
Abstract
Background:
Ionizing radiation (IR) is a well-known risk factor for papillary thyroid cancer, and it has been reported to deregulate microRNA expression, which is important to thyroid carcinogenesis. Therefore, this study investigated the impact of IR on microRNA expression profile of the normal thyroid cell line (FRTL-5 CL2), as well as its effect on radiosensitivity of thyroid cancer cell lines, especially the human anaplastic thyroid carcinoma cell line (8505c).
Methods:
The global microRNA expression profile of irradiated FRTL-5 CL2 cells (5 Gy X-ray) was characterized, and data were confirmed by quantitative real-time polymerase chain reaction evaluating the expression of rno-miR-10b-5p, rno-miR-33-5p, rno-miR-128-1-5p, rno-miR-199a-3p, rno-miR-296-5p, rno-miR-328a-3p, and rno-miR-541-5p in irradiated cells. The miR-199a-3p and miR-10b-5p targets were validated by quantitative real-time polymerase chain reaction, Western blot, and luciferase target assays. The effects of miR-199a-3p and miR-10b-5p on DNA repair were determined by evaluating the activation of the protein kinases ataxia-telangiectasia mutated, ataxia telangiectasia, and Rad3-related and the serine 39 phosphorylation of variant histone H2AX as an indirect measure of double-strand DNA breaks in irradiated FRTL-5 CL2 cells. The impact of miR-10b-5p on radiosensitivity was analyzed by cell counting and MTT assays in FRTL-5 CL2, Kras-transformed FRTL-5 CL2 (FRTL KiKi), and 8505c cell lines.
Results:
The results reveal that miR-10b-5p and miR-199a-3p display the most pronounced alterations in expression in irradiated FRTL-5 CL2 cells. Dicer1 and Lin28b were validated as targets of miR-10b-5p and miR-199a-3p, respectively. Functional studies demonstrate that miR-10b-5p increases the growth rate of FRTL-5 CL2 cells, while miR-199a-3p inhibits their proliferation. Moreover, both of these microRNAs negatively affect homologous recombination repair, reducing activated ataxia-telangiectasia mutated and Rad3-related protein levels, consequently leading to an accumulation of the serine 39 phosphorylation of variant histone H2AX. Interestingly, the overexpression of miR-10b-5p decreases the viability of the irradiated FRTL5-CL2 and 8505c cell lines. Consistent with this observation, its inhibition in FRTL KiKi cells, which display high basal expression levels of miR-10b-5p, leads to the opposite effect.
Conclusions:
These results demonstrate that IR deregulates microRNA expression, affecting the double-strand DNA breaks repair efficiency of irradiated thyroid cells, and suggest that miR-10b-5p overexpression may be an innovative approach for anaplastic thyroid cancer therapy by increasing cancer cell radiosensitivity.
Introduction
M
Several studies have shown that deregulated miRNA expression contributes to thyroid carcinogenesis, contributing to the development and the progression of the malignant phenotype (2 –4). For example, overexpression of the miR-221/222 cluster in papillary thyroid carcinomas (PTC) increases thyroid cell proliferation by downregulating p27 kip1 protein levels and subsequent enhancement of the G1/S transition (3,4), whereas downregulation of let-7 family members in differentiated and undifferentiated thyroid carcinomas favors cancer progression due to their ability to modulate cell cycle regulators and oncogenes, including RAS, HMGA1 and HMGA2, MYC, CCNB1, and CCNE2 (4).
Moreover, miRNA expression also has a critical role in the differentiation and proliferation of normal thyroid cells. For example, it has previously been reported that thyrotropin (TSH) stimulation of rat thyroid cells leads to the downregulation of miR-1, miR-28a, miR-290-5p, miR-296-3p, and miR-297a that target CREB1, a transcription factor activated by the cAMP pathway and required for thyroid cell proliferation and differentiation (5).
Ionizing radiation (IR) is one of the critical risk factors for PTC (6 –8). This is illustrated by several studies analyzing the effect of the Chernobyl accident, which have documented a significant increase of the PTC incidence (7.7 × ) in the children living in Ukraine and Belarus due to the carcinogenic effects of beta and gamma radiation after exposure to the radioactive 131I isotope (7,8). Data on the nuclear explosions in Japan, exposure to X-ray therapy for conditions such as tinea capitis and cervical and childhood tumors also illustrate that radiation is a risk factor for thyroid carcinogenesis (6). It has also been shown that radiation can induce RET/PTC1 rearrangements in human normal thyroid tissues transplanted into SCID mice (9), a gene fusion that represents the driving genetic alteration in 13–43% of PTCs (10).
The carcinogenic effects of IR can be attributed to DNA damage, especially, double-strand breaks (DSB), which can for example generate RET/PTC rearrangements (11). DSB repair signaling, in turn, mostly homologous recombination (HR) and non-homologous end joining (NHEJ), are triggered to maintain genomic integrity (12). In this context, alterations in the miRNA expression profile have been reported as fundamental in radiation-induced response (13). Several miRNAs, such as miR-421 and miR-182, which are regulated by IR, also target HR genes such as ATM and BRCA1, thereby sensitizing cells to IR (13).
Anaplastic thyroid carcinoma (ATC) is a rare (1–2%) and highly aggressive and lethal type of cancer (median survival time of five to six months) that is largely refractory to all conventional therapies, including radiotherapy (14). Global miRNA analysis revealed several deregulated miRNAs in ATC samples (15), which have been previously reported to be important for radioresistance, including miR-146 that targets BRCA1 (16), miR-30 that regulates the PI3K pathway (17), and let-7, which is important for thyroid differentiation and radiosensitivity (18).
The identification of IR-induced changes in miRNA expression in thyroid cells and their impact on DNA repair efficiency, as well as the characterization of potential targets that sensitize tumor cells to IR, represent an important issue, and was therefore the aim of this study. Consequently, the global miRNA profile in irradiated differentiated rat thyroid cells was analyzed. Among the IR-deregulated miRNAs, attention was focused on miR-10b-5p and miR-199a-3p, which are upregulated and downregulated, respectively, by IR. The study demonstrates that miR-10b-5p and miR-199a-3p target Dicer1 and Lin28b, respectively. Moreover, miR-10b-5p overexpression stimulates thyroid cell proliferation and favors thyroid cell radiosensitivity, whereas miR-199a-3p shows the opposite effect.
Material and Methods
Small RNA sequencing
Small RNA sequencing experiments were performed on four samples: FRTL-5 CL2 non-exposed (control) and irradiated samples at 1 and 6 hours post 5 Gy X-ray exposure by Genomix4Life (Salerno, Italy). Prior to further analysis, a RNA quality check was performed on sequencing data using Agilent 4200 TapeStation System (Agilent Technologies). Then, indexed libraries of the samples were sequenced (1 × 50, ∼30,000,000 total reads/sample) on a Hiseq2500 Illumina platform. Mature miRNA identification was performed on all four samples using the online tools sRNAtoolbox (19). The analysis was performed by aligning the reads on the Rattus norvegicus genome using the MiRBase v21 database. Differential expression probability for each one of the miRNA (p ≥ 0.7) and fold change (>2) were calculated using NOISeq (20), comparing irradiated with paired control samples (Table 1).
The values indicate the fold change comparing irradiated with paired control samples. Probability means the differential expression probability for each miRNA. The microRNAs highlighted in bold are those validated by quantitative real-time polymerase chain reaction.
miR or miRNA, microRNA.
