
Abstract
Background:
Seasonal changes in various physiological events have been reported in humans, including metabolism, immune function, and mood. However, the molecular and endocrine basis of these seasonal changes remains unclear.
Summary:
Animals that breed seasonally, such as Japanese quail and the Siberian hamster, have sophisticated seasonal mechanisms, and hence they provide excellent opportunities to understand the underlying processes. Functional genomic analysis in quail uncovered the photoperiodic signal transduction pathway, which regulates avian seasonal reproduction: a long-day stimulus induces secretion of thyrotropin (TSH) from the pars tuberalis (PT) of the anterior pituitary gland. This PT-derived TSH locally activates thyroid hormone within the hypothalamus, which in turn induces gonadotropin-releasing hormone and then gonadotropin secretion, leading to gonadal growth.
Conclusions:
Studies using TSH receptor-null mice confirmed the involvement of PT-derived TSH in mammalian seasonal reproduction. The pars distalis of the anterior pituitary gland is the major source of circulating TSH. Although the pars distalis and PT are in close proximity, tissue-specific glycosylation of circulating TSH alters its function to avoid cross talk.
Introduction
T
Animals Use Changes in Photoperiod as a Calendar
Animals living outside tropical zones are subjected to dynamic changes in their environment. The impact of environmental changes may be greater in wild animals compared to humans because wild animals do not wear clothes or use air conditioners. To cope with seasonal fluctuations, animals adapt their physiology and behavior, leading to changes in reproduction, migration, molting, and hibernation. Although temperature and precipitation change annually, they provide unreliable seasonal information because summers can often be cool and/or winters warm. In contrast, changes in photoperiod serve as reliable indicators because solstices and equinoxes always occur at exactly the same time of the year. Therefore, it is reasonable that most organisms use changes in photoperiod as a primary cue to sense the seasons. Accordingly, this phenomenon is called photoperiodism (11).
Seasonal Reproduction in Animals
Seasonal reproduction is a clever strategy adopted by animals to maximize survival of their offspring. Birds and small mammals that have short incubation and gestation periods (e.g., a few weeks) mate and give birth to offspring during spring and summer (for review, see Yoshimura (12) and Shinomiya et al. (13)). Hence, they are so-called long-day (LD) breeders. In contrast, relatively large animals, such as goats and sheep, have long gestation periods of around six months. They mate during fall and give birth to their young in spring. Therefore, they are so-called short-day (SD) breeders. In both LD and SD breeders, offspring are born and grow up during spring and summer, when food is abundant and the climate is moderate. Vertebrate reproductive activity is regulated by the hypothalamus–pituitary–gonadal (HPG) axis. Gonadotropin-releasing hormone (GnRH) secreted from the hypothalamus regulates the secretion of gonadotropins (luteinizing hormone [LH] and follicle-stimulating hormone [FSH]) from the pars distalis (PD) of the anterior pituitary gland. Therefore, the HPG axis is activated only at a specific time of the year in seasonally breeding animals.
Seasonal Reproduction Is Highly Sophisticated in Birds
In contrast to mammals, birds have highly sophisticated photoperiodic mechanisms due to flight adaption; birds have a low body weight and develop gonads only during the breeding season. Interestingly, their gonadal size changes more than a hundred-fold within a few weeks (14). Among various avian species, the Japanese quail (Coturnix japonica) is an ideal model for understanding the mechanisms of seasonal reproduction. Japanese quail migrate between Japan and China and show huge differences in gonadal size. In addition, when quail are transferred from SD to LD conditions, an increase in plasma LH concentrations can be observed toward the end of the first LD (15). A classic lesion study using quail suggested that the mediobasal hypothalamus (MBH) is the center that regulates seasonal reproduction in birds (16).
