3 and of the thyroid-specific Pax8 transcription factor is required to achieve full Pax8 promoter activity. In contrast, no evidence could be obtained on the functional relevance of the binding of nTTF to the A7 sequence.
Pax8 stimulates transcription of its own promoter
We next asked whether the Pax8 promoter could be activated in HeLa cells by the Pax8 protein. To this end, we cotransfected pGL14.2 into HeLa cells, together with an expression vector encoding Pax8. As Figure 5 shows, the presence of Pax8 increases transcription from pGL14.2 about fivefold; in contrast, the activity of the mutated promoter pGL14.2 C4m, unable to bind to Pax8, is not affected by the expression of this transcription factor. These findings show that binding of Pax8 protein is able to activate its own promoter.
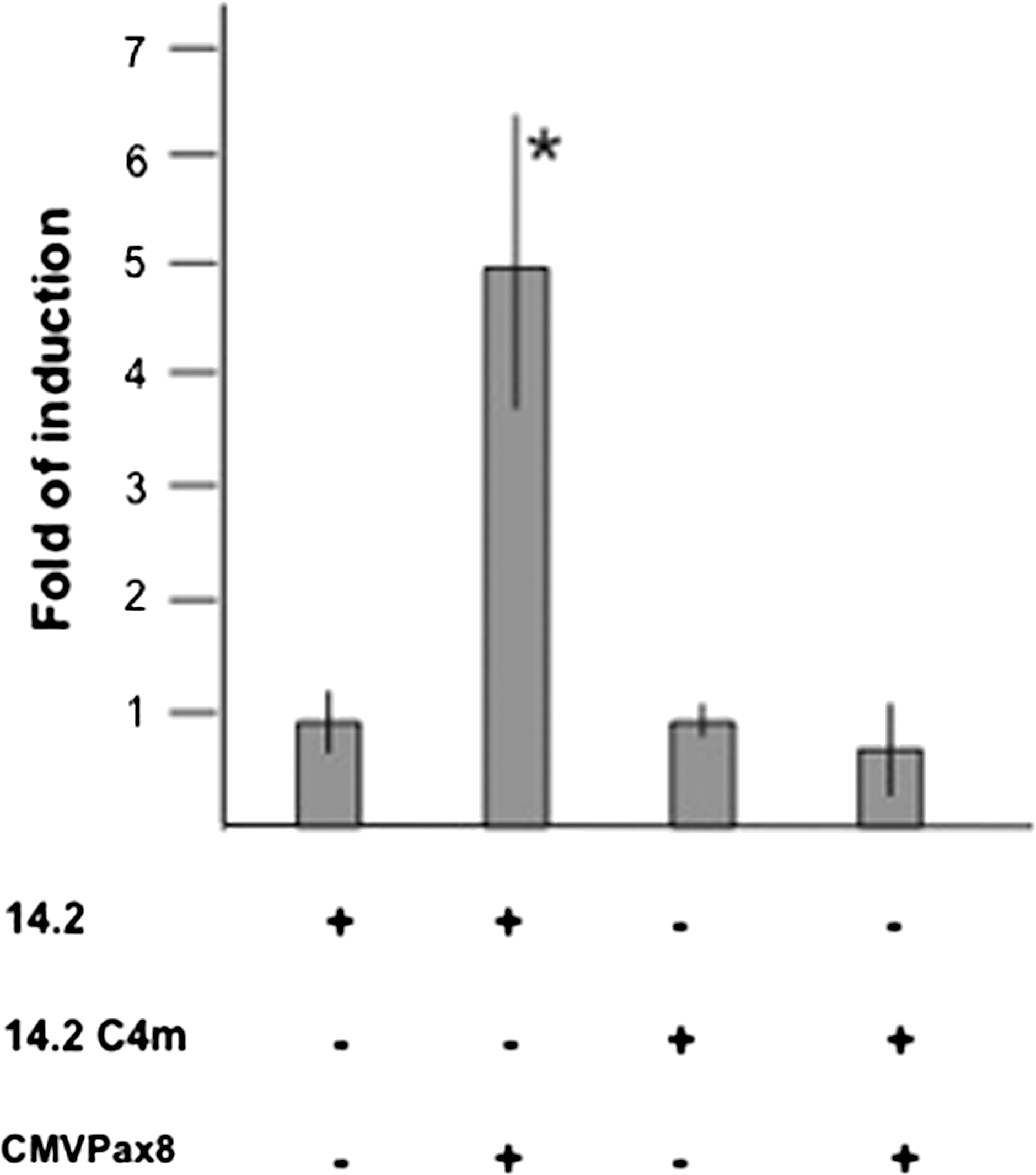
Pax8 protein activates its own promoter. 14.2 and 14.2 C4m (containing a mutated Pax8-binding site) were transiently transfected in HeLa cells in the absence or in the presence of the expression vector encoding Pax8 (CMV-Pax8). Data are expressed as fold induction over the activity of 14.2 in absence of CMV-Pax8, whose value was set at 1.0. Values are mean±SD of at least three independent experiments each performed in triplicate. *p<0.05 by unpaired, two-tailed Student's t-test.