Cell culture, irradiation, and transfection
FRTL-5 CL2 thyroid cells, derived from three-week-old Fischer rats, were cultured in Coon's modified Ham's F-12 medium (Euroclone), supplemented with 5% calf serum, 1% L-glutamine 10 mM, 1% penicillin/streptomycin (Life Technologies), and a six-hormone mixture (1 mIU/mL of TSH, 10 μg/mL of insulin, 5 μg/mL of transferrin, 10 nM of hydrocortisone, 10 ng/mL somatostatin, and 10 ng/ml glycyl-L-histidyl-L-lysine acetate- 6H; Sigma–Aldrich) (21 –23). Kras-transformed FRTL-5 CL2 cells (FRTL KiKi) were grown in Coon's modified Ham's F-12 medium (Euroclone), supplemented with 10% calf serum, 1% L-glutamine 10 mM, and 1% penicillin/streptomycin (Life Technologies) and without 6H (22,23). Both FRTL-5 CL2 and FRTL KiKi cell lines were generated at the Dipartimento di Medicina Molecolare e Biotecnologie Mediche, Università degli Studi di Napoli Federico II, Naples, Italy. The human papillary thyroid carcinoma cells lines, TPC-1 (RET/PTC1 rearrangement) and BCPAP (BRAFV600E mutation), the ATC cell lines, FRO (p53-null) and 8505c (BRAFV600E mutation), and the human embryonic kidney cell line (HEK293) were grown in Dulbecco's modified Eagle's medium (Life Technologies) supplemented with 10% fetal bovine serum, 1% L-glutamine 10 mM, and 1% penicillin/streptomycin (Life Technologies). The Short Tandem Repeat (STR) profile of the human thyroid carcinoma cell lines used in this study was carried out on the samples of each provided cell line using nine highly polymorphic STR loci plus amelogenin (Cell IDTM System; Promega) by IRCCS Azienda Ospedaliera Universitaria San Martino—IST (Genoa, Italy). Detection of amplified fragments was obtained using a ABI PRISM 3100 Genetic Analyzer (Thermo Fisher Scientific). Data analysis was performed with GeneMapper software v4.0 (Thermo Fisher Scientific). The STR profile of all the tested human thyroid carcinoma cell lines did not show any difference when compared to the profiles published by one or more of the international databases.
Cells were exposed to a single X-ray dose of 5 Gy (1 Gy/min at 320 KV, 12.5 mA, 50 cm source to surface distance; X-RAD 320; Precision X-Ray) at about 70% confluence and collected after 1–48 hours. As an index of cell viability, the commercially available MTT assay was used (Sigma–Aldrich). Briefly, cells (2 × 103 cells/well) were seeded in 96-well plates and incubated for one hour with MTT reagent, diluted at final concentration of 0.5 mg/mL in cell medium, and then solubilized in 100 μL of dimethyl sulfoxide. Measures were performed at 570 nm using an ELx800 microplate reader (BIO-TEK).
For transient overexpression of miR-199a-3p and miR-10b-5p and miR-10b-5p inhibition, cells were transfected with 50 nM pre-miRNA precursors (pre-miR-10b-5p, PM10133; pre-miR-199a-3p, PM11779; Anti-miR-10b-5p, AM17000; Ambion) or control no-targeting scramble (#1, AM17110; Ambion) using Lipofectamine 2000 (Life Technologies) following the manufacturer's instructions.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from cells (2 × 105 cells/60 cm2 culture dish) using the Trizol reagent (Life Technologies). Total RNA (1 μg) of each sample was used to obtain single-strand cDNA with the QuantiTect Reverse Transcription Kit (Qiagen). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed with the CFX96 thermocycler (Bio-Rad) in 96-well plates. Each PCR reaction included 10 μL of 2 × Sybr Green (Bio-Rad), 200 nM of each primer, and 20 ng of the previously generated cDNA. The oligonucleotides for qRT-PCR, comprising exon–exon junctions, were purchased from Integrated DNA Technologies and designed with Primer-BLAST software (Dicer1: Fw CAC ATG CCT CCT ACC ACT ACA AT, Rv TGC TTG GTT ATG AGG TAG TCC A; Lin28b: Fw AGC CCC TTGG ATA TTC CAG TC, Rv AAT GTG AAT TCC ACT GGT TCT CCT; Rpl4: Fw GAT GAA TTG TAC GGC ACT TGG, Rv TCT TTG GAT CTC TGG GCT TTT TC). Relative gene expression was determined using the comparative C(T) method (24). Rpl4 was used as the housekeeping gene.
To assess miRNA expression, 1 μg of total RNA of each sample was reverse transcribed with the miScript reverse transcription Kit (Qiagen) according to the manufacturer's instruction. The cDNA served as the template for qRT-PCR. qRT-PCR analysis was performed using a miRNA-specific miScript Primer Assay (forward primer) (Qiagen), as described above, and the miScript SYBR Green PCR Kit (Qiagen), which contains the miScript Universal Primer (reverse primer) and QuantiTect SYBR Green PCR Master. The following miScript Primers used for this work were: miR-10b-5p (MS00033194), miR-33-5p (MS00033439), miR-128-1-5p (MS00026656), miR-199a-3p (MS00013195), miR-296-5p (MS00016457), miR-328a-3p (MS00027349), miR-541-5p (MS00013720), and Let-7b (MS00000007). RNU6 (MS00033740) was used for normalization.
Western blot
Cells (1 × 106 cells) were homogenized in RIPA lysis buffer (20 mM of Tris-HCl, pH 7.5, 5 mM of EDTA, 150 mM of NaCl, 1% Nonidet P40, and a mix of protease inhibitors). Cell lysate proteins (50 μg) were then subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto Immobilon-P Transfer membranes (Millipore). The membranes were blocked with 5% non-fat milk proteins and probed with the indicated antibodies at the appropriate dilutions: Dicer1 (1:1000; sc-136981; Santa Cruz Biotechnology), Lin28b (1:1000; #5422; Cell Signaling Technology), γ-tubulin (1:1000; sc-8035; Santa Cruz Biotechnology), HMGA2 (1:500; α939, polyclonal antibody raised against a synthetic peptide located in the NH2-terminal region), Vinculin (1:1000; sc-7649; Santa Cruz Biotechnology), p-ATM (1:1000; Rockland), ATM (1:1000; ab91; Abcam), ATR (1:2000; Novus Biological), γH2AX (1:1000; #05-636; Upstate), and Gapdh (1:3000; sc-32233; Santa Cruz Biotechnology). Membranes were then incubated with a horseradish peroxidase-conjugated secondary antibody (1:3000) for 60 minutes at room temperature, and the signals were detected by a Western blot detection system (Thermo Fisher Scientific).
Flow cytometry
FRTL-5 CL2 cells (2 × 105 cells/well) were seeded in six-well culture plates, and 24 hours later, the cells were exposed to 5 Gy X-ray, as mentioned above. After 24 hours, cell pellets were re-suspended in 500 μL of propidium iodide solution (0.1% Triton X-100, 0.1% RNAse, and 2 μg/mL propidium iodide) and incubated on ice for five minutes. Cell cycle analysis was conducted on a FACScan flow cytometer (Becton Dickinson) and analyzed with CELL-FIT software (Becton Dickinson).
Plasmids and luciferase target assay
Predicted targets of miR-10b-5p and miR-199a-3p were searched for using the online software packages TargetScan (
Growth curve assay
Cells (2 × 105 cells) were plated in 60 cm2 dishes, and 24 hours later, cells were transfected or irradiated as described above. A time course at 0, 1, 6, 12, 24, and 48 hours was performed using trypan blue to evaluate cell growth rate.
Statistical analysis
Results are expressed as mean ± standard deviation and were analyzed by the nonparametric Mann–Whitney test (when comparing two groups) or by the nonparametric Kruskal–Wallis test followed by Dunn's multiple comparison tests (when comparing three or more groups). Statistical analyses were performed using the software Graphpad Prism v6.0 (GraphPad Software, Inc.), and the difference was considered significant when p < 0.05.
Results
Effects of IR on FRTL-5 CL2 cells
First, rat thyroid FRTL-5 CL2 cells, which maintain markers of thyroid differentiation in vitro (21), were exposed to 5 Gy X-ray. Then, cell proliferation, viability, and cycle were assessed (Fig. 1). Regarding the dose of exposure, a dose curve (1–10 Gy X-ray) was previously performed, and no cytotoxicity was observed in this cell line (25). The decision to proceed with the 5 Gy dose is based on the linear-exponential model (cancer relative risk and radiation dose to thyroid curve) based on studies analyzing the thyroid cancer risk after exposure to external radiation in several cohorts in children (6). IR-exposed FRTL-5 CL2 cells exhibited a reduced growth rate compared to non-exposed paired control cells (Fig. 1A), whereas no changes were observed in cell viability up to 48 hours after irradiation (Fig. 1B). Moreover, irradiated cells accumulated in the G2/M phase of the cell cycle, 24 hours after the exposure, in comparison with controls (34.64% vs. 23.84%; p < 0.05; Fig. 1C). In parallel, IR led to a decrease of the percentage of cells in the S phase (10.48% vs. 20.93%; Fig. 1C).