Local Thyroid Hormone Activation Within the MBH Regulates Seasonal Reproduction in Birds
It has been demonstrated that light pulses given at a specific time of the day, called the photoinducible phase, is sufficient to induce testicular development in quail kept under SD conditions (16). Since the photoinducible phase occurs every 24 hours, the circadian clock is involved in the measurement of photoperiodic time. Based on this observation, differential subtractive hybridization analysis was performed using MBH samples from quail, either exposed to light pulse or kept in the dark during the photoinducible phase (17). This analysis identified the LD-induced and -suppressed genes encoding type 2 and type 3 iodothyronine deiodinase (DIO2 and DIO3), respectively, within the ependymal cells (also known as tanycytes) lining the ventrolateral walls of the third ventricle (17,18). DIO2 encodes a thyroid hormone-activating enzyme, which catalyzes the conversion of the precursor, thyroxine (T4), to its bioactive form, triiodothyronine (T3), whereas DIO3 encodes a thyroid hormone-inactivating enzyme that metabolizes both T4 and T3 to inactive reverse T3 and 3,5-diiodo-L-thyronine, respectively. Accordingly, this LD-induced DIO2/DIO3 switching appears to regulate the bioactive thyroid hormone concentration locally within the MBH. Indeed, the T3 concentration within the MBH was about 10-fold higher under LD than SD conditions. However, such T3 concentration changes were not observed in other parts of the brain or peripheral circulation (17). Thyroid hormones were believed to traverse the plasma membrane by passive diffusion due to their lipophilic nature. However, it has been demonstrated that T4 is transported by the organic anion transporting polypeptide 1c1 (Oatp1c1) for conversion to bioactive T3 by DIO2 in tanycytes (19). Thyroid hormones play critical roles in the development and plasticity of the central nervous system (20). Since expression of thyroid hormone receptors was observed in the median eminence where GnRH is secreted to the anterior pituitary gland (17), its ultrastructure was examined by electron microscopy. As a result, it was demonstrated that locally activated thyroid hormone within the MBH induces dynamic morphological changes between the GnRH nerve terminals and glial endfeet in the median eminence (21,22). Many GnRH nerve terminals were encased by the glial endfeet under SD conditions. In marked contrast, many GnRH nerve terminals were in direct contact with the basal lamina located between the median eminence and the hypophyseal portal vessel under LD conditions. Therefore, these morphological changes appear to regulate or modulate seasonal GnRH secretion from the hypothalamus to the portal capillary (17,21,22).
Pars Tuberalis Thyrotropin Is a Springtime Hormone
The availability of the chicken genome sequence opened a new avenue to examine avian function using a genomics approach. Genome-wide transcriptome analysis using microarrays has identified the phototransduction cascade that regulates seasonal reproduction in quail (23). When birds were transferred from nonbreeding SD conditions to breeding LD conditions, the production and secretion of thyrotropin (TSH) was induced in the pars tuberalis (PT) but not in the PD of the pituitary gland (Fig. 1). The PT consists of thin layers of cells that surround the median eminence and pituitary stalk. Unexpectedly, the LD-induced, PT-derived TSH acted on TSH receptors expressed in the adjacent tanycytes within the MBH to induce the expression of DIO2 and reduce the expression of DIO3 through the Gsα-cAMP signaling pathway (23).

Schematic representation of the pituitary gland.
Birds Receive Light Information Directly Within the Brain
In mammals, the eye is believed to be the only photoreceptive organ. In contrast, the pineal organ is also photoreceptive in non-mammalian vertebrates, such as birds. Although the photoreceptive pineal organ resets the circadian clock in birds (24), removal of both eyes and pineal does not affect seasonal reproduction (25). However, blocking light penetration into the brain by injection of India ink under the scalp abolishes LD-induced testicular recrudescence (26). Therefore, these results suggest the existence of extra-retinal and extra-pineal deep brain photoreceptors. Indeed, local illumination of the MBH or the forebrain has been demonstrated to induce gonadal development (27,28). The vertebrate retina contains rod and cone photoreceptors for image-forming vision. These rods and cones contain photopigments called rhodopsin and cone opsins, respectively. In addition to these classic image-forming photopigments, recent studies have identified non-image-forming photopigments, such as pinopsin, VA-opsin, and melanopsin for the entrainment of the circadian clock and pupillary light reflex (24,29 –31). Expression of rhodopsin, VA-opsin, and melanopsin within the avian brain has been reported (32 –35). However, their functional significance has remained unclear. Recently, the expression of a novel opsin, OPN5, was reported in the cerebrospinal fluid (CSF)-contacting neurons of the paraventricular organ in quail (36,37). The CSF-contacting neuron is a bipolar neuron and extends knob-like terminals into the ventricles. Because its morphology resembles the photoreceptor cells in the developing retina, the CSF-contacting neuron was long considered to be a candidate deep brain photoreceptor. The intrinsic photosensitivity of the OPN5-positive CSF-contacting neurons was demonstrated using a slice patch clamp technique (38,39). Importantly, projections of OPN5-positive CSF-contacting neurons into the PT, where the springtime hormone, TSH, is secreted, has been demonstrated (36), and knockdown of OPN5 attenuated LD-induction of TSH in the PT (38). Altogether, OPN5 is considered one of the deep brain photoreceptors that regulates avian seasonal reproduction.
