
Abstract
Background:
Deiodinases (DIO1, 2, and 3) are key enzymes in thyroid hormone (TH) activation and inactivation with impact on energy metabolism, development, cell differentiation, and a number of other physiological processes. The three DIO isoenzymes thus constitute sensitive rate-limiting components within the TH axis, prone to dysregulation by endocrine disruptive compounds or disease state. In animal models and cell culture experiments, they serve as readout for local TH status and disarrangement of the hormonal axis. Furthermore, some human diseases are characterized by apparent deiodinase dysregulation (e.g., the low triiodothyronine syndrome in critical illness). Consequently, these enzymes are targets of interest for the development of pharmacological compounds with modulatory activities. Until now, the portfolio of inhibitors for these enzymes is limited. In the clinics, the DIO1-specific inhibitor propylthiouracil is in use for treatment of severe hyperthyroidism. Other well-known inhibitors (e.g., iopanoic acid or aurothioglucose) are nonselective and block all three isoenzymes. Furthermore, DIO3 was shown to be a potential oncogenic gene, which is strongly expressed in some tumors and might, in consequence, protect tumor tissue form differentiation by TH. With respect to its role in tumorigenesis, specific inhibitors of DIO3 as a potential target for anticancer drugs would be highly desirable. To this end, a flexible and convenient assay for high-throughput screening is needed. We recently described a nonradioactive screening assay, utilizing the classic Sandell–Kolthoff reaction as readout for iodide release from the substrate molecules. While we used murine liver as enzyme source, the assay was limited to murine DIO1 activity testing. Here, we describe the use of recombinant proteins as enzyme sources within the assay, expanding its suitability from murine Dio1 to human DIO1, DIO2, and DIO3.
Methods:
As proof-of-concept, deiodination reactions catalyzed by these recombinant enzymes were monitored with various nonradioactive substrates and confirmed by liquid chromatography–tandem mass spectrometry.
Results:
The contrast agent and known DIO inhibitor iopanoic acid was characterized as readily accepted substrate by DIO2 and Dio3. In a screening approach using established endocrine disrupting compounds, the natural food ingredient genistein was identified as a further DIO1-specific inhibitor, while xanthohumol turned out to potently block the activity of all three isoenzymes.
Conclusions:
A rapid nonradioactive screening method based on the Sandell-Kolthoff reaction is suitable for identification of environmental, nutritive and pharmacological compounds modulating activities of human deiodinase enzymes.
Introduction
D
Genetic inactivation of Dio2 in mice results in, for example, inner ear defects (4) and impaired immunological function (5). In the clinics, dysregulated DIO expression is considered of key importance during the development of the low-triiodothyronine (T3) syndrome in critical illness (6) and overexpression of DIO3 in juvenile hemangioma results in consumptive hypothyroidism (7). Overexpression of the TH-inactivating isoenzyme DIO3 is also proposed as an underlying cause of tissue dedifferentiation and tumor growth (e.g., in colorectal cancer or basal cell carcinomas) (8).
We have recently described a nonradioactive method to determine murine Dio1 activities (9). It depends on the iodide release from various substrate molecules, utilizing endogenous Dio1 activity from murine liver extracts. Subsequently, liberated iodide is quantified via a classical iodide determination reaction, the Sandell–Kolthoff (SK) reaction (10), as it was recently described for cellular iodide uptake assays (11).
Alternative assay setups have several significant disadvantages. The classical DIO assay relies on the liberation of radioactive iodide from an expensive tracer substrate. Therefore, the method depends on the availability of specifically trained human resources, a radiotracer substance with limited half-life, and a radioactive lab workplace. Furthermore, waste handling is costly and an issue of legal regulations for radioactively contaminated material and liquids. A more technically sophisticated method includes separation and identification of THs via high-performance liquid chromatography, but also depends on radioactively labeled tracer molecules with the mentioned disadvantages in availability and handling (12).
The third and most up-to-date option is liquid chromatography–tandem mass spectrometry (LC-MS/MS) (13). This method allows analyses of metabolites of all available DIO substrates and enables quantification of both the educt and subsequently formed products within the DIO-driven reaction (14). Unfortunately, LC-MS/MS analytics depends on costly equipment, specially trained employees, specific internal standards, and sensitive preanalytical extraction steps, which all are crucial for obtaining reliable results. Overall, just a few laboratories are equipped and trained for using this state-of-the-art method in the area of analytics of low abundant hormones.