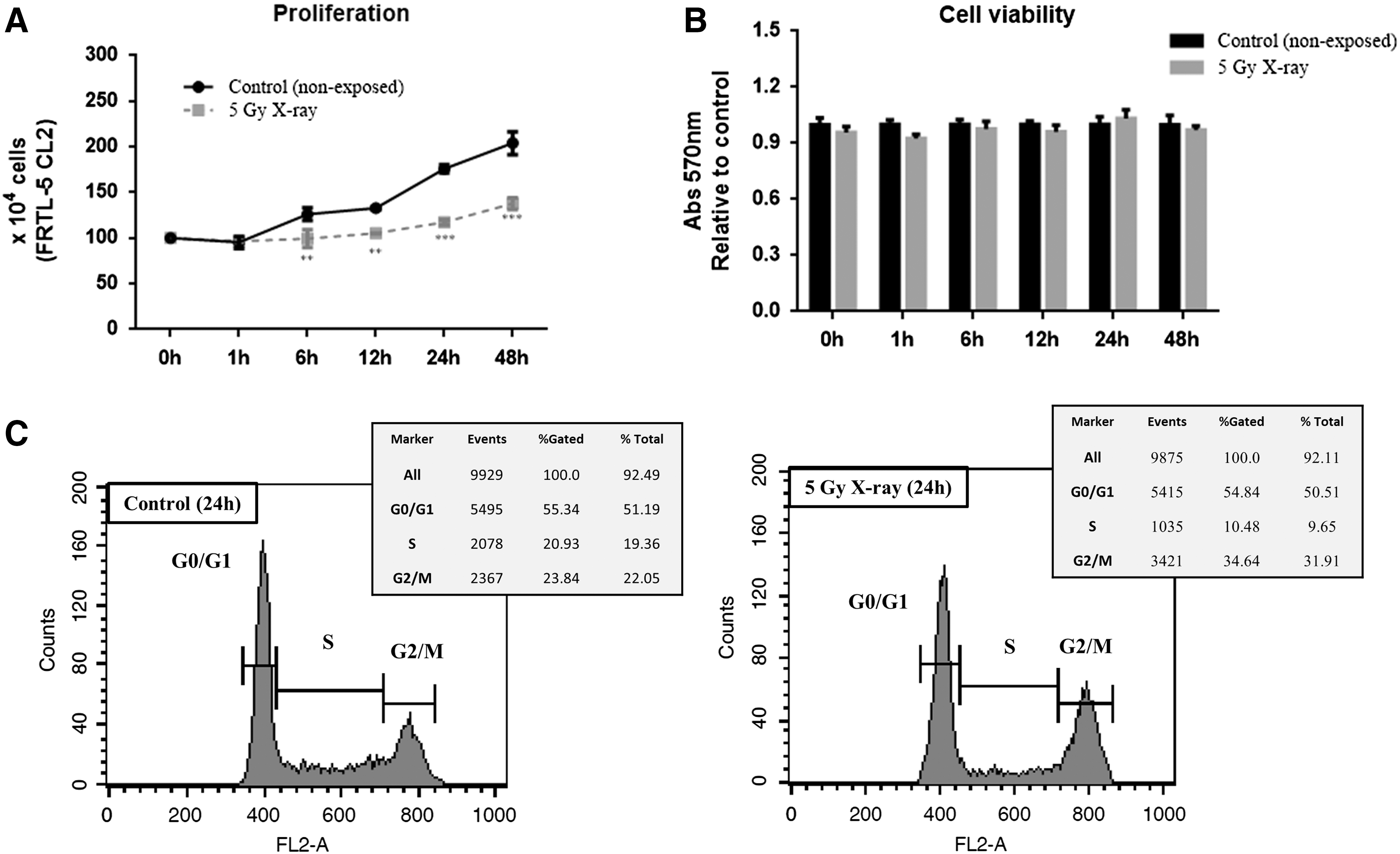
Effects of ionizing radiation (IR) on FRTL5 cell line. FRTL-5 CL2 cells were exposed to 5 Gy X-ray, and the following parameters were evaluated. (
Differential miRNA expression profile of irradiated FRTL-5 CL2 cells
Next, the miRNA expression profile was analyzed by small RNA sequencing in the irradiated FRTL-5 CL2 cell line. A list of differentially expressed miRNAs in irradiated FRTL-5 CL2 cells and paired non-exposed control cells was obtained, one and six hours post irradiation (fold change ≥2.0; differential expression probability ≥0.70; reads >20; Table 1). Seven miRNAs that were significantly downregulated one hour post irradiation and 10 miRNAs that were upregulated six hours after exposure were found (Table 1).
Then, the sequencing results were validated by assessing the expression of miR-10b-5p, miR-33-5p, miR-128-1-5p, miR-199a-3p, miR-296-5p, miR-328a-3p, and miR-541-5p in irradiated cells in comparison with paired non-exposed control cells at 1, 6, 12, and 24 hours post irradiation by qRT-PCR (Fig. 2). As shown in Figure 2, the expression of these miRNAs is downregulated one hour post irradiation and then upregulated after six hours, returning to non-exposed control expression levels 24 hours after irradiation. Subsequently, the study focused on miR-10b-5p and miR-199a-3p because of their previously shown role in thyroid carcinogenesis (23,26) and the biological relevance of their predicted targets for thyroid cancer.
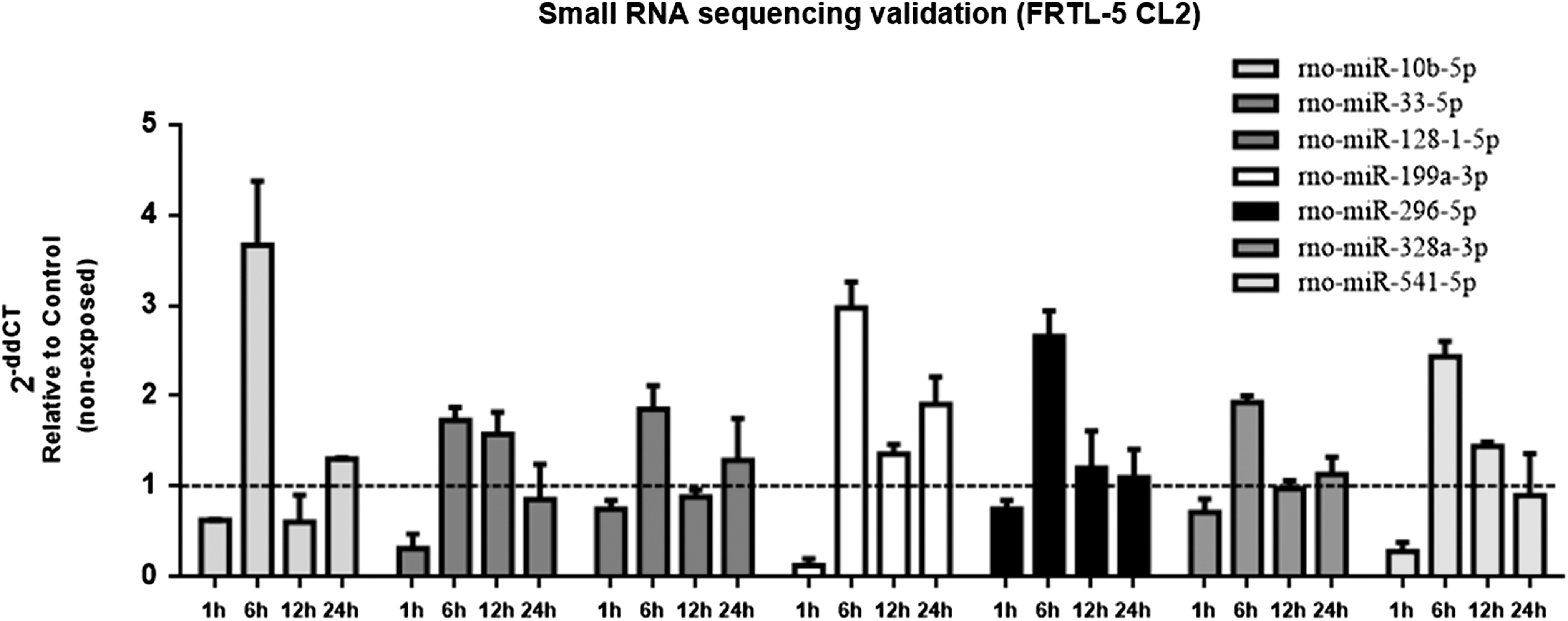
Validation of small RNA sequencing. Samples of control (non-exposed) and irradiated (5 Gy X-ray) FRTL-5 CL2 cells used for array sequencing (1 and 6 hours), as well as additional samples at 12 and 24 hours after IR exposure were selected to validate the small RNA sequencing, evaluating the expression of the following mature miRNAs by quantitative real-time polymerase chain reaction (qRT-PCR): rno-miR-10b-5p, rno-miR-33-5p, rno-miR-128-1-5p, rno-miR-199a-3p, rno-miR-296-5p, rno-miR-328a-3p, and rno-miR-541-5p. Results are expressed as relative to paired non-exposed control. U6 was used to normalize miRNA levels. Data are represented as the mean value ± SD from two independent experiments, including the samples used for the small RNA sequencing. miR or miRNA, microRNA.