Low Temperature–Induced Peripheral T3 Accelerates Testicular Regression
In contrast to the photoinduction process that regulates testicular growth, the mechanisms involved in seasonal testicular regression have remained largely unexplored. In quail, a low-temperature stimulus accelerates SD-induced testicular regression by shutting down the HPG axis and inducing meiotic arrest and germ-cell apoptosis (40 –42). Low temperature increases the amount of circulating T3, probably mediated by DIO2 in the liver through increased food intake (42). This T3 appears to contribute to adaptive thermogenesis. The timing of T3 induction coincides with the climax of testicular regression, and expression of the genes involved in tadpole metamorphosis were upregulated in the quail testis during this period (42). It has been suggested that thyroid hormones have been involved in seasonal reproduction for several decades. However, some studies reported stimulatory effects, while others reported inhibitory effects. Thus, the effects of thyroid hormones have often been contradictory (14). Consequently, thyroid hormones were considered to play a permissive role in the regulation of seasonal reproduction. Results of recent studies in Japanese quail, however, explain these contradictory effects. Central activation of thyroid hormones by LD-induced DIO2/DIO3 switching results in seasonal testicular growth, whereas peripheral activation by low temperature regulates adaptive thermogenesis and seasonal testicular regression. Therefore, thyroid hormones play dual roles in the regulation of the seasonal reproduction process. Interestingly, thyroid hormones also have opposite effects during metamorphosis in the tadpole; thyroid hormones induce both the outgrowth of the limb and resorption of the tail (43).
Melatonin Decodes Night Length in Mammals
Unlike birds, the eyes are believed to be the only photoreceptive organ in mammals. For example, removal of both eyes completely abolishes entrainment of the circadian clock and the photoperiodic response (44). In addition to the image-forming visual pigments rhodopsin and the cone opsins, the non-image-forming visual pigment melanopsin is expressed in the intrinsically photosensitive retinal ganglion cells and detects light to entrain the circadian clock. Light is detected by visual and non-visual opsins, and this signal is transmitted to the suprachiasmatic nucleus (SCN), which is known as the master circadian pacemaker, via the retinohypothalamic tract (44,45). The SCN controls the synthesis and release of melatonin in the pineal gland. Melatonin is only secreted at night. Therefore, the melatonin secretion profile encodes the length of night. Since pinealectomy abolishes seasonal responses and melatonin replacement therapy restores them in both LD and SD breeders (44 –46), melatonin plays a crucial role in the regulation of seasonal reproduction in mammals.
Most Laboratory Mice Cannot Produce Melatonin
Melatonin is synthesized from serotonin by two enzymes: arylalkylamine N-acetyltransferase (AA-NAT) and hydroxyindole-O-methyltransferase (HIOMT). Most of the commonly used laboratory mice, including C57BL, DBA, BALB, and 129, cannot produce detectable levels of melatonin because they lack the enzymes AA-NAT and/or HIOMT (47,48). Exceptions to this are the mouse strains CBA, C3H, and Mus musculus castaneus. Laboratory mice, even melatonin-proficient strains, are generally considered non-seasonal and inappropriate models to study seasonal reproduction. However, it was noticed that melatonin-proficient mice do not breed well during winter, even though they are maintained under constant temperature and light–dark conditions throughout the year. Therefore, a test was conducted to see whether laboratory mice have the ability to respond to seasonal changes. Although no difference was observed in testicular size, the photoperiodic gene regulation of TSH in the PT and of DIO2/DIO3 in the ECs was observed in the melatonin-proficient CBA strain. In contrast, no response in testicular size or in TSH and DIO2/DIO3 gene expression was observed in the melatonin-deficient C57BL strain (49). These results indicate that mice could be a useful model to study the molecular mechanisms of mammalian seasonal reproduction. Since daily intraperitoneal melatonin injections mimicked the effect of SD conditions on the expression of the aforementioned genes (49), the effect of melatonin was further examined in MT1 and MT2 melatonin receptor knockout mice, as well as in TSH receptor knockout mice. The results obtained demonstrated the role of the MT1 melatonin receptor and TSH receptor in the melatonin-regulated switching of DIO2/DIO3 in mammals (Fig. 2) (49,50).