Our nonradioactive assay was recently described for high-throughput screening (HTS) purposes using hepatic murine tissue as a source of enzymatic activity (9). We characterized it as an affordable and straightforward method. The protocol is applicable for almost every lab in the basic sciences. As Dio2 and Dio3 activity are not found in healthy tissues in appropriate amounts, we were limited to murine Dio1 activity measurements.
To close the gap of interspecies differences, we have now expanded the spectrum of the assay by using recombinant human DIO enzymes expressed in human embryonic kidney 293 (HEK293) cells. These cells have no detectable endogenous DIO activity but are able to correctly synthesize active selenoproteins and produce sufficient enzymatic activity under selenium (Se)-supplemented conditions.
As a follow-up to our recent study, we tested the unspecific DIO inhibitor iopanoic acid (IA) as a potential substrate for human DIO2 and murine Dio3, as it was previously shown for murine Dio1.
We now applied our screening approach to potential inhibitors/endocrine disruptors of human DIO1, DIO2, and DIO3 activities. The overall substrate acceptance and preference was investigated in a side-by-side approach with the nonradioactive HTS variant and LC-MS/MS analytics.
On this fundament, a panel of various compounds with known effects on the TH axis was screened for interaction with enzymatic deiodination. Two compounds were identified as potential DIO inhibitors: xanthohumol (XN), a prenylated chalconoid from hops, inhibited all three isoenzymes while genistein (GEN), a phytoestrogen from soybeans, exclusively blocked DIO1 without interfering with DIO2 and DIO3 activities. The calculated half maximal inhibitory concentration (IC50) values were in the lower μmolar range indicating that these substances may serve as valuable lead compounds for further pharmacological development.
Material and Methods
Materials
All chemicals were of analytical grade and obtained from Merck or Sigma-Aldrich. Iodothyronines were purchased from Sigma-Aldrich and Formula GmbH. Daidzein was a generous gift of Dr. Sosic-Jurjevic (University of Belgrade, Serbia). Aurothioglucose (ATG), propylthiouracil (PTU), and Dowex W50-X2 were acquired from Sigma-Aldrich and were solved as stock solutions in double-distilled water or 40 mM sodium hydroxide, respectively. The origin and handling of the various test compounds (Table 1) and THs has been described elsewhere (13,15). The vacuum plate prep manifold was from SUPELCO, and 96-deep well plates as well as filter plates, used to build the Dowex columns, were taken from the Eppendorf Perfectprep Plasmid 96 Vac Direct Bind kit as described before (9).
Compounds chosen because of their known interactions with the triiodothyronine (T3) receptor (15). The identities (IDs) are the same as those shown in Figure 3A.
TR, thyroid hormone receptor.
Preparation of cell lysates with DIO activities
The open-reading frames for all three human deiodinases were amplified via polymerase chain reastion from a cDNA library. The fragments for human DIO1, 2, and 3 and the murine Dio3 were ligated into an expression plasmid, pcDNA3. The plasmid backbone already contained a human GPX4-selenocysteine insertion element, as described before (16). Furthermore, all open reading frames were modified to contain a FLAG-tag sequence (CTT GTC ATC GTC ATC CTT ATA ATC CTT GTC ATC ATC CTT ATA ATC) at the C-terminus. In the beginning of our experimental work, HEK293 cells were transiently transfected with polyethyleneimine-40 as transfection agent. Later on, expression cassettes were subcloned into the expression vector pCDH-puro and stable HEK293 cell lines were established by puromycine-selection. For optimal DIO expression, all cell populations were cultured in Dulbecco's modified Eagle's medium:F-12 (
After a washing step with ice-cold phosphate-buffered saline, cells were harvested by scraping in homogenization buffer [250 mM sucrose, 20 mM HEPES (pH 7.0), 1 mM EDTA, and 1 mM dithiothreitol (DTT)]. After sonification (3×15 pulses on ice), the total cell homogenate was checked for detectable DIO activities by the nonradioactive approach, protein concentration was determined by Bradford reagent (BioRad) and enzyme stocks were adjusted to 10 μg/μL protein and frozen at −80°C.