miR-199a-3p targets Lin28b
Bioinformatic algorithms predicted several targets for miR-199a-3p. Among them, the study focused on Lin28b because it has the highest target score in the miRDB and TargetScan software analyses and also contains two conserved binding sites for miR-199a-3p. To validate Lin28b as target of miR-199a-3p, miR-199a-3p in FRTL-5 CL2 cells were overexpressed (Fig. 3A). A reduction of 48% and 63% of Lin28b protein levels was observed at 48 and 72 hours, respectively, in the miR-199a-3p-transfected cells in comparison with the scrambled oligonucleotide (Fig. 3B). Lin28b mRNA levels were also downregulated in the cells expressing miR-199a-3p (Fig. 3C), indicating that miR-199a-3p decreases Lin28b protein levels by affecting, at least partially, mRNA stability.

Lin28b is a target of miR-199a-3p. FRTL-5 CL2 cells were transfected with miR-199a-3p or no-targeting scramble oligonucleotide precursors and 48 and 72 hours after transfection the following parameters were evaluated. (
Since it is known that Lin28a and Lin28b selectively block the expression of let-7 miRNAs (27) that in turn target HMGA2, an important protein involved in thyroid cancer progression (28,29), the HMGA2 protein levels in the miR-199a-3p-transfected cells were also evaluated. In agreement with the ability of miR-199a-3p to target Lin28b, a reduction of HMGA2 protein levels by 53% and 33% at 48 and 72 hours after transfection, respectively, was observed compared to the control (Fig. 3B) and was associated with an increase in let-7b expression (Fig. 3D). It is noteworthy that the overexpression of miR-199a-3p caused no changes in let-7a/e expression. These results indicate that miR-199a-3p also regulates HMGA2 expression and suggest that the Lin28b-let-7b pathway could be involved in this phenomenon.
To determine whether the direct interaction between miR-199a-3p and Lin28b mRNA is responsible for the attenuation of Lin28b protein levels, 3′-UTR of Lin28b, comprising the two conserved binding sites for miR-199a-3p in sense and antisense orientation, was inserted downstream of the luciferase ORF. The luciferase activity of Lin28b 3′-UTR in sense orientation, evaluated in HEK293 cells, was significantly diminished (59%) compared to the scrambled oligonucleotide-transfected cells, whereas only a slight decrease in the luciferase activity was observed when cells were transfected with the antisense construct (Fig. 3E). Therefore, these results indicate that miR-199a-3p directly regulates Lin28b expression by binding to the 3′-UTR of Lin28b, leading to a decrease of its expression.
miR-10b-5p targets Dicer1
Potential targets of miR-10b-5p were also searched for using the online software TargetScan, which predicts miRNA binding sites within 3′-UTRs of mRNAs (30). Among the predicted targets, our attention was concentrated on Dicer1 because it has two conserved binding sites for miR-10b-5p, plays a central role in miRNA processing (31), and has been reported to be critical for thyroid cell proliferation and differentiation (32,33). To validate the influence of miR-10b-5p on Dicer1 expression, the Dicer1 protein levels in FRTL-5 CL2 cells transfected with miR-10b-5p oligonucleotide precursor or control-no-targeting oligonucleotide were evaluated by Western blot analysis. A significant decrease by 59% and 36% levels at 48 and 72 hours, respectively, of the Dicer1 protein was found in the miR-10b-5p precursor-transfected cells in comparison with the control cells (Fig. 4A and B). Conversely, no changes were detected in Dicer1 mRNA levels after transfection with miR-10b-5p (Fig. 4C), indicating that post-transcriptional mechanisms might be responsible for the downregulation of Dicer1 mediated by miR-10b-5p.
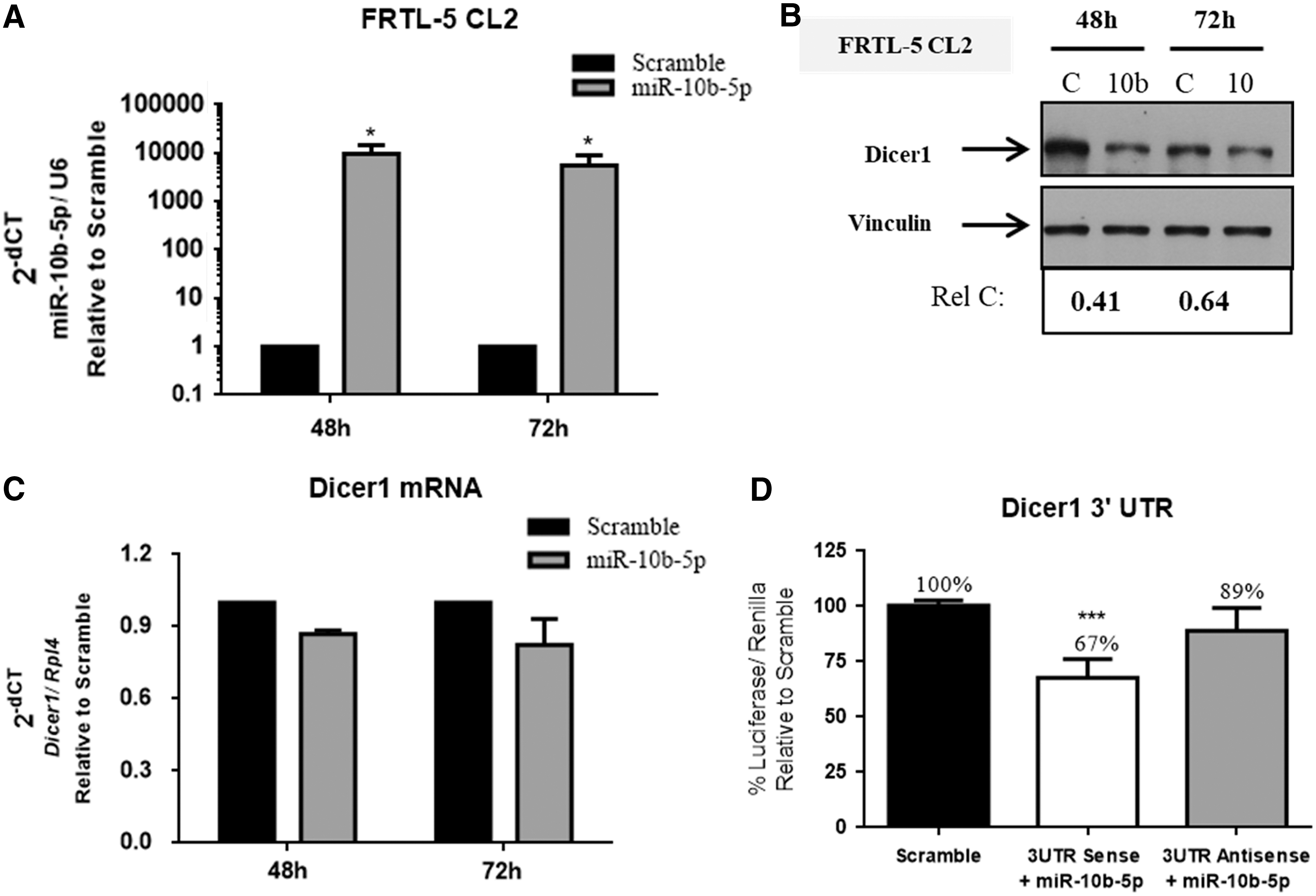
Dicer1 is a target of miR-10b-5p. FRTL-5 CL2 cells were transfected with miR-10b-5p or no-targeting scramble oligonucleotide precursors, and 48 and 72 hours after transfection the following parameters were evaluated. (
To investigate whether miR-10b-5p could interact with Dicer1 mRNA, the 3′-UTR sequence of Dicer1, containing the predicted miR-10b-5p binding site, was inserted downstream of the luciferase ORF, in sense or antisense orientation. The luciferase activity of Dicer1 3′-UTR in sense orientation was reduced after transfection of miR-10b-5p when compared to scramble (33%; p < 0.001), while a weak change was observed with the Dicer1 3′-UTR antisense vector (11%; Fig. 4D). The validation of the miR-10b-5p binding sites within the 3′-UTR region of Dicer1 mRNA and the decreased protein levels of Dicer1 by miR-10b-5p without affecting its mRNA levels strongly suggest that repression of its translation is the responsible post-transcriptional mechanism.