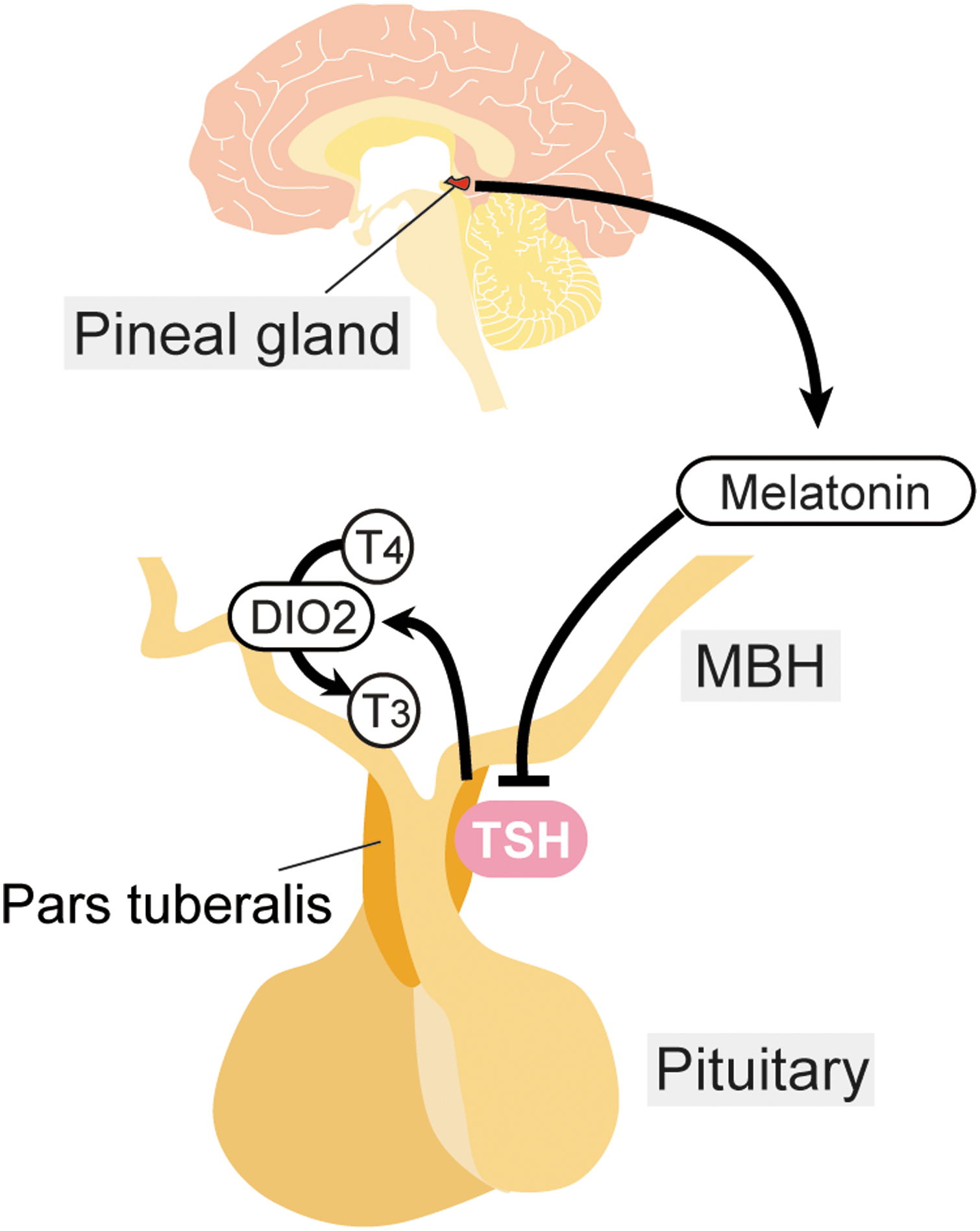
The pars tuberalis (PT) thyrotropin (TSH) is under the control of melatonin in mammals.
