
Abstract
Background:
Thyroid iodide uptake, mediated by the sodium–iodide symporter (NIS), is essential for thyroid hormone synthesis and also for treatment of thyroid diseases, such as thyroid cancer, through radioiodine therapy. Therefore, compounds able to increase thyroid iodide uptake could be clinically useful, and it is of great importance to unravel the mechanisms underlying such an effect. It has been shown previously that the flavonoid rutin increases thyroid radioiodide uptake in vivo in rats. This study aimed to investigate the mechanisms involved in the stimulatory effect of rutin on iodide uptake.
Methods:
This study evaluated iodide uptake, NIS expression and its subcellular distribution, iodide efflux, reactive oxygen species levels, and the intracellular pathways involved in NIS regulation in a rat thyroid PCCL3 cell line treated with rutin.
Results:
Similar to previous results found in vivo, rutin increased radioiodide uptake in PCCL3 cells, which was accompanied by increased NIS expression (at both the mRNA and protein levels) and a reduction of radioiodide efflux. Moreover, the results suggest that rutin could regulate NIS subcellular distribution, leading to higher levels of NIS at the cell membrane. In addition, rutin decreased the levels of intracellular reactive oxygen species and phospho-5'-adenosine monophosphate-activated protein kinase.
Conclusions:
The flavonoid rutin seems to be an important stimulator of radioiodide uptake, acting at multiple levels, an effect that can be due to decreased oxidative stress, reduced 5′-adenosine monophosphate-activated protein kinase activation, or both. Since thyroid iodide uptake is crucial for effective radioiodine therapy, the results suggest that rutin could be useful as an adjuvant in radioiodine therapy.
Introduction
I
Thyrotropin (TSH) is the main regulator of thyroid cell differentiation, proliferation, and function, including iodide uptake (7). TSH acts via a basolateral surface receptor (TSHr), which is a G protein-coupled receptor, leading predominantly to the activation of adenylyl cyclase, cyclic monophosphate adenosine (cAMP) accumulation, and thus activation of protein kinase A (PKA) (8). Classically, TSH increases iodide accumulation by positively regulating NIS expression at protein and mRNA levels via the PKA/cAMP/CREB pathway (9). In addition, TSH signaling is required for NIS targeting to, and/or retention at, the basolateral membrane (10).
TSH can also modulate NIS by additional pathways, such as extracellular signal-regulated kinase/mitogen-activated protein kinase (ERK/MAPK) and p38MAPK (11). However, the actual role of these kinases on NIS function and expression is still controversial (11 –13). Besides, it has been shown that the inhibition of the phosphatidylinositol-3-kinase (PI3K) pathway in thyroid cells increases iodide uptake through upregulation of NIS expression (14 –16). Moreover, 5′-adenosine monophosphate-activated protein kinase (AMPK) acts as a suppressor of iodide uptake in thyroid cells, and TSH reduces AMPK phosphorylation/activation, increasing NIS expression (17,18).
The consumption of flavonoids, polyhydroxy polyphenols found in vegetables of normal human diet, has been associated with thyroid dysfunction (19 –21). In 2008, Giuliani et al. showed that the flavonoid quercetin could inhibit the PI3K/Akt pathway, downregulating iodide uptake and NIS mRNA levels in the FRTL5 cell line (22). Moreover, it has been shown that incubation of FTC-133 cells transfected with NIS with the flavonoid myricetin for four days could increase intracellular iodide content, even though most flavonoids tested decreased iodide uptake in this cell line. The stimulatory effect of myricetin was attributed to both increased iodide uptake and decreased iodide efflux. However, its mechanism of action was not determined (23).
Rutin (5,7,3′,4′-OH,3-rutinose), the glycosidic form of quercetin, is a flavonol widely consumed as part of a common diet and because of its pharmacological properties, including vascular-protective, antitumor, antiviral, antioxidant, anti-inflammatory, myocardial protective, and anti-diabetic effects (24 –28). It has recently been shown that among the flavonoids tested, rutin was the only one able to increase thyroid iodide uptake significantly in vivo, besides increasing NIS mRNA and protein levels in rats. Stimulation of NIS was not due to TSH, which remained unchanged, thus suggesting that rutin could directly modulate NIS expression (29).
Since NIS remains active in almost all differentiated thyroid carcinomas, radioiodine therapy is frequently used after total thyroidectomy, aiming at the ablation of the remaining thyroid tissue (30,31). Nevertheless, in a subset of patients, NIS expression decreases, thus rendering radioiodine therapy ineffective and reducing therapeutic options and survival. It has previously been shown that the flavonoid rutin is able to increase thyroid iodide uptake in vivo. Therefore, the objective of this study was to study the mechanisms involved in the stimulatory effect of rutin using the well-characterized PCCL3 rat thyroid cell line. A better mechanistic understanding of the effect of rutin might be useful for the development of new therapeutic strategies to improve the efficacy of radioiodine therapy in thyroid cancer.
Materials and Methods
Cell culture
PCCL3 cells (donated by Prof. Roberto Di Lauro, Stazione Zoologica Anton Dohrn) were maintained in Coon's modified Ham's F-12 medium, supplemented with 5% fetal bovine serum and a six-hormone mixture (1 mIU/mL of TSH, 10 mg/mL of insulin, 5 mg/mL of transferrin, 10 nM of hydrocortisone, 10 ng/mL of somatostatin, and 10 ng/mL of glycyl-L-histidyl-L-lysine acetate; complete medium). PCCL3 cells were incubated with rutin (Sigma–Aldrich, St. Louis, MO), dissolved in dimethyl sulfoxide (DMSO; 0.1%) at different concentrations ranging from 0 to 100 μM for 24 and 48 h, and cell viability was evaluated in order to test if rutin is toxic to these cells. Control cells (without rutin) were cultivated in the presence of vehicle (0.1% [v/v] DMSO).