Deiodination experiments with iodothyronines
Deiodination reactions were prepared in quadruplicates in 250 μL reaction vessels with a total volume of 100 μL each. Final concentrations were 0.1 M potassium phosphate (pH 6.8 for DIO1 and 2, pH 8.0 for Dio3), 40 mM DTT and with or without inhibitor (1 mM PTU for DIO1, 1 mM ATG for DIO2 and Dio3). A broad spectrum of iodo-L-thyronines (T4, T3, rT3, 3,3′-T2, 3′,5′-T2, 3,5-T2, 3-T1, and 3′-T1 at final concentration of 5 μM) were incubated with an appropriate amount of enzyme-containing homogenate (10 μg for DIO1, 100 μg for DIO2, and 25 μg for Dio3) over a time period of 4 h at 37°C with constant shaking.
As negative control, homogenates from HEK293 cells without recombinant DIO were used or reactions were performed in which the activities were blocked by ATG (DIO2 and 3) or PTU (DIO1).
Subsequently, one reaction vessel per TH assay was transferred to the LC-MS/MS analytics (Supplementary Fig. S1; Supplementary Data are available online at
Deiodination of iopanoic acid
Iopanoic acid was incubated with recombinant DIO1, DIO2, or Dio3 under optimal, enzyme specific conditions for up to 90 min at 37°C. In parallel, the preferred substrates of the individual isoenzymes were incubated under the same assay conditions as control experiments. Subsequent processing of the samples was as described before.
Screening of endocrine active substances as potential inhibitors