Glycosylation Diversifies the Function of TSH
In textbooks, TSH is described as a glycoprotein hormone mainly secreted from the PD of the anterior pituitary gland, and it stimulates the production and secretion of thyroid hormones in the thyroid gland. Thyrotrophs in the PD express receptors for thyrotropin-releasing hormone (TRH) and thyroid hormones, and are under the control of the hypothalamus–pituitary–thyroid (HPT) axis (Fig. 3) (51). PD-derived TSH is positively regulated by TRH and negatively regulated by thyroid hormones by a negative-feedback loop. Unlike PD thyrotrophs, PT thyrotrophs lack receptors for TRH and thyroid hormones, and therefore the regulation of PT-derived TSH is independent of the HPT axis. Instead, PT thyrotrophs have a dense distribution of melatonin receptors, and the synthesis and secretion of PT-derived TSH is under the control of melatonin in mammals (Fig. 2) (52 –54). It is interesting that the regulatory mechanisms and physiological functions of TSH in the PD and PT are completely different. However, the mechanism used by the two TSHs, from the PD and PT, to avoid functional cross talk within the body was not known. Recently, it has been reported that PT-derived TSH secreted into the circulation has minimal bioactivity on the thyroid gland (54). Structural analysis of both TSHs revealed differences in their N-glycosylation pattern. It is well established that glycosylation affects the half-life and bioactivity of glycoprotein hormones (55,56). For example, LH contains N-glycans with sulfate modifications, while FSH has sialylated N-glycans. Although LH and FSH are both regulated by pulsatile GnRH secretion, only