To evaluate the participation of ROS and AMPK in the effect of rutin, PCCL3 cells were treated for 24 h with 25 μM of rutin in the presence or absence of 25 μM of H2O2 and in the presence or absence of 500 μM of AICAR, a pharmacological activator of AMPK. The incubation with H2O2 was for 1 h prior to performing the iodide uptake assay, while AICAR treatment was performed for the same period of treatment with rutin (i.e., 24 h).
Cell viability
As an index of cell viability, the commercially available MTT assay was used (Sigma–Aldrich), according to the manufacturer's recommendations. This is a colorimetric assay determining the number of viable cells. The assay is based on the cellular conversion of the tetrazolium salt into formazan that is soluble in cell culture medium and is measured directly at 490 nm in a 96-well plate. Absorbance is directly proportional to the number of living cells. PCCL3 cells were seeded and cultured, as described above. After treatment, cells were incubated with MTT (0.5 mg/mL) for 3 h at 37°C in a humidified 5% CO2 atmosphere. Then, cells were lysed with DMSO (P.A.). All determinations were done in triplicate.
Iodide uptake assay
Iodide uptake assay was performed, as previously described (32,33). Briefly, 105 PCCL3 cells were grown in a 24-well plate in different assay conditions (0–100 μM of rutin for 24 and 48 h). Then, cells were incubated for 45 min at 37°C with 1 mL of Hank's balanced salt solution (HBSS) containing ∼0.1 μCi of carrier-free Na125I (PerkinElmer Life and Analytical Sciences, Woodbridge, Canada) and 100 μM of NaI. For each experimental condition, some wells received 10 mM of KClO4, a competitive NIS inhibitor, in order to determine the nonspecific radioiodide uptake. After incubation, cells were washed once with ice-cold HBSS and lysed with 0.1 M of NaOH, and the radioactivity was measured in a gamma-counter. Specific iodide uptake was obtained by subtracting iodide uptake in the absence and presence of KClO4 and related to protein concentration, measured by the Bradford method (34). Results are expressed as specific units of iodide accumulation relative to control. Since a significant increase was found in iodide uptake with 25 μM of rutin treatment for 24 h, subsequent measurements were done using this time and concentration.
Immunocytochemistry
In order to evaluate whether rutin could affect subcellular localization of NIS, immunocytochemistry of NIS was performed. Thus, cells were plated on glass coverslips, previously positioned in 24-well plates. After treatment, cells were washed twice with 0.01 M of phosphate-buffered saline (PBS), pH 7.4, fixed with 4% paraformaldehyde (Sigma–Aldrich) at room temperature for 15 min and then washed again with PBS. After washing, cells were incubated with 50 nM of NH4Cl in PBS for 30 min and then incubated for 30 min with a solution of 5% bovine serum albumin, 0.1% gelatin, 0.1% Triton X-100, 0.05% Tween®20 (Sigma–Aldrich) in PBS for 30 min. After this step, PCCL3 cells were incubated overnight with a rabbit anti-NIS polyclonal antibody in a humidified chamber at 4°C (anti-NIS antibody was kindly provided by Dr. Nancy Carrasco, Department of Cellular and Molecular Physiology, Yale University School of Medicine, New Haven, CT). Subsequently, cells were incubated with a fluorescence-conjugated secondary antibody (goat anti-rabbit IgG Alexa 488; Invitrogen, Carlsbad, CA) for 2 h at room temperature. Coverslips were mounted with N-propyl gallate (Sigma–Aldrich) and observed using a Zeiss Axiovert 100 microscope (Carl Zeiss AG, Oberkochen, Germany). DAPI (4′,6-diamidino-2-phenylindole dilactate; Sigma–Aldrich) was used for nuclear staining. Negative control was done omitting the primary antibody. The images (63/1.4 oil objective lens) were taken using a Leica TCS SP5 confocal microscope (Leica Camera AG, Wetzlar, Germany) using LAS AF software. All images were processed using Image J v1.45s.
Iodide efflux assay
The procedure was performed, as previously described (23,33,35). Briefly, 105 PCCL3 cells were seeded in 24-well plate, and on the subsequent day, cells were incubated with or without 25 μM of rutin for 24 h. After treatment, cells were washed twice with HBSS and then incubated with 1 mL of HBSS containing 0.1 μCi of carrier-free Na125I and 100 μM of NaI for 45 min. Then, the medium was removed and replaced with fresh HBSS without NaI. This procedure was repeated every 3 min until 12 min had elapsed. The content of 125I in the collected supernatant was measured in a gamma-counter. After the last time point (12 min), cells were lysed with 0.1 M of NaOH to measure the residual intracellular radioactivity. An aliquot of cell lysates was used to measure protein concentration, and the results were expressed as specific units of iodide accumulation relative to initial radioactivity. In order to compare the capacity of iodide retention better, iodide uptake at time zero was considered as 100% in both conditions (with and without rutin).