Lin28b and Dicer1 protein levels correlate with miR-199a-3p and miR-10b-5p expression in the IR-exposed FRTL-5 CL2 cells
Having demonstrated that Lin28b and Dicer1 are the targets of miR-199a-3p and miR-10b-5p, respectively, the next step was to verify the presence of a correlation between the expression of these miRNAs and their respective targets in FRTL-5 CL2 irradiated cells by performing qRT-PCR and Western blot analyses. The data unravel that IR induces a biphasic increment in Lin28b mRNA levels (around 2 × ), 1 and 12 hours post irradiation (Fig. 5A). Consistently, IR increased Lin28b protein levels one hour after exposure (2 × ) when compared to non-exposed control cells, returning to control levels after six hours (1.0 × ; Fig. 5B). It is noteworthy that IR also induced HMGA2 protein levels one hour after exposure (1.6 × ), which correlates with IR-mediated Lin28b expression at this point (Fig. 5B). No correlation was observed between HMGA2 and Lin28b protein levels, 6 and 24 hours after IR exposure (Fig. 5B).

IR modulates Lin28b and Dicer1 expression in the FRTL-5 CL2 cell line. Cells were exposed to 5 Gy X-ray, and (
As far as Dicer1 expression is concerned, its protein levels increase one hour post irradiation (1.77 × ), then decrease after six hours (0.50 × ), returning to control levels 24 hours after irradiation (0.90 × ; Fig. 5C). Nevertheless, no alteration in Dicer1 mRNA levels was observed after IR exposure (Fig. 5D).
Role of miR-10b-5p and miR-199a-3p in thyroid cell proliferation and DSB repair machinery
Then, the study considered whether the overexpression of miR-10b-5p and miR-199a-3p could influence cell proliferation and DSB-damage response induced by IR in the FRTL-5 CL2 cell line. First, FRTL-5 CL2 cells were transfected with miR-10b-5p, miR-199a-3p, or no-targeting scramble oligonucleotide precursors to evaluate the impact of these miRs on the proliferation of FRTL-5 CL2 cells. The results demonstrate that miR-10b-5p increases the growth rate of FRTL-5 CL2 cells compared to cells transfected with scrambled oligonucleotides (Fig. 6A). Conversely, the overexpression of miR-199a-3p attenuates cell proliferation (Fig. 6A). To investigate whether these miRNAs could modify the effects of IR on thyroid cell proliferation, FRTL-5 CL2 cells were irradiated 48 hours after the transfection with scramble, miR-10b-5p, or miR-199a-3p oligonucleotide precursors (the start point, P0) because at that point, miR-10b-5p and miR-199a-3p have the highest expression levels compared to scramble and efficiently downregulate their respective targets, Dicer1 and Lin28b. Since miR-10b-5p increases and miR-199a-3p decreases the growth rate of thyroid cells (Fig. 6A), the start point has different cell numbers for each condition. It was observed that the growth rate of miR-10b-5p-overexpressing cells was severely compromised by IR up to 24 hours post irradiation (Fig. 6B). In contrast, IR slightly reduced the number of cells transfected with the scrambled oligonucleotide or miR-199a-3p six hours after exposure, but cells recovered the ability to proliferate after 24 hours (Fig. 6B). Moreover, cell viability was evaluated with an MTT assay that showed a decrease in the relative number of viable miR-10b-5p-overexpressing cells (20%) compared to controls 24 hours post irradiation, while cells transfected with miR-199a-3p displayed increased viability (30%; Fig. 6C). Taken together, these results indicate that miR-10b-5p induces radiosensitivity in FRTL-5 CL2 cells, whereas miR-199a-3p acts in the opposite way.