LH shows a pulsatile secretion profile. This is because hepatic receptors specifically interact with sulfated N-glycans, and LH, but not FSH, is rapidly metabolized by the liver. PD-derived TSH has sulfated N-glycans with a very short half-life. However, PT-derived TSH has sialylated N-glycans with a longer half-life and greater stability. Although the bioactivities of the PT- and PD-derived TSHs were not different at the cellular level, the PT-derived TSH formed macro-TSH complexes with immunoglobulins and albumin in the circulation to abolish its bioactivity (Fig. 4) (54). Thus, tissue-specific glycosylation appears to differentiate the functions of the TSHs in order to prevent functional cross talk within the body.
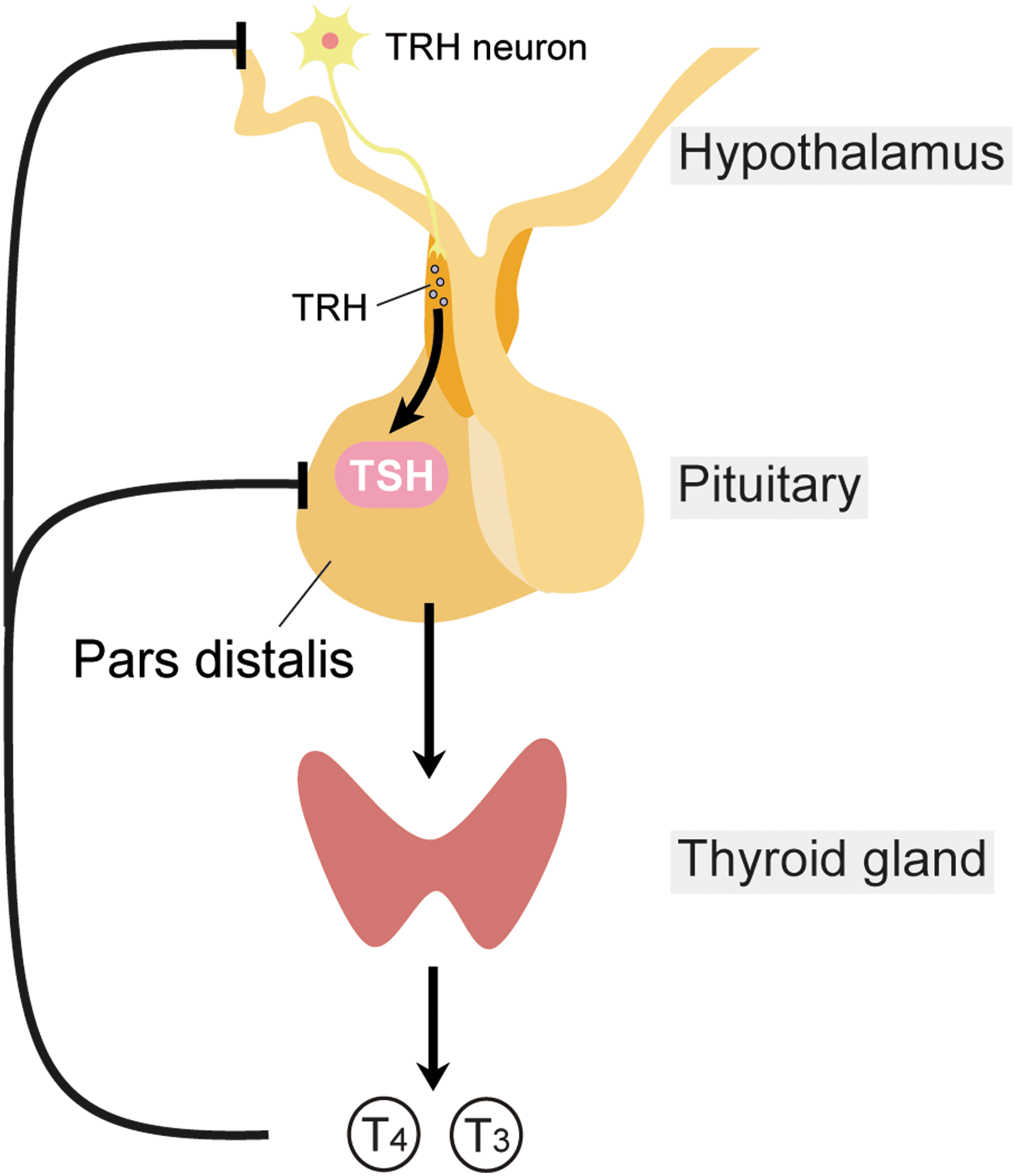
The pars distalis TSH is under the control of the hypothalamus–pituitary–thyroid axis.
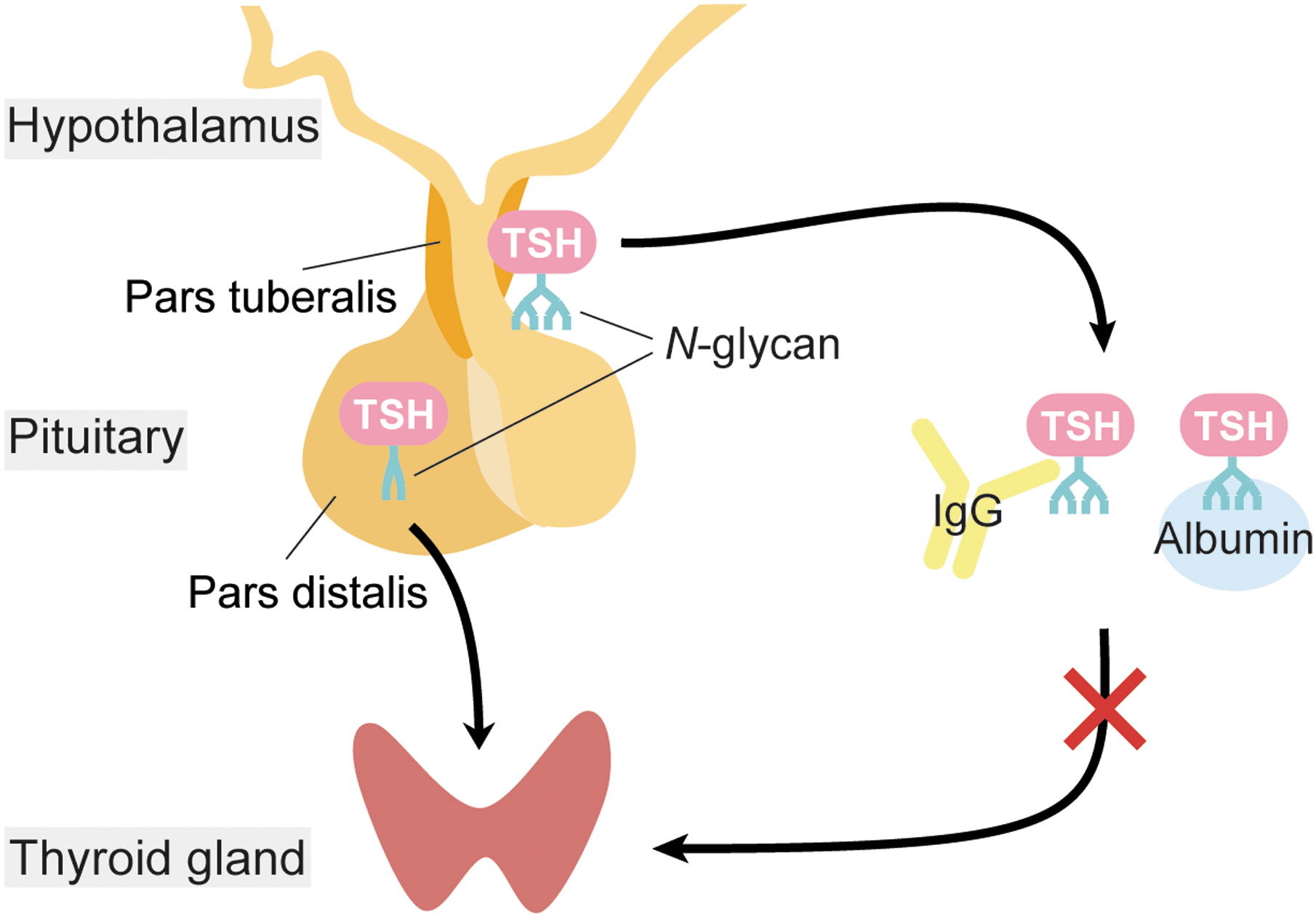
Glycosylation diversifies the function of TSH. PT-TSH has tissue-specific N-glycan modifications and forms macro-TSH complexes with immunoglobulin (IgG) and albumin in the circulation. Macro-TSH cannot stimulate the thyroid gland, which prevents functional cross talk between the two TSHs.
Seasonal Rhythm in Humans
As mentioned earlier, various physiological activities in humans show seasonal fluctuations (1). For example, body weight in humans increases during winter and decreases in summer. SAD, a disease characterized by depression-like symptoms, is prevalent from autumn to winter with remission, or a change to slight hypomania from spring to summer (9). The relationship between thyroid hormones and depression has long been discussed (57). Understanding the