Immunoblotting
After 24 h of treatment with 25 μM of rutin or vehicle, PCCL3 cells were homogenized in lysis buffer containing 135 mM of NaCl, 1 mM of MgCl2, 2.7 mM of KCl, 20 mM of Tris, pH 8.0, 1% Triton, 10% glycerol, and protease and phosphatase inhibitor cocktails (Sigma–Aldrich). An aliquot was used to determine protein concentration by BCA protein assay kit (cat. no. 23 227; Pierce, Rockford, IL), according to the manufacturer's recommendations. Samples were then subjected to SDS/PAGE electrophoresis, transferred to polyvinylidene fluoride membranes, and probed with the indicated antibodies. The NIS antibody was kindly provided by Dr. Nancy Carrasco. TSHr, phospho (Tyr 204), and total-ERK primary antibodies were purchased from Santa Cruz Biotechnology (Dallas, TX), and phospho (Ser133), total-CREB, phospho (Thr172), and total-AMPKα were from Cell Signaling Technology (Danvers, MA). For loading control, GAPDH (Sigma–Aldrich) and alfa/beta-tubulin (Cell Signaling Technology) antibodies were used. Then, membranes were incubated with the appropriate secondary antibodies. The detection of the proteins was performed using ECL (Millipore, Billerica, MA).
Real-time polymerase chain reaction
PCCL3 cells were seeded in six-well plates and treated as described above. Subsequently, medium was removed, and cells were washed twice with PBS. Trizol® Reagent (300 μL; Life Technologies™, Carlsbad, CA) was added to the wells. Extraction procedures were performed according to manufacturer's instructions. In order to quantify total RNA concentration of the samples and to test for integrity and purity, the absorbance of extracted RNA was measured at the wavelengths 260 and 280 nm in a spectrophotometer Nano Vue Plus (GE Healthcare Life Sciences, Little Chalfont, United Kingdom). The relation between the absorbances was used as an indication of purity, and only samples with a relation between 1.5 and 2 were included. cDNA synthesis reaction was performed using a High Capacity Reverse Transcriptase kit (Applied Biosystems™, Carlsbad, CA) using 1 μg of RNA, following the manufacturer's instructions. In the real-time polymerase chain reaction (RT-PCR) assay, cDNAs were amplified in the presence of specific oligonucleotides, both forward and reverse, as described in Table 1. Amplifications were performed in a thermocycler ABI Prism 7500 (Applied Biosystems™) in 96-well plates. The final reaction volume was 12 μL/well (3 μL of sample cDNA diluted 1:30, 0.25 μL of 10 nM forward and reverse oligonucleotides, 6 μL of Power SYBR Master MIX from Applied Biosystems™, and 2.5 μL RNase-free water). The amplification program used was: 50°C for 2 min, 95°C for 10 min, followed by 40 cycles at 95°C for 10 s and 60°C for 1 min. RT-PCR efficiency was assessed using serial dilutions of the template cDNA, and melting curve data were collected to check RT-PCR specificity. Every cDNA was amplified in triplicate, and a corresponding sample without reverse transcriptase (no-RT sample) was included as negative control. The β-actin gene was used as internal control of amplification, and the content of the target transcripts were normalized to that of β-actin. The relative amount of target gene expression was determined using the comparative CT method, expressed by the formula 2–(ΔΔCT) (36), in which CT refers to threshold cycle and is determined for each well by the 7500 Real-Time PCR System Sequence Detection Software (Applied Biosystems™). ΔCT is the difference between the CT of the target mRNA and the CT of β-actin. Results are expressed relative to controls (vehicle-treated cells).
NIS, sodium–iodide symporter; TSHr, thyrotropin receptor.
Intracellular reactive oxygen species
Intracellular reactive oxygen species (ROS) was measured using a dihydrodichlorofluorescein diacetate (DCFH-DA) probe (37). Cells were plated (104 cells/well) in 96-well white-colored plates and treated as described above. Later, cells were washed with D-PBS buffer and then incubated with 10 μM DCFH-DA at 37°C for 30 min. After that, fluorescence was determined at 485/530 nm excitation/emission using a microplate reader (Victor X4; PerkinElmer). The resulting fluorescence is expressed relative to controls.
Results
Effect of rutin on cell viability and iodide uptake
Initially, the influence of rutin on PCCL3 cell viability was investigated. The data demonstrate that treatment with rutin does not seem to be toxic to PCCL3 cells in the tested conditions (Fig. 1A and B), even at the highest concentration and time of exposure (100 μM of rutin, 48 h; Fig. 1B).