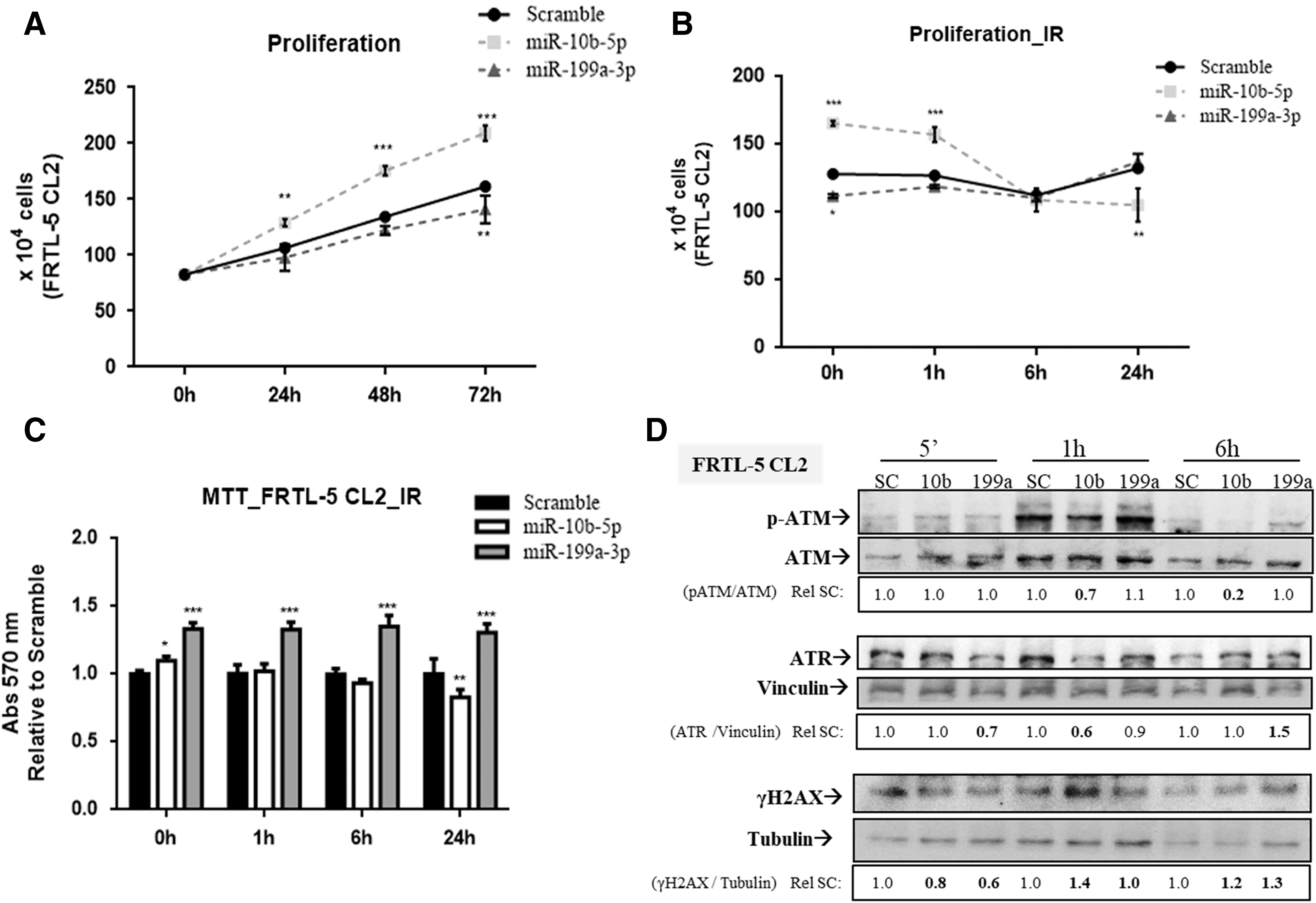
Role of miR-199a-3p and miR-10b-5p in proliferation and double-strand break (DSB) repair of the FRTL-5 CL2 cell line. (
Subsequently, the efficiency of the DSB repair machinery was analyzed by Western blot analysis. FRTL-5 CL2 cells were transfected with miR-10b-5p and miR-199a-3p or scramble, and 48 hours after transfection, cells were exposed to 5 Gy X-ray (Fig. 6D). Cells overexpressing miR-10b-5p have lower levels of activated ATM (p-ATM; 1 h: 0.7 vs. 1.0; 6 h: 0.6 vs. 1.0) and ATR (1 h: 0.7 vs. 1.0; 6 h: 0.6 vs. 1.0), one and six hours post irradiation compared to scramble. As far as the cells transfected with miR-199a-3p are concerned, they have lower basal protein levels of ATR when compared to scramble, and its relative expression was progressively induced by IR, with a peak six hours after irradiation (1.5 vs. 1.0). However, no relative changes of p-ATM levels were observed in miR-199a-3p-transfected cells.
Modification of γH2AX was used as an indirect measure of DSB lesions, since it is an early DSB sensor (34). The results show that γH2AX levels are higher in cells transfected with miR-10b-5p than scramble at one (1.4 vs. 1.0) and six hours (1.2 vs. 1.0) after irradiation (Fig. 6D). Interestingly, even though no relative alteration in γH2AX levels was noticed in miR-199a-3p-overexpressing cells in comparison with scramble in the first hour after irradiation, an accumulation of γH2AX levels was observed six hours later compared to scramble (1.3 vs. 1.0; Fig. 6D). Therefore, these results suggest that miR-10b-5p and miR-199a-3p decrease DSB repair efficiency of irradiated FRTL-5 CL2 cells.
Effects of miR-10b-5p inhibition on irradiated Kras-transformed FRTL-5 CL2 cells
Next, the effects of IR on Kras-transformed FRTL-5 CL2 (FRTL KiKi) cells were analyzed. This choice was motivated by previous studies showing miR-10b-5p overexpression in these cells (23). First, it was confirmed that FRTL KiKi cells have higher basal levels of miR-10b-5p than FRTL-5 CL2 cells (around 25 × ; Fig. 7A). Further, FRTL KiKi cells were exposed to 5 Gy X-ray in order to evaluate the expression of miR-10b-5p. As shown in Figure 7B, IR upregulates miR-10b-5p in a time-dependent manner one hour (2 × ) and six hours (6 × ) post irradiation, returning to control levels after 24 hours. Moreover, the data show that IR strongly reduces the growth rate (Fig. 7C), with a consistent decrease in the number of viable cells (Fig. 7D) of the irradiated cells in comparison with paired non-exposed cells (control). Therefore, these results indicate that irradiated FRTL KiKi cells recapitulate the IR-induced behavior of FRTL-5 CL2 cells transfected with miR-10b-5p precursor oligonucleotide.
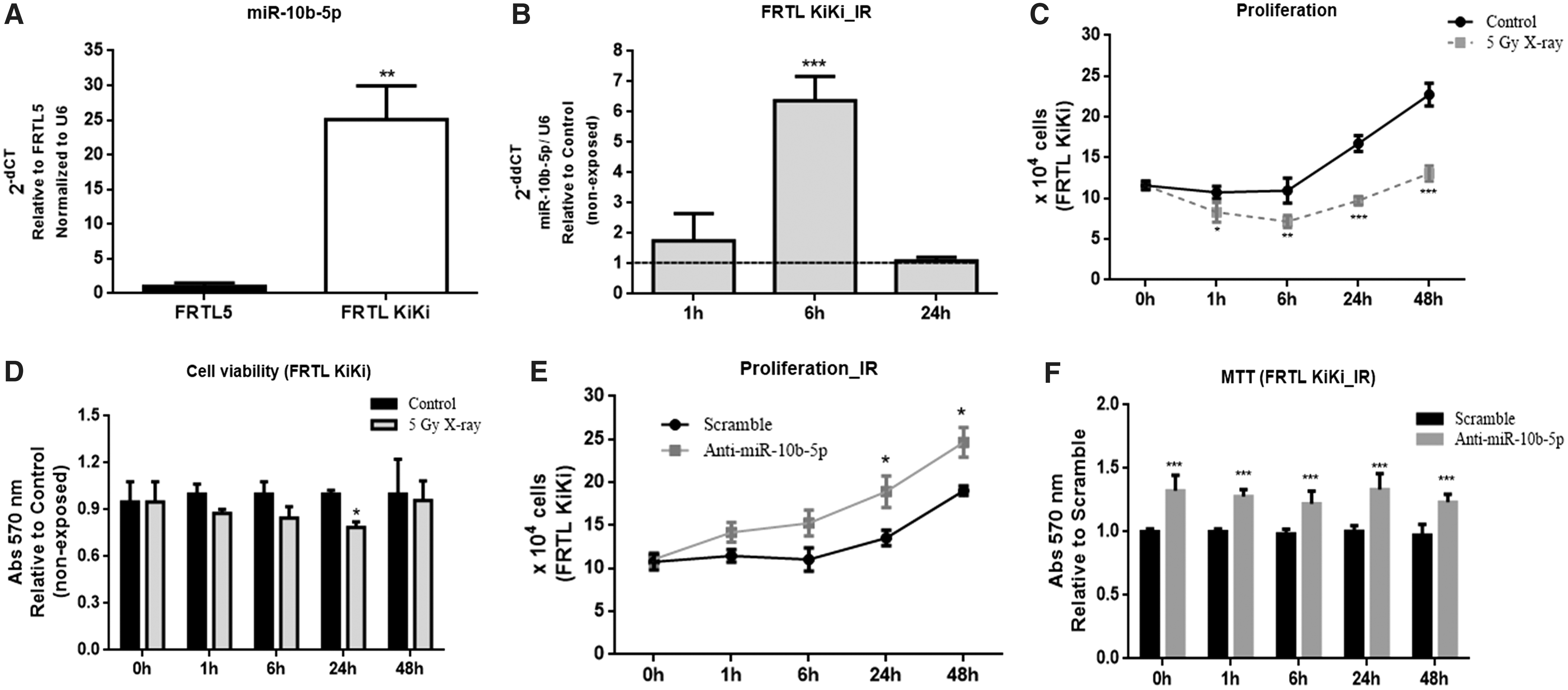
Effects of miR-10b-5p on Kras-transformed FRTL5 cells (FRTL KiKi). (
Next, miR-10b-5p expression (anti-miR-10b-5p) was inhibited in irradiated FRTL KiKi cells in order to assess its role on cell proliferation and viability. Interestingly, the transfection of anti-miR-10b-5p stimulated the growth rate of irradiated FRTL KiKi cells in comparison with scramble (Fig. 7E). In agreement with that, the inhibition of miR-10b-5p consistently increased the relative number of viable cells compared to scramble (around 30%) (Fig. 7F). Taken together, these results indicate that miR-10b-5p sensitizes FRTL KiKi cells to IR.