mechanisms that animals use to sense seasonal changes may help us to understand human seasonality. Recently, small compounds that lengthen the circadian period by acting directly on the clock protein, CRY, have been discovered (58). This study has been extended by performing structure-activity relationship analyses and novel period-shortening molecules have been developed (59). Since disruption of the circadian clock is associated with SAD (60), development of small molecules that influence clock function might contribute to the treatment of seasonal human disorders such as SAD.
Conclusions
The mechanisms of vertebrate seasonal reproduction had long been a mystery. However, recent comparative studies have uncovered related signal transduction pathways involved in regulating this process in various vertebrates. These studies have revealed the universality and diversity of these photoperiodic mechanisms; the molecules (TSH, DIO2, thyroid hormones) involved are conserved. However, the tissues responsible for these seasonal effects are different (39). Comparative studies can be a powerful approach to uncover the underlying mechanisms of human seasonality.
Footnotes
Acknowledgments
We thank Drs. Peter A. Kopp and Yoshiharu Murata for inviting us to write a review. We also thank Dr. Kathy Tamai for comments on the manuscript. This work was supported in part by JSPS KAKENHI “Grant-in-Aid for Specially Promoted Research” (26000013) and by the Human Frontier Science Program (RGP0030/2015). WPI-ITbM is supported by the World Premier International Research Center Initiative (WPI), MEXT, Japan.
Author Disclosure Statement
The authors have nothing to disclose.