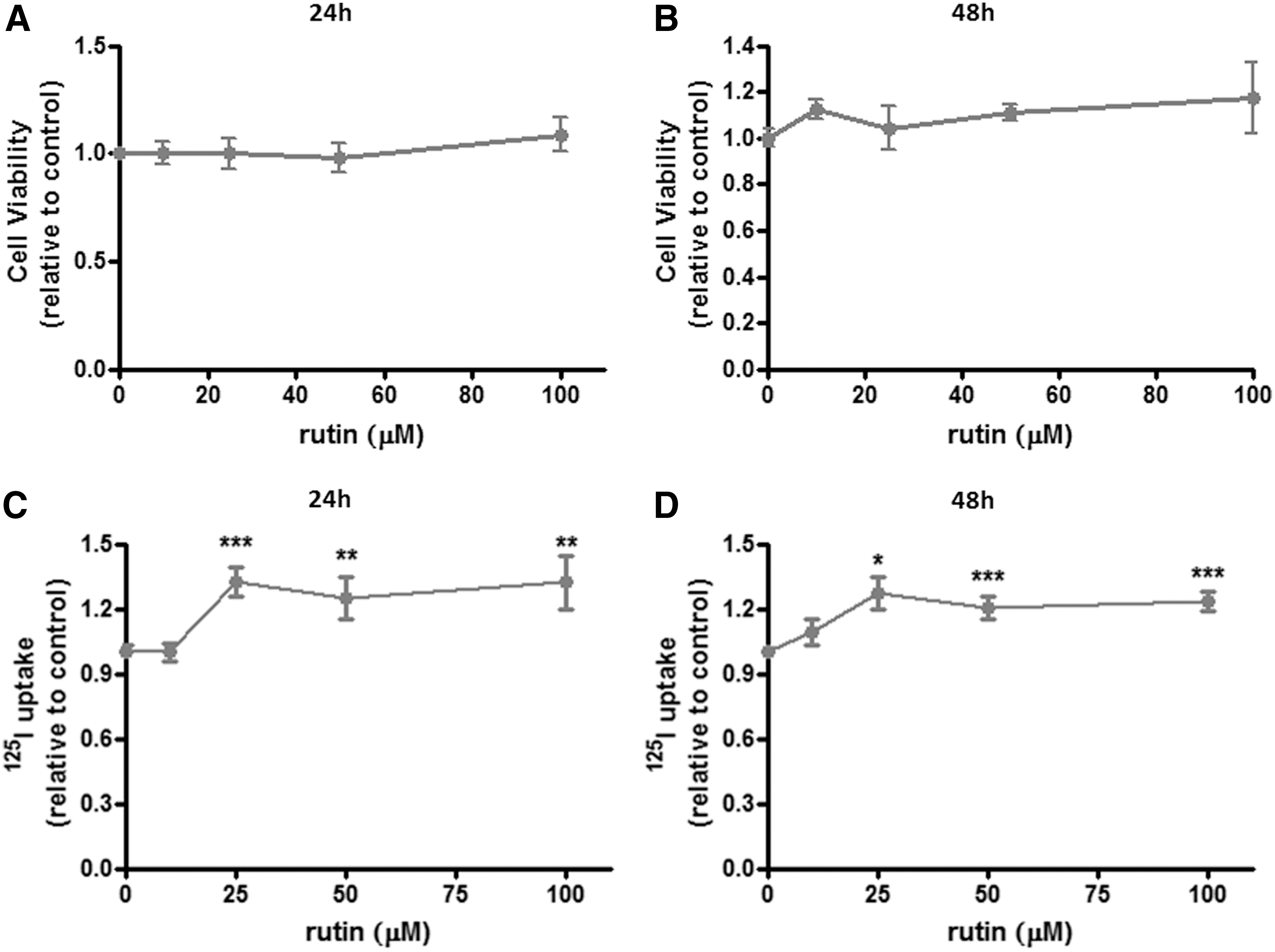
Cell viability and radioiodide uptake in rutin-treated PCCL3 cells. Rat thyroid PCCL3 cells were treated with the flavonoid rutin at concentrations of 0, 10, 25, 50, and 100 μM for 24 and 48 h, and cell viability (
Next, the study evaluated if rutin treatment could modulate PCCL3 radioiodide uptake, as was previously found for the rat thyroid in vivo (29). As shown in Figure 1C and D, iodide uptake was significantly increased by exposure to rutin in concentrations ranging from 25 to 100 μM after both 24 and 48 h of treatment. Therefore, the lowest concentration/time condition able to increase radioiodide uptake (i.e., 25 μM of rutin, 24 h) was used in the subsequent experiments.
Effect of rutin on NIS expression
In order to evaluate whether rutin regulates NIS at transcriptional and/or translational levels, NIS mRNA and protein expressions were evaluated. In fact, rutin treatment was able to upregulate NIS protein (Fig. 2A) and mRNA (Fig. 2B) levels, besides increasing iodide uptake (Fig. 2C), thus suggesting that the stimulatory effect of this flavonoid on NIS function is due to an increment in NIS expression and not related to a direct interaction of rutin with the protein. Additionally, immunofluorescence data demonstrate that NIS staining is more pronounced at the plasma membrane in rutin-treated cells (yellow arrows), while control cells show a pronounced staining near the nucleus (white arrows; Fig. 3). Moreover, as shown in Supplementary Figure S1 (Supplementary Data are available online at
