
Abstract
Background:
The secretion of pituitary hormones oscillates throughout the 24-hour period, indicating that circadian clock-mediated mechanisms regulate this process in the gland. Additionally, pituitary hormone synthesis has been shown to be altered in hypo- and hyperthyroidism. Although thyroid hormones can modulate the other peripheral clocks, the interaction between thyroid hormone levels and circadian clock gene expression in the anterior pituitary has yet to be elucidated.
Methods:
Male Wistar rats were divided into three groups: control, hypothyroid, and hyperthyroid. Following the experimental procedures, animals were euthanized every three hours over the course of a 24-hour period. The anterior pituitary glands were excised and processed for mRNA expression analysis by quantitative reverse transcriptase polymerase chain reaction. One- and two-way analysis of variance as well as cosinor analysis were used to evaluate the time-of-day-dependent differential expression for each gene in each experimental group and their interactions.
Results:
Hyperthyroidism increased the mRNA expression of core clock genes and thyrotrophic embryonic factor (Tef), as well as the mesor and amplitude of brain and muscle Arnt-like protein-1 (Bmal1) and the mesor of nuclear receptor subfamily 1 (Nr1d1) group D member 1, when compared to euthyroid animals. Hypothyroidism disrupted the circadian expression pattern of Bmal1 and period circadian regulator 2 (Per2) and decreased the mesor of Nr1d1 and Tef. Furthermore, it was observed that the pituitary content of Dio2 mRNA was unaltered in hyperthyroidism but substantially elevated in hypothyroidism during the light phase. The upregulated expression was associated with an increased mesor and amplitude, along with an advanced acrophase. The gene expression of all the pituitary hormones was found to be altered in hypo- and hyperthyroidism. Moreover, prolactin (Prl) and luteinizing hormone beta subunit (Lhb) displayed circadian expression patterns in the control group, which were disrupted in both the hypo- and hyperthyroid states.
Conclusion:
Taken together, the data demonstrate that hypo- and hyperthyroidism alter circadian clock gene expression in the anterior pituitary. This suggests that triiodothyronine plays an important role in the regulation of pituitary gland homeostasis, which could ultimately influence the rhythmic synthesis and/or secretion of all the anterior pituitary hormones.
Introduction
Organisms are exposed to a variety of physiological demands and environmental stresses throughout the day, and must adjust accordingly to maintain homeostasis. For example, circadian rhythms in biological processes have been widely observed during sleep/wake and feeding/fasting cycles, metabolic and body temperature regulation, reproductive behavior, hormone synthesis and secretion, as well as hormone receptor affinity and abundance, among others. These adaptive features are vital for proper cell functioning and survival of the organism (1).
In mammals, in addition to the master circadian clock, located in the hypothalamic suprachiasmatic nucleus, other regions of the brain and tissues (i.e., the liver, kidney, and heart) possess intrinsic circadian clocks that respond to environmental stimuli (i.e., light/dark periods, feeding/fasting cycles, exercise-induced stress and nutrition) (2 –5). The molecular mechanisms involved in the regulation of the central and peripheral circadian clocks consist of an autoregulation of core clock genes through feedback loops, with a periodicity of about 24 hours (6,7).
The brain and muscle Arnt-like protein-1 (BMAL1)/circadian locomotor output cycles kaput (CLOCK) heterodimer is the positive core clock component that activates the transcription of many genes through its binding to the enhancer box (E-box) sequence near to their promoter regions, including the negative core clock elements period circadian regulators 1 and 2 (Per1/2) and cryptochrome circadian regulators 1 and 2 (Cry1/2), as well as clock-controlled genes (CCG). The PER/CRY complex inhibits the activity of BMAL1/CLOCK through both transcriptional and post-transcriptional mechanisms (1,6,8). A secondary regulatory loop includes the retinoic acid orphan-related receptor alpha (RORa) enhancer and the nuclear receptor subfamily 1 group D member 1 (NR1D1) repressor, which ultimately modulate Bmal1 transcription (9,10). In addition to transcriptional modulation, post-transcriptional and post-translational regulation of clock genes and CCG has also been demonstrated (2,11,12).
The pituitary gland possesses a functional intrinsic circadian clock that is involved in the regulation of many cellular functions, including hormone and transcriptional processes, as well as cellular growth and death (13). The anterior pituitary is composed of five different endocrine cell types whose hormonal products directly regulate reproduction, growth, development, and metabolism. Pituitary hormone secretion is mainly under the control of hypothalamic factors and fluctuates over a 24-hour period (14 –19). Furthermore, the in vitro expression of prolactin (Prl) and growth hormone (Gh) mRNA presents a circadian pattern (20), while the basal expression of thyrotropin beta (Tshb) can be regulated negatively by NR1D1 (21) or positively by thyrotrophic embryonic factor (Tef), a transcription factor mainly expressed during embryogenesis, which in turn is controlled by the circadian clock (13,22). Thus, all these data indicate that there is a correlation between pituitary function and its circadian clock.