Various compounds (Table 1) were tested for their inhibitory potential on DIO activities. All DIO isoenzymes were incubated with their preferred substrates under optimal assay conditions and in the presence or absence of individual test compounds. In all cases, the final concentration of substrates was chosen to correspond to 10 μM (Table 1).
Compounds with significant inhibitory effects >30% (positive candidates) were taken into further experiments to evaluate their inhibitory potential in more detail. To this end, dilution series of XN and GEN were tested in the DIO isoenzyme–specific assays along with the appropriate controls (0.1% dimethyl sulfoxide). IC50 and nonlinear regression curves were calculated by Graphpad PRISM (GraphPad Software, Inc.).
Results
Deiodination experiments with iodothyronines
For all three DIO isoenzymes, a characteristic substrate preference spectrum was achieved, essentially mirroring the facts known from the literature (13,17) (Fig. 1B–D). DIO1 efficiently deiodinated rT3, while the best substrates for DIO2 and Dio3 were (thyroxine) (T4) and T3 respectively.
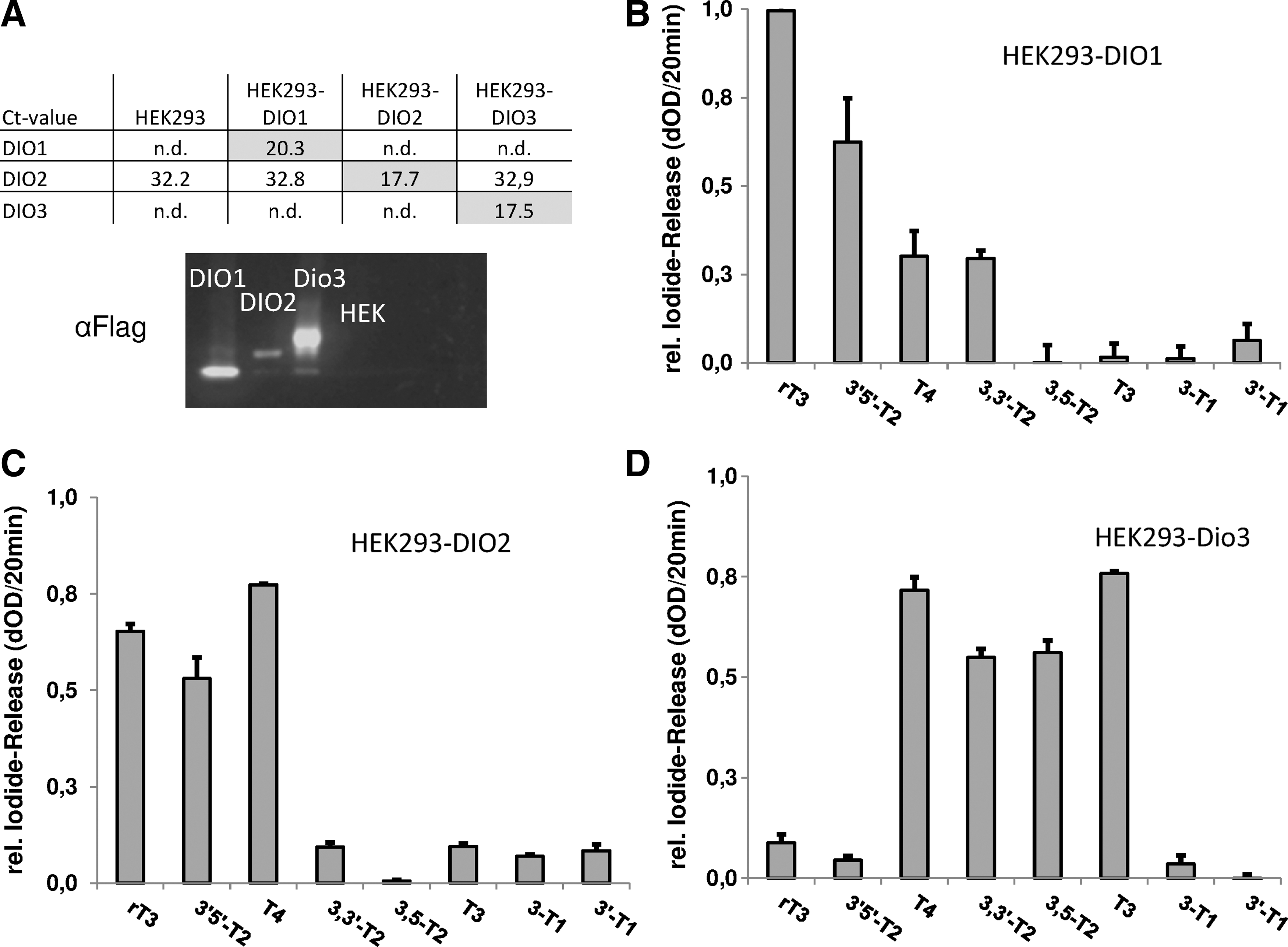
Overall, the values of the nonradioactive assay reliably reflect the patterns found in the LC-MS/MS measurements (Supplementary Fig. S1). While the LC-MS/MS analysis quantifies the disappearance of the substrates and sequential formation of products, the nonradioactive assay does not discriminate between primary and secondary deiodination steps, reflecting the total iodide released. This fundamental difference in assay setup likely explains the minor variations observed between these results.
Deiodination of iopanoic acid