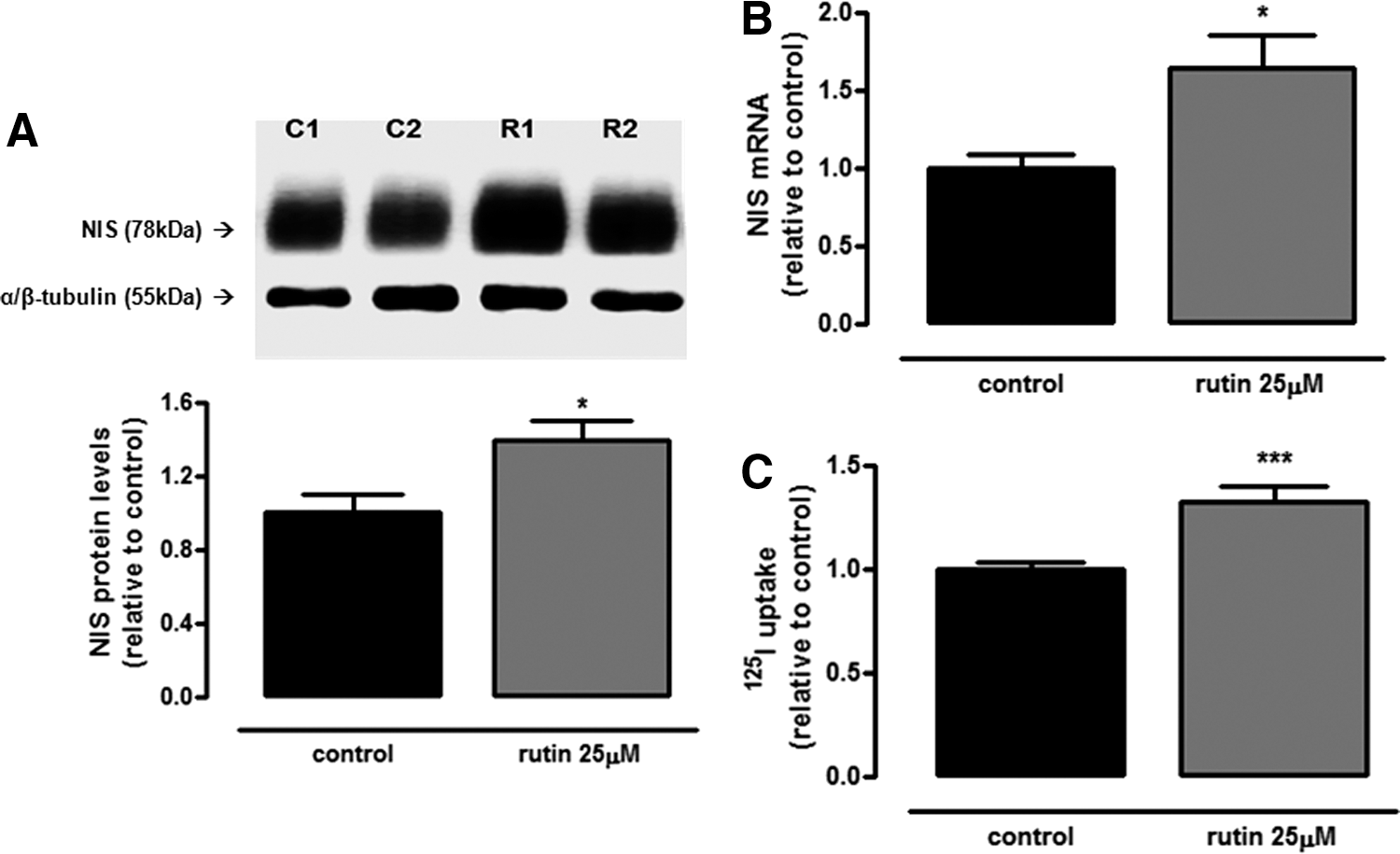
Rutin stimulates sodium–iodide symporter (NIS) expression and radioiodide uptake. (

The subcellular distribution of NIS is regulated by rutin. PCCL3 cells, treated or not with 25 μM of rutin for 24 h, were stained for NIS and nuclei (DAPI). Under control conditions (upper panels), there is marked staining for NIS in perinuclear compartments (arrows, top-right panel), while rutin-treated cells show a strong staining for NIS in the plasma membrane (arrows, bottom right). Bars = 20 μm (bar). In the control condition, cells were incubated with vehicle at the same concentration as in rutin-treated conditions (0.1% v/v DMSO).
Effect of rutin on radioiodide efflux
The intracellular iodide concentration depends not only on iodide uptake but also on iodide efflux. Thus, next the study investigated if the increase in intracellular iodide content observed in rutin-treated cells could also be related to a decrease in iodide efflux. As shown in Figure 4A, incubation of PCCL3 cells with rutin increased the amount of radioiodide remaining in thyrocytes, besides augmenting the area under the curve of cellular radioiodide retention (Fig. 4B). However, treatment with rutin did not change mRNA levels of pendrin or anoctamin-1 (Fig. 4C), thus suggesting that rutin does not affect iodide efflux by transcriptional