To gain some evidence on the relevance of Pax8 in controlling its own promoter in a chromosomal context, we studied Pax8 promoter activity in the FRTL-5/ER-Ras cells carrying a tamoxifen-inducible HRasV12 -ER allele; in these cells, Pax8 protein and mRNA levels (27,28) drastically decrease upon H-RASV12ER activation by tamoxifen. We performed ChIP assays on genomic DNA from FRTL-5/ER-Ras cells treated or not treated with tamoxifen. The crosslinked chromatin was immunoprecipitated using either an antibody against Pax8 or an unrelated, control antibody. The enrichment of the Pax8 promoter region was monitored by real-time PCR amplification (see the Materials and Methods section). Figure 6A shows that the Pax8 antibody is able to immunoprecipitate the chromatin containing the promoter; however, after tamoxifen treatment, the amount of immunoprecipitated chromatin containing the promoter is diminished, matching the reduced expression of the Pax8 protein (Fig. 6B). In addition, as an indicator of amount of transcription from the Pax8 promoter, we immunoprecipitated the cross-linked chromatin using an antibody against RNA polymerase II (Fig. 6C). The assay shows that the amount of RNA polymerase II bound to the Pax8 gene is reduced 24 hours after tamoxifen treatment, thus supporting the regulatory role of Pax8 in maintaining adequate levels of transcription from its own promoter.
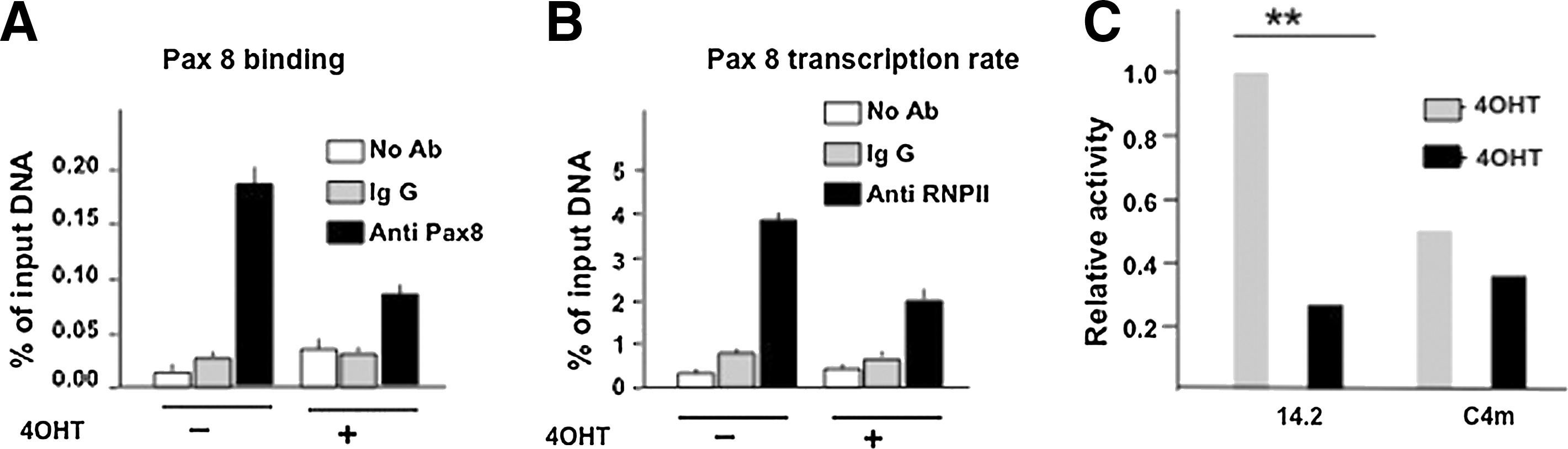
Pax8 binding to its own promoter is associated with a transcriptional effect.