MiR-10b-5p promotes radiosensitivity in human ATC cell line (8505c)
ATCs exhibit an aggressive behavior and are refractory to most cancer therapies, including radiotherapy (14). Thus, the study considered whether miR-10b-5p could also sensitize ATC cells to IR. First, the expression of miR-10b-5p was screened in a panel of human thyroid carcinoma cell lines. The 8505c cell line was chosen because (i) these cells are derived from human ATC, and (ii) they express the lowest levels of miR-10b-5p in comparison with all tested human thyroid carcinoma cell lines (Fig. 8A). Therefore, miR-10-5p was overexpressed in these cells (Fig. 8B). It was observed that miR-10b-5p increases the proliferation rate of 8505c cells (Fig. 8C). Subsequently, it was found that miR-10b-5p attenuates the growth rate (Fig. 8D) and viability (16%) (Fig. 8E) of irradiated 8505c cells in comparison with cells transfected with SC. Therefore, these data clearly indicate that miR-10b-5p induces radiosensitivity in the 8505c cell line.
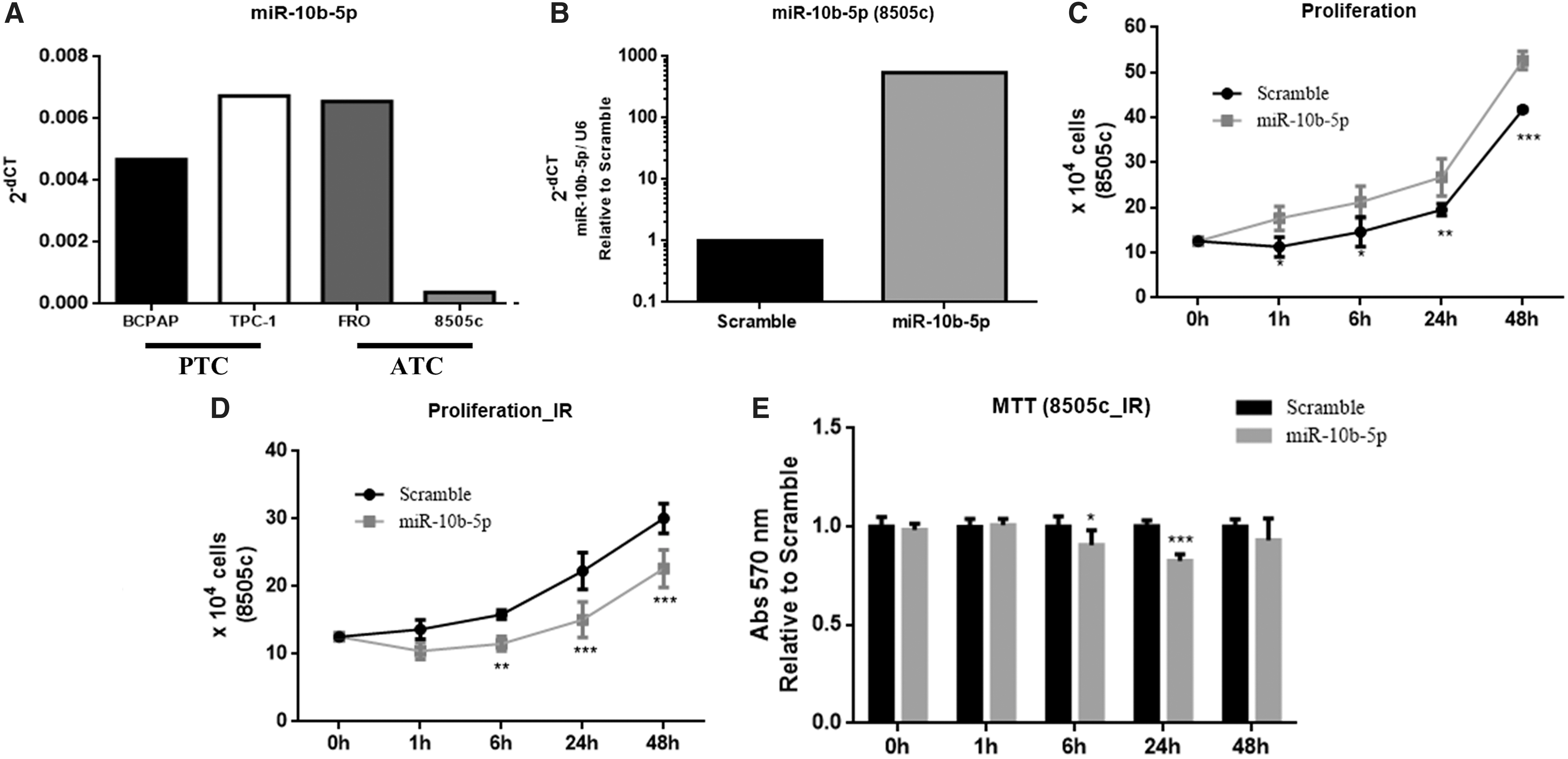
MiR-10b-5p increases proliferation and sensitizes the human anaplastic thyroid carcinoma cell line 8505c to IR. (
Discussion
Epigenetic mechanisms such as miRNA expression profiles have been reported as stress sensors involved in radiation-induced responses (13). In this context, this study investigated the influence of IR, a well-known risk factor for PTC development (6) (7), on differential global expression miRNA in the differentiated thyroid cell line FRTL-5 CL2 (21). This analysis revealed the induction or the repression of distinct miRNAs in irradiated FRTL-5 CL2 cells, depending on the exposure time. It was found that seven miRNAs downregulated one hour post irradiation, when FRTL-5 CL2 cells have a peak of γH2AX modification, as previously demonstrated (25), whereas 10 miRNAs were significantly upregulated by IR after six hours, when most of the DSB lesions were repaired (25). Consistently, gene ontology analysis demonstrates that most of the altered miRNAs belong to a class of “cellular response to stress” or “gene silencing by miRNA,” targeting DSB repair genes. Then, the study focused on miR-10b-5p and miR-199a-3p because these miRNAs were deregulated by IR at both one and six hours after irradiation and are described to have an oncogenic (23) and a tumor suppressor (26) activity in thyroid cancer, respectively.
Since the HR pathway is critical for DSB repair in thyroid cells, preventing RET/PTC rearrangement (35), the role of these miRNAs in DSB repair was investigated. It was found that miR-199a-3p and miR-10b-5p increase IR-induced DSB, indirectly measured by γH2AX levels (34). Since HMGA2 activates ATR in cancer cell lines (36), basal lower protein levels of ATR might be related to the delayed DSB repair in miR-199a-3p-overexpressing cells. Conversely, the downregulation of Dicer1 could account for the reduced levels of activated ATM and ATR, as well as the increased γH2AX levels in FRTL-5 CL2 cells transfected with miR-10b-5p precursor oligonucleotide. Indeed, DICER1 plays a central miRNA biogenesis (31), and it is also crucial for DSB repair signaling and ATM phosphorylation in eukaryotes (37 –39).
Interestingly, it has also been reported that defects in HR are related to radiosensitivity in cancer cells, predisposing them to accumulate unrepaired DSB, a lethal type of DNA lesion (40). Accordingly, the overexpression of miR-10b-5p slightly attenuated cell viability and compromised the growth of normal and neoplastic irradiated thyroid cell lines. These effects are likely due to the deficiency in HR activation after IR exposure, resulting in DSB accumulation, even though a role of the increased growth rate induced by miR-10b-5p overexpression cannot be excluded. Apart from that, IR exposure independently affects mitochondrial function and DNA. Although the MTT assay has classically been used to measure cell survival after irradiation (41), it has been reported that high doses of radiation (>1 Gy) impacts the MTT assay by affecting the dehydrogenase activity, and thus the results obtained with MTT assays are less reliable that those obtained by using cell counting after trypan blue staining (42,43). These results appear in contrast with the radioresistance promoted by miR-10b-5p in glioblastoma cells (44), suggesting that the effects of miR-10b-5p on cell sensitivity to IR is dependent on the cellular context, as already reported for other miRNAs (45), such as miR-205 that induces radioresistance in nasopharyngeal carcinoma cell lines (46) but sensitizes breast cancer cells to IR (47).