regulation of these anion channels. On aggregate, the results suggest that rutin increases PCCL3 ability to concentrate iodide intracellularly due to both an increase in iodide uptake and a decrease in iodide efflux, the mechanism of which remains unclear.
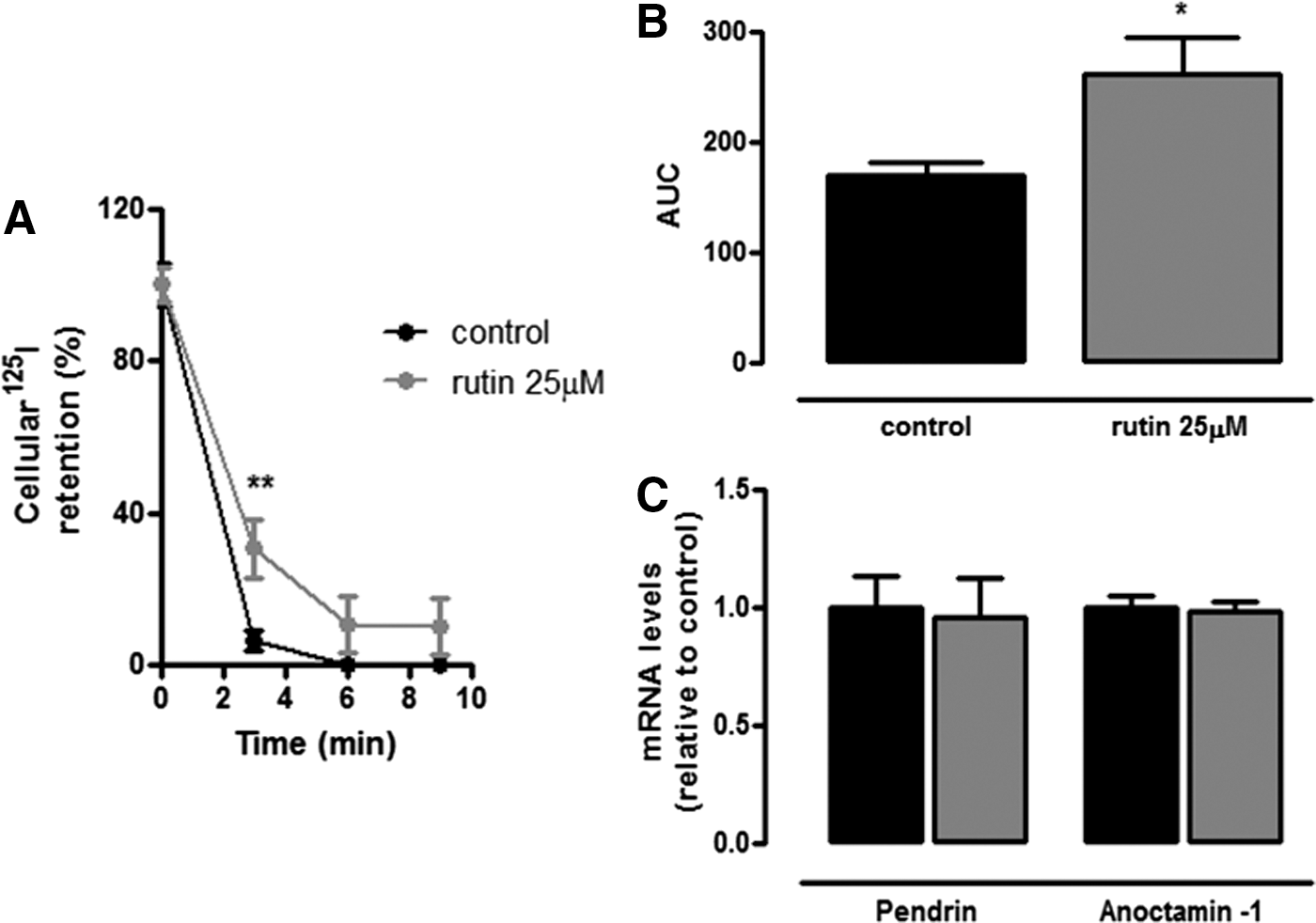
Effect of rutin on radioiodide retention and mRNA levels of pendrin and anoctamin-1. (
Mechanism of NIS regulation by rutin
Since TSH is the main regulator of thyroid function (6), the study evaluated whether rutin treatment could affect TSHr mRNA and/or protein levels, besides phosphorylated and total CREB levels, in order to clarify the mechanism through which rutin regulates NIS expression and function in PCCL3 cells. As shown in Figure 5A and B, treatment with rutin did not modulate TSHr expression at the mRNA or protein level. Accordingly, treatment with rutin did not change CREB phosphorylation or total CREB levels, as shown in Figure 5C.
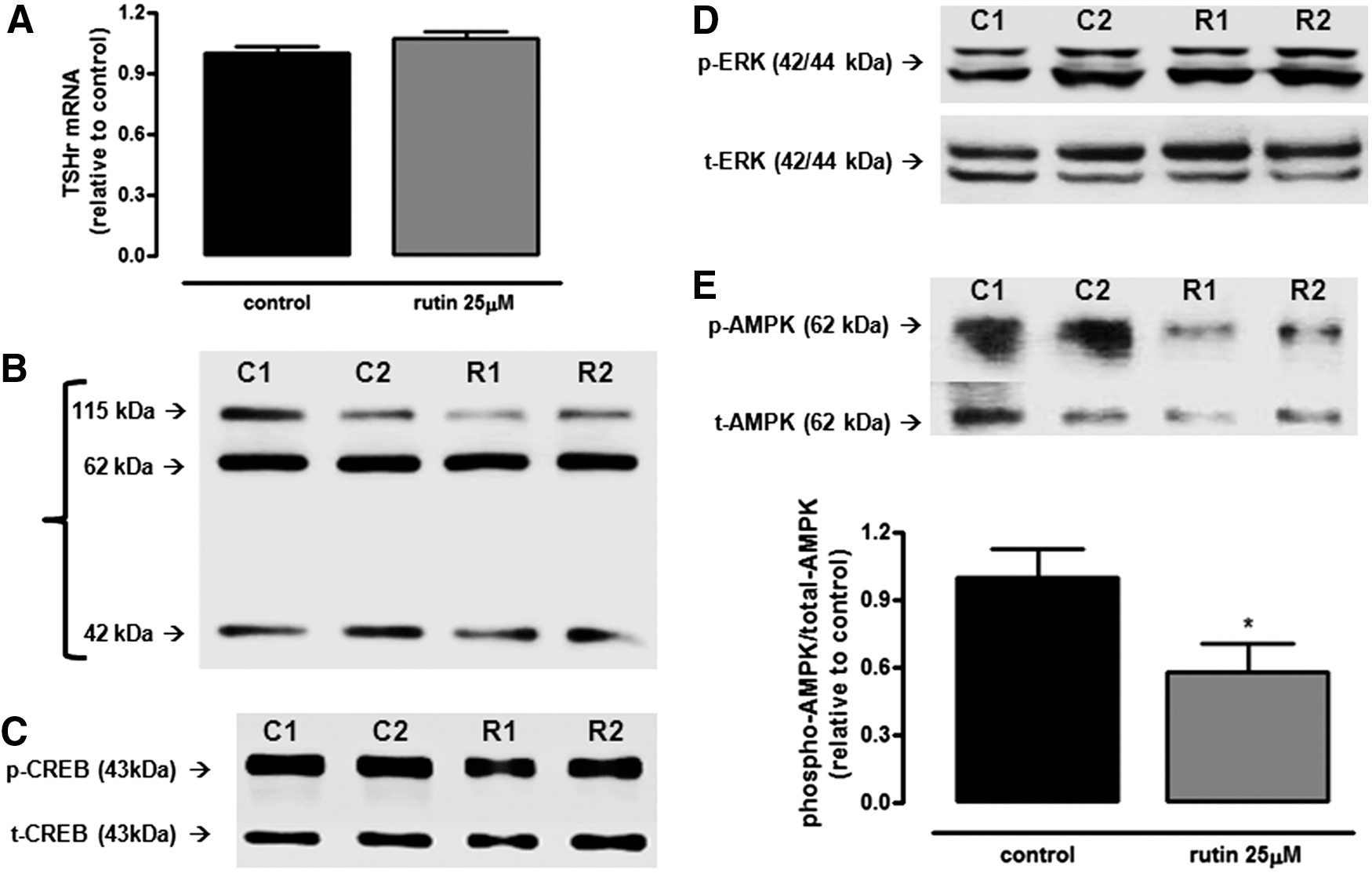
Expression of proteins involved in NIS regulation in rutin-treated PCCL3. Thyrotropin receptor mRNA (
Since treatment with rutin does not seem to modulate the classical TSHr–PKA–CREB pathway, next, the effect of rutin treatment on MAPK/ERK signaling, which has also been shown to modulate NIS (12,13), was evaluated. However, the MAPK/ERK pathway does not seem to be involved in the rutin effect either, since phospho-ERK and total-ERK levels remained unchanged (Fig. 5D).