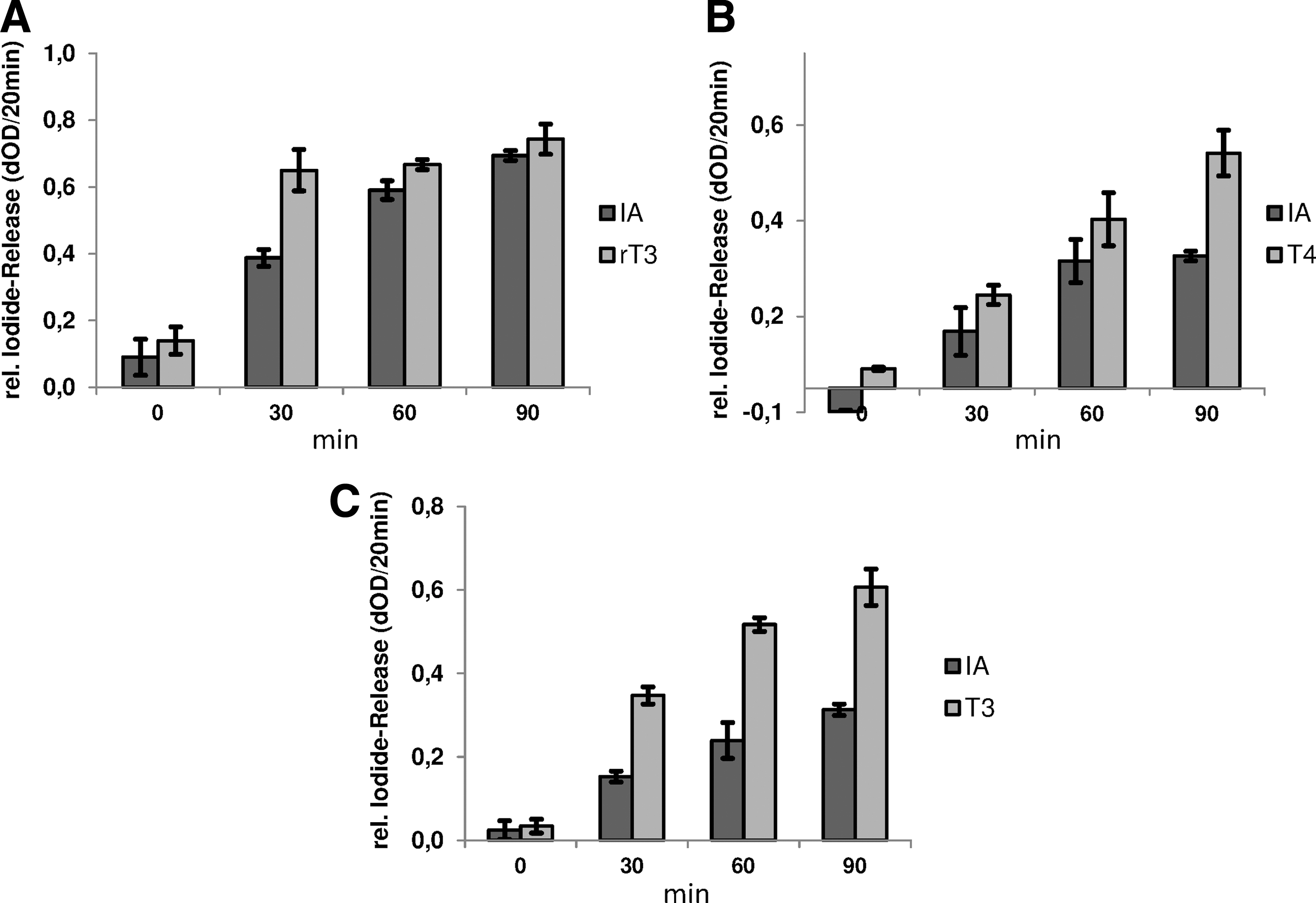
Incubation of the individual deiodinase isoenzymes with IA led to a time-dependent release of iodide within the reaction mixtures. This result is well in agreement with the data on the murine Dio1 shown before (9). However, the comparably strong deiodination activities observed with DIO2 and Dio3 are new (Fig. 2A–C) and qualify IA as a universal substrate for this enzyme family.
Screening of endocrine active substances as potential DIO inhibitors
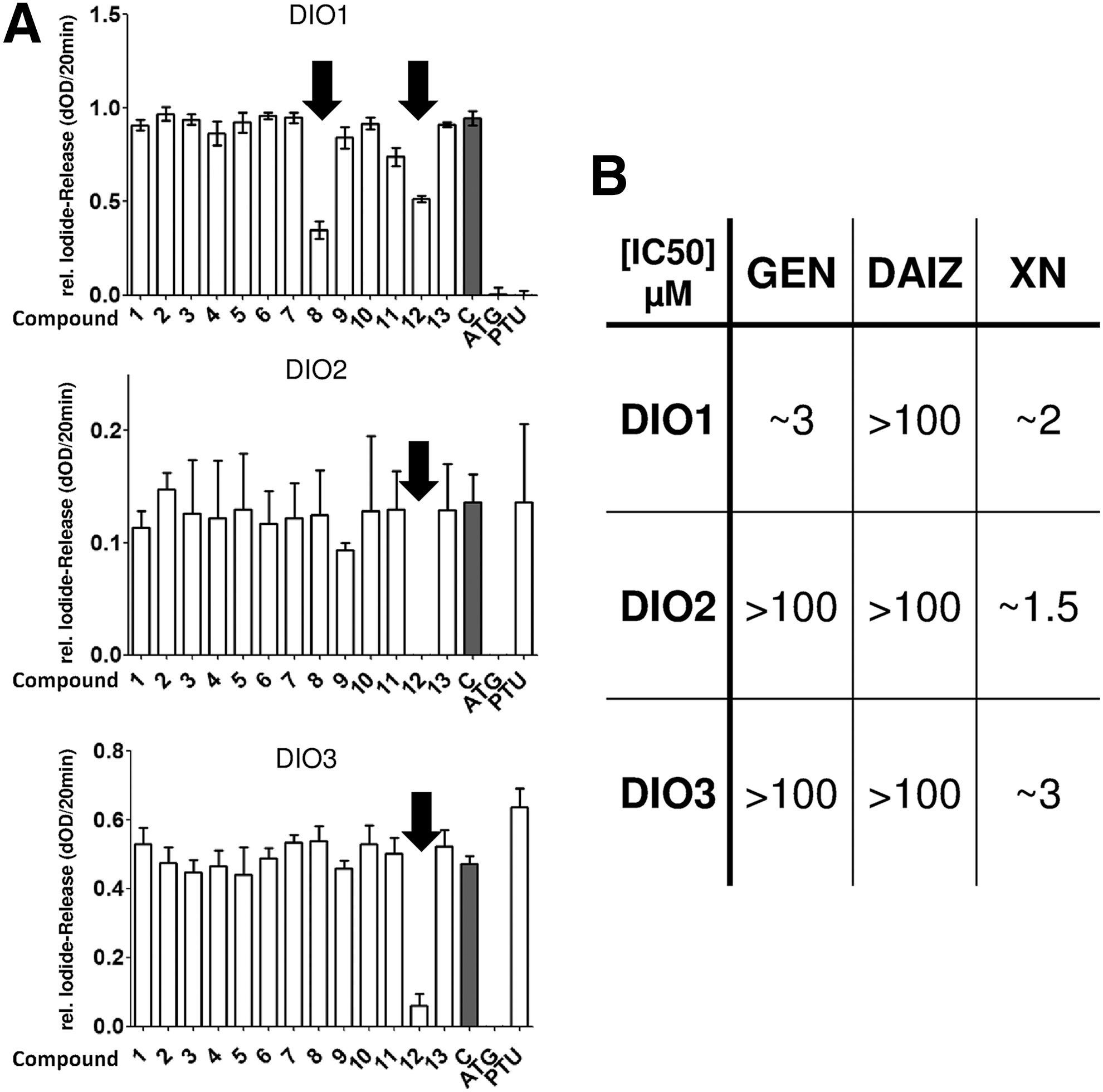
As a prood-of-concept experiment, two well-characterized DIO inhibitors ATG and PTU were chosen. As expected, PTU exclusively inhibited DIO1 activity without significantly affecting DIO2 or DIO3. In comparison, ATG suppressed all three DIO activities efficiently (Fig. 3).
Screening approach identifies genistein (GEN) and xanthohumol (XN) as potent inhibitors of DIO activity with different isoenzyme specificities.
For further identification of interacting molecules we used a collection of compounds (Table 1), which were recently identified as modulators of TH-receptor activation (15). In a first screening approach with all three DIO isoenzymes, GEN emerged as a compound that exclusively inhibits rT3-deiodination by DIO1. In contrast, XN turned out to be a nonselective but specific inhibitor of all three DIO isoenzymes (Fig. 3B, Supplementary Fig. S2).
To determine their relative potencies, dilution series of these compounds were tested in the DIO assays. The IC50 of GEN was found to be 3 μM, which is close to the value of PTU (∼1.5 μM). DIO2 and DIO3 activities were not blocked by GEN even in the highest concentrations tested.
Daidzein, as close relative to GEN in terms of structure, was included into the experimental setup. Notably, daidzein did not show any inhibitory potency under these conditions.
XN turned out to be a potent novel inhibitor for all three DIO. The experimentally determined IC50 of 1.5 to 3 μM is in the same range as observed for PTU or GEN with DIO1.
Discussion
Identification of and screening for isoenzyme-specific deiodinase inhibitors or modulators is of pharmacological and basic scientific interest, as they are discussed as key players in thyroid physiology. Moreover, recent studies indicate their potential involvement in tumorigenesis (8). PTU as a well-established inhibitor of DIO1 represents an important medication in hyperthyroidism and alternatives for PTU as therapeutics specifically targeting thyroid hormone activation are rare but urgently needed considering increased awareness of significant adverse effects of currently available antithyroid drug regimens (18,19).
In this technical report, we follow up on our recent work about a HTS-compatible method for Dio1 determination by refining and expanding the technique to DIO2 and DIO3, which are probably the more interesting targets for therapy and compound-screening efforts.
In order to do so, we first successfully established the recombinant expression of human DIO isoenzymes in HEK293 cells as a source of sufficient activities for the nonradioactive assays.