The data reported herein confirm that Pax8 binds to its own promoter in vivo and suggest that this binding is associated with a transcriptional effect. To gain further support for the role of Pax8 in the antiregulatory loop, we transfected the pGL14.2 into FRTL-5/ER-Ras cells and evaluated its ability to drive the expression of the reporter gene. As expected, after tamoxifen induction, the transcriptional activity of the promoter is strongly reduced. In contrast, the activity of the mutated promoter pGL14.2 C4m, unable to bind to Pax8, failed to significantly change in presence of an active H-RASV12 (Fig. 6D). This latter experiment nicely correlates the role of Pax8 in regulating transcription from its promoter, both if inserted in its natural chromosomal context and if present in a transfected chimeric construct.
Discussion
We show in this study that the genomic region abutting the transcription start site of the Pax8 gene displays promoter activity in a thyroid-cell-specific manner. Furthermore, we provide data to support the notion that this cell-type-specific expression might be explained, at least in part, by the binding of Pax8 itself. Thus, based on these findings, we propose that thyroid cells have an autoregulatory loop that maintains physiological levels of the Pax8 protein, a feature already proven to be operating in other members of the Pax gene family (29). This observation might explain the haploinsufficiency of PAX8, which has been deduced by the development of congenital hypothyroidism in humans carrying inactivating mutations of this gene in heterozygosity (11,30). Indeed, if Pax8 contributes in maintaining its own expression, then a mutation resulting in the presence of only one allele producing a functional protein could result in a more dramatic reduction in overall protein levels, because also the transcription of the wild-type allele could significantly decrease. In this respect, the recent finding of a mutation in the human PAX8 promoter associated with a hypothyroid phenotype (31) is of relevance, since it strongly underlines the importance of delineating the regulatory region of this gene, also for a better understanding of human thyroid dysgenesis. It should however be pointed out that the mutation identified in the human PAX8 gene is located in a region of the human promoter that does not show significant similarity with the mouse promoter used in the present study (Fig. 7). However, it has been clearly shown that despite the conservation in the transcription factors involved, a significant divergence exists between human and mouse in promoter architecture (32). Thus, it is conceivable that also in the case of the Pax8 promoter, similar factors might be involved in mice and humans, but given their diversified arrangement, it is not easy to locate them without a previous identification of the cognate proteins. The same reasoning applies to the thyroid-specific factor recognizing the A7 region, for which at present no data exist on the conservation between the two species, even though 18/24 nucleotides are conserved in the corresponding position in the human promoter (data not shown).

Architecture of the mouse Pax8 promoter. The sequence of the mouse Pax8 promoter (M) is shown, aligned with the human sequence (H). Dashes indicate spacing introduced to maximize the alignment between the two sequences. In the human sequence, only the nucleotide changes with the mouse sequence are reported. The C-to-T substitution labeled Patient is the mutation reported in a patient with congenital hypothyroidism (30). The location of the binding sites for nuclear factors demonstrated in this study is also indicated. The arrow in the bottom line of the mouse sequence indicates the transcriptional start site.
While the data reported in this article suggest a simple mechanism for maintenance of Pax8 gene expression, and hence of thyroid cell differentiation, based on an autoregulatory loop, it remains to be explained how transcription of Pax8 is initiated. While this study fails to address this issue, it is tempting to speculate that the thyroid-specific protein recognizing the A7 sequence that we have called nTTF might be involved in the process. The reason for which transcription of a transfected Pax8 promoter in cultured thyroid cells appears to be unaffected by mutation in the A7-binding site can be due to several factors. First, nTTF might be operating only in a chromosomal context. Another hypothesis envisages a separate control for initiation and maintenance of Pax8 gene expression, as appears to be the case during mouse thyroid development (4). Such diverse mechanisms for initiation and maintenance of gene expression have been demonstrated first in bacteriophages (33) and have also been documented in plants (34) and animals (35 –38). Hence, it is conceivable that in differentiated thyroid cells, Pax8 itself has taken over the control of its own transcription, and thus there is no strong requirement for an additional thyroid-specific protein such as nTTF.
The identification of nTTF or of the factor binding to the site mutated in a patient with congenital hypothyroidism and the assessment of their role in development and differentiation of the thyroid gland will be essential to distinguish among these hypotheses. It will also be of interest to assess whether similar mechanisms operate in other, recently discovered, sites of Pax8 expression (39,40), and also whether their aberrant regulation might explain, at least in part, the involvement of this transcription factor in cancers (41 –43).
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.