AMPK has been shown to be an important regulator of NIS function and expression both in vivo and in vitro (17,18). Therefore, the effect of rutin treatment on AMPK phosphorylation levels was evaluated. In fact, as shown in Figure 5E, the treatment with rutin significantly decreased the relation between phospho-AMPK and total-AMPK protein levels when compared to control. This result suggests that the inactivation of AMPK by rutin treatment might be involved in the positive effect of this flavonoid on iodide uptake.
It has been shown previously that the increase of ROS levels in thyrocytes can inhibit NIS function (38). Additionally, rutin has been described as a potent ROS scavenger (39). Thus, it was speculated that rutin could exert antioxidant activity in PCCL3 cells, which could contribute to the increment of iodide uptake by reducing ROS levels. In fact, PCCL3 incubation with rutin significantly decreased intracellular ROS concentrations (Fig. 6A).

Antioxidant effect of rutin on PCCL3 cells and involvement of reactive oxygen species (ROS) and 5′-adenosine monophosphate-activated protein kinase (AMPK) on rutin action in iodide uptake. (
In order to confirm that the effect of rutin involves both ROS reduction and AMPK inhibition, rutin was associated with H2O2 and with AICAR, a pharmacological inhibitor of AMPK. As expected, H2O2 incubation reduced iodide uptake, an effect reversed by incubation with rutin (Fig. 6B). Moreover, AMPK activation by AICAR reduced iodide uptake, as previously described (17), and this reduction was prevented by addition of rutin in the medium.
Therefore, the reduction of ROS levels and AMPK activity might contribute to the increment in iodide uptake produced by rutin in PCCL3 cells.
Discussion
Besides playing a key role in thyroid hormone synthesis, iodide uptake is also important for diagnosis and treatment of some thyroid diseases, including thyroid cancer. Since the development of radioiodine therapy, it has been one of the pillars in the treatment of differentiated thyroid cancer (30,31). Usually, radioiodine is administered after thyroidectomy, aiming at the complete ablation of the remaining thyroid tissue (40). However, NIS function is decreased during tumor progression in some thyroid cancer patients, thus reducing the efficacy of radioiodine therapy (41). Therefore, it is of great importance to find new compounds able to increase iodide uptake, besides understanding the mechanisms involved.
According to the present results, the flavonoid rutin shows a consistent effect enhancing NIS expression and function. It was observed that incubation of PCCL3 cells with rutin from 25 to 100 μM for 24 and 48 h was able to increase radioiodine uptake. Besides the increase in radioiodine uptake, the treatment with rutin was able to upregulate NIS at the mRNA and protein levels. These results are in accordance with a prior in vivo study, in which it was shown that treatment with rutin increased NIS expression and function in rat thyroids (29), suggesting that the stimulatory effect of rutin on NIS is consistent among different experimental models. Even though the increase in radioiodide uptake by rutin is about 20%, it is important to underline that the experiment was done in the presence of TSH, the main stimulator of NIS. Thus, iodide uptake was already greatly stimulated by TSH, and rutin was able to stimulate iodide uptake additionally. In addition, in the present work, the effect of the flavonoid rutin on NIS subcellular localization was studied. PCCL3 cells treated with rutin showed higher NIS expression in the plasma membrane when compared to control, suggesting that rutin modulates NIS subcellular distribution. Thus, it is possible that the increased iodide uptake induced by rutin is not only due to the stimulation of NIS expression, but also to the increased translocation of the symporter to the plasma membrane in PCCL3 cells.
The effect of flavonoids on NIS function and expression is controversial, according to the literature. One of the pioneer works about the effect of flavonoids on NIS in vitro showed that most of the studied flavonoids decreased NIS function and, among the tested flavonoids, only myricetin was able to increase radioiodide uptake in FTC-133 cells transfected with NIS (23). Recently, it was demonstrated that the flavonoid quercetin decreases radioiodide uptake and NIS expression in vitro (22) and in vivo (42). Therefore, quercetin and rutin, despite their structural similarities, seem to have opposite effects on NIS. Later, it was demonstrated that the incubation with the flavonoid apigenin increased radioiodine uptake in PCCL3 cells with inducible expression of BRAFV600E or RET/PTC3 without modulation of NIS expression, and this effect was dependent on previous inhibition of the Akt pathway (43,44).