Using 5 μg to 100 μg of total protein homogenate per reaction was sufficient for achieving a well-detectable range of released iodide after 4 h of incubation with the preferred substrates, depending on the DIO isoenzyme (specific activities declined in the order of DIO1>DIO3>DIO2). The deiodination patterns for a collection of natural iodothyronines were consistent with the known substrate preferences from the literature (13,17). In addition, findings of the novel nonradioactive DIO assays were supported by educt and product analysis based on LC-MS/MS technology. We recently described IA as a substrate for murine Dio1 activity (9). As IA is described as a potential inhibitor of all three DIO isoenzymes, our results suggest that IA becomes accepted as a “nonselective” substrate by all DIOs. Its inhibitory activity is not inert, but rather involves iodide release which might be either of advantage in hyperthyroidism by blocking thyroid function via the Wolff–Chaikoff effect or even problematic in hyperthyroid patients already exposed to high iodine concentrations (e.g., by amiodarone or other iodine containing drugs) (20).
IA indeed turns out to be an interesting molecule to study DIO isoenzyme specificity. While it is a small molecule with just one phenolic ring and quite different from the TH in structure, IA becomes readily accepted by the active centers of all three DIO isoenzymes. Therefore, it might serve as a promising lead structure for new and specific competitive inhibitors.
For our screening approach, we selected thirteen synthetic or natural compounds (Table 1), depending on their proposed interaction with T3 receptor activation and signaling (15). Among synthetic receptor agonists or antagonists (GC-1, F21388, and NH3), most of them are suspected to be so-called endocrine disrupters and therefore in recent focus of concern in view of the relatively high prevalence rates of thyroid diseases and the globally increasing incidence of thyroid cancer (21,22).
GEN, a soy flavonoid, was found to inhibit DIO1 activity, which is consistent with earlier studies. It was shown to inhibit the endogenous Dio1 activity from FRTL-5 rat thyroid cells (23) and also to influence T4 to T3 conversion in vivo (24). We have now refined the picture by demonstrating that there is a direct influence of this compound on DIO1 activity. Notably, the isoenzymes DIO2 and 3 are unaffected by GEN under these conditions, reminding of a selective competitive genistein interaction with T4 binding in transthyretin but not with T4 binding sites of thyroxine-binding globulin or albumin (25).
In comparison, XN is a prenylated chalcone from hops and emerged as a potent inhibitor of all three DIO isoenzymes. Its inhibitory effects were observed in almost the same concentration range pointing to a complex effect on the thyroid hormone axis in vivo. Other studies have shown that hepatic expression of transcripts and activity of DIO1 is increased in XN-treated rats (26). In light of these new observations, this effect could indicate hepatic accumulation and impaired elimination of TH and their metabolites by direct inhibition of the DIO isoenzymes.
While the activities of DIO2 and DIO3 in tissues are relatively low compared with DIO1 (e.g., in hepatic samples) we are afraid that our method may not be sensitive enough for the analysis of physiological samples due to the limited sensitivity of the SK reaction. We were, for example, not able to detect Dio3 activity in murine placental tissue, even under optimal conditions and with long incubation times (14 h at 37°C; data not shown). For these purposes, alternative methods are indispensable.
The relatively high concentrations of substrates used in our measurements may be considered as another important limitation of our assay. However, in vitro enzyme assays are often performed under nonphysiological conditions to achieve reliable data (e.g., by using DTT as cofactor in the classical DIO enzyme assay with radioactive substrates). Moreover, the use of recombinant instead of endogenous enzymes is noteworthy. This has the goal to achieve isoenzyme specificity and to account for the fact that the relatively high enzyme activities needed for our HTS method is usually not found in tissues. Such limitations need to be considered when interpreting assay results, yet the data obtained and described with the known inhibitors of DIO activity are in line with expectations and argue for the suitability of our combination of recombinant enzymes with the SK reaction for inhibitor screening purposes.
In summary, by using human recombinant, enzymatically active DIO preparations, we have successfully established and improved nonradioactive versatile screening assays for all three deiodinase reactions, which allow identifying novel and potent global (e.g., xanthohumol) or isoenzyme-selective (e.g., genistein) DIO inhibitors in a cost-effective HTS format based on the detection of iodide release by the classical SK reaction.
Footnotes
Acknowledgments
The work was supported by the Bundesministerium für Bildung und Forschung Funding no. 0315370C (to J.K. and K.R.) and by the Deutsche Forschungsgemeinschaft (GraKo 1208, TP3 and TP6, Scho849/4-1; RE3038/1-1, Ho 5096/1-1, SCHW914/3-1).
Author Disclosure Statement
No competing financial interests exist.