Recently, it was shown that triiodothyronine (T3) is an important modulator of pituitary function. This hormone not only acts on the hypothalamus–pituitary–thyroid (HPT) axis through the inhibition of thyrotropin (TSH) synthesis/secretion (23 –26), but also modulates the synthesis and secretion of GH, luteinizing hormone (LH), and follicle-stimulating hormone (FSH) by transcriptional and post-transcriptional mechanisms (27 –30). Furthermore, in the pituitary gland, intracellular T3 availability is mainly regulated by the activity of deiodinase type 2 (DIO2), which is controlled by the clock (31,32).
Taken together, these findings led to the hypothesis that T3 could directly and/or indirectly alter the pituitary clock by perturbing the patterns of pituitary hormone synthesis and secretion. This hypothesis is supported by a recent observation, which demonstrated that the circadian clock in rat hearts is modulated by T3 (33,34). Due to the fact that the pituitary gland plays a central role in endocrine physiology and that thyroid disorders represent the second most prevalent endocrinopathy in the world, the present study sought to evaluate the repercussions of hypo- and hyperthyroidism on the expression of the pituitary circadian clock and the daily expression profiles of pituitary hormones and CCG involved in pituitary function.
Methods
Animals
Male Wistar rats, 45–50 days old and weighing 180–200 g, were obtained from the animal facility at the Institute of Biomedical Sciences (Animal Facility Network at the University of São Paulo, São Paulo, Brazil). The animals were kept on a 12-hour/12-hour light/dark cycle (light phase: lights on at 6:00am = zeitgeber time ZT 0; dark phase: red filter Kodak 1A, 0.5–1 lux) in a temperature-controlled room (25 ± 2°C) and had access to food and water ad libitum. The experimental protocol was approved by the local Committee for Animal Experimentation and followed the National Guidelines for the Ethical Use of Animals in Research (#81, 21-03).
Treatments and experimental designs
The animals were divided into three experimental groups: control, hypothyroid, and hyperthyroid. Hypothyroidism was induced during puberty (45–50 days old) by surgical thyroidectomy after being deeply anesthetized with ketamine and xylazine. Following the procedure, the rats were given 0.03% methylmercaptoimidazole (MMI) and 4.5 mM calcium chloride in their drinking water for 20 days. Hyperthyroidism was induced at the end of puberty (55–60 days old) by intraperitoneal injections of T3 (0.75 μg/100 g), twice a day, at ZTs 2 and 14, for five consecutive days. The control animals underwent a sham operation, were not subjected to MMI treatment, and received saline injections at the same ZTs as the hyperthyroid group.
Young adult rats (60 days old) were euthanized every three hours over a 24-hour period, and the pituitaries were rapidly excised, immediately frozen in liquid nitrogen, and stored at −80°C prior to subsequent analysis. At least five animals were included at each ZT per experimental group.
Quantitative reverse transcriptase polymerase chain reaction
Total RNA was extracted from the pituitary gland using Trizol according to the manufacturer's instruction and was quantified spectrophotometrically using Gen v1.10 software and an Epoch™ Microplate Spectrophotometer coupled to a Take3™ Multi-Volume Plate (BioTek, Winooski, VT). Afterwards, 1 μg of total RNA was used for first-strand cDNA synthesis (reverse transcriptase [RT]) using MMLV-RT (Promega, Madison, WI). The reverse transcription products were diluted in DEPC water and submitted to real-time quantitative polymerase chain reaction (PCR; 0.5 ng/reaction) using Platinum® Syber® qPCR Supermix-UDG and specific oligonucleotides directed toward the candidate genes (Table 1).
RT-qPCR Primer Sequences
RT-qPCR, quantitative reverse transcriptase polymerase chain reaction.
The PCR conditions were as follows: 95°C for 2 min followed by 40 cycles of 95°C for 15 s, 60°C for 1 min, and 72°C for 20 s. The reactions were performed in a Corbett Research system (Corbett Life Sciences, Mortlake, Australia). Ct values were recorded for each gene, and the results were normalized using an internal control, ribosomal protein L19 (Rpl19). The efficiency and slope values were close to the optimal values required for the ΔΔCT analysis for