In addition to the stimulation of NIS function and expression, it was found that rutin also decreases radioiodide efflux from PCCL3 cells, although without changes in pendrin or anoctamin-1 mRNA levels. Data from the literature show that most of the tested flavonoids do not modulate iodide efflux, irrespective of the thyroid cell line tested (23,43,44). However, it was demonstrated that myricetin, besides stimulating iodide uptake, was also able to decrease iodide efflux from FTC-133 cells, indicating that this flavonoid could be useful to increase radioiodine retention (23). Recently, Lakshmanan et al. (44) showed that despite the stimulation of iodide uptake, treatment with apigenin did not modulate pendrin or anoctamin-1 mRNA levels, similar to the results found in the present study. Hence, the data suggest that treatment with rutin could increase intracellular iodide storage ability of PCCL3 due to the stimulus of NIS expression, increased NIS translocation into the cell plasma membrane, and reduced iodide efflux.
Activation of the PKA/CREB pathway by TSH is the main stimulus for NIS function and expression (9,11). Moreover, the MAPK/ERK and p38MAPK pathways have been described to be able to downregulate NIS function/expression, even though the results are controversial (12,13). In the present study, treatment of PCCL3 cells with rutin did not affect TSHr expression or phosphorylation of CREB and ERK. These results suggest the participation of other intracellular effector(s) in the action of rutin.
In 2011, the effect of AMPK, an important sensor of cellular energy levels, on NIS regulation was shown both in vivo and in vitro (17). The authors showed that the stimulation of AMPK by AICAR, a pharmacological activator, led to a decrease in radioiodide uptake in PCCL3. Conversely, when these cells were incubated in the presence of an AMPK inhibitor, compound C, NIS function was increased in parallel with decreased phosphorylated AMPK levels (17). Subsequently, it was proposed by another group that the increase in NIS function was probably due to CREB activation in PCCL3 cells after 96 h of incubation with compound C (45). Still in 2014, it was suggested that inhibition of NIS function and expression by activated AMPK was a consequence of the stimulus of lysosomal degradation of NIS, since the incubation of PCCL3 with bafilomyicin, a lysosomal degradation inhibitor, prevented these effects (18). Therefore, it was conjectured that rutin could stimulate NIS function and expression by decreasing AMPK activation. In fact, a decrease in the ratio between phosphorylated and total AMPK was observed in rutin-treated PCCL3 cells, and the co-treatment of rutin with AICAR reversed the inhibitory effect of this drug on iodide uptake. Thus, the data suggest that the increase in NIS function could occur, at least in part, due to reduced AMPK activation in the presence of rutin.
Rutin seems to be beneficial in the treatment of diseases in which the redox homeostasis is affected due to its important ROS scavenger effect (46,47). Since thyrocytes physiologically produce high amounts of H2O2, which is essential for thyroid hormone synthesis, and since the increase in ROS levels has been shown to reduce iodide uptake in thyrocytes (38), it was speculated that rutin could have antioxidant activity in the present model, which could contribute to the increase in NIS function. Indeed, a reduction of intracellular ROS levels was found in rutin-treated cells, thus reinforcing the idea that rutin has an important antioxidant effect. Moreover, rutin was capable of preventing the inhibitory effect of H2O2 on iodide uptake. These results are in accordance with previous studies showing an antioxidant effect of rutin in other experimental models (39,48). Li et al. (39) showed that rutin could reduce ROS production due to a decrease in NOX4 expression. Since previous studies have shown that ROS could inhibit NIS function and expression (38,49), the opposite may also be true, that is, a decrease in ROS availability might produce an enhancement of iodide uptake. Hence, the antioxidant effect of rutin could contribute to the stimulatory effect of rutin on NIS. It is worth mentioning that it has been shown that ROS activates AMPK in other experimental models (50), which could in turn inhibit NIS. Therefore, the reduction of intracellular reactive species observed in rutin-treated cells could contribute to the decreased AMPK activation and thus to NIS stimulation.
In summary, according to the present results, the stimulatory effect of rutin on iodide uptake and NIS expression might be due to both its antioxidant effect and its inhibitory effect on AMPK. Moreover, the reduction of AMPK activation by rutin can be related to the antioxidant effect of this flavonoid. It is important to underline that rutin was shown to be not only a potent stimulator of radioiodide uptake, but also an inhibitor of radioiodide efflux, thus leading to an increased radioiodide content in the thyrocyte. This ability is fundamental to radioiodine therapy efficiency, and it is tempting to speculate that the flavonoid rutin could be useful as an adjuvant to radioiodine therapy.
In conclusion, treatment with rutin was able to increase radioiodide content significantly in PCCL3 cells by enhancing NIS function and expression and by decreasing iodide efflux. The mechanisms underlying these effects seem to involve both antioxidant properties and the pAMPK inhibitory effect of rutin.
Footnotes
Acknowledgments
We are grateful for the technical assistance of Norma Lima de Araújo Faria, Wagner Nunes Bezerra, and José Humberto Tavares de Abreu. This work was supported by Pro Onco Vivi Nabuco-Fundação do Câncer, Fundação de Amparo à Pesquisa do Rio de Janeiro (FAPERJ), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Author Disclosure Statement
No competing financial interests exist.