It is noteworthy that some of the findings might have clinical relevance in the understanding of the mechanisms involved in thyroid carcinogenesis, and also for the development of new strategies for the therapy of ATC. The findings highlight the targets of the miR-199a-3p (Lin28b) and miR-10b-5p (Dicer1) that are upregulated and downregulated, respectively, in human thyroid carcinomas. The study demonstrates that miR-199a-3p targets Lin28b, leading to a reduced expression of HMGA2, and it is speculated that the overexpression of let-7b could be involved in this phenomenon. Accordingly, this study reports that let-7 decreases RAS expression and thus compromises MAPK activation, which is fundamental for thyroid cell proliferation (48). Therefore, miR-199a-3p downregulation in thyroid carcinomas would lead to an increased expression of HMGA2, thereby contributing to cancer progression. Similarly, the study reports that miR-10b-5p targets the 3′-UTR region of Dicer1 mRNA, repressing its translation, resulting in a reduction of its protein levels, a mechanism that has already been associated with thyroid cell transformation (32,33). Moreover, the ability of both miRNAs to modulate IR-induced DSB may account for the reduced ability of thyroid cancer cells in DSB repair when these miRNAs are deregulated. Furthermore, the study reports an interesting role of miR-10b-5p in promoting the radiosensitivity of thyroid cancer cells, and thus it is hypothesized that the induction of miR-10b-5p-overexpression in malignant thyroid neoplasias, in particular ATCs, which are refractory to radiotherapy, might result in a better response to radiotherapy. Interestingly, recent studies provide reasonable perspectives of a therapy based on miRNA restoration. Indeed, liposomal encapsulation technology and biodegradable polymers delivery systems are used to deliver therapeutic miRNAs in vivo (49). For example, intranasal administration of let-7 inhibited lung tumor growth in Kras-mutant mice (50). Moreover, the restoration of miR-34 by infusion and the intravenous administration of miR-16 are in a Phase I clinical trial with patients with advanced solid tumors (49). The results of miR-10b-5p-overexpressing ATC cells are in agreement with the biphasic expression levels of miR-10b-5p in irradiated FRTL5 cells. Since miR-10b-5p stimulates thyroid cell proliferation, it is expected that miR-10b-5p expression levels are reduced at the first hour after irradiation, when cells stop proliferating in order to repair DNA damage. Once the DNA damage is repaired, six hours after irradiation, miR-10b-5p is upregulated in order to recover their proliferation. Since miR-10b-5p overexpression stimulates thyroid cell proliferation and negatively affects DNA repair efficiency, cells that overexpress this microRNA might be more susceptible to IR.
In conclusion, the results reported here demonstrate that miR-199a-3p and miR-10b-5p, that respectively target Dicer1 and Lin28b, are deregulated by IR and compromise DSB repair efficiency of thyroid cells by affecting the activation of ATM and ATR, hence predisposing cells to accumulate DSB after X-ray exposure (Fig. 9). Moreover, the data indicate that miR-10b-5p promotes radiosensitivity in a cell line, derived from ATC known to be refractory to radiotherapy, suggesting that the restoration of miR-10b-5p expression could form a new approach in the treatment of ATC.
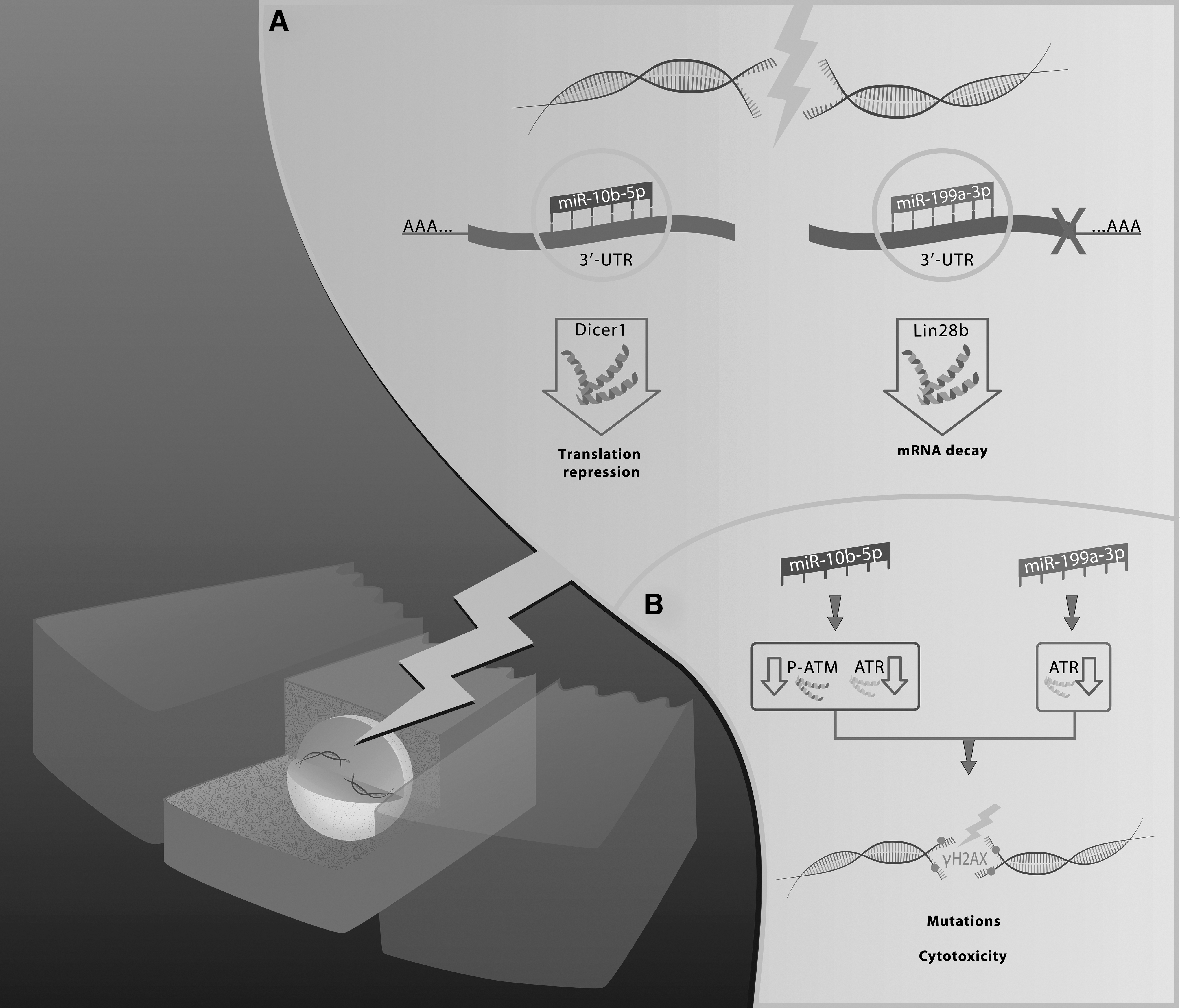
Schematic representation of the findings in this study. (
Footnotes
Acknowledgments
This study has been supported by grants from CNR Flagship Projects (Epigenomics-EPIGEN) and the Associazione Italiana per la Ricerca sul Cancro (AIRC IG 11477). R.C.C.P. was the recipient of a fellowship CAPES for the program “Ciencias sem Fronteiras.” A.F. was granted with a Special Visiting Researcher (PVE) fellowship sponsored by CAPES. Thanks to Simone Eboli for the graphic art.
Author Disclosure Statement
The authors have declared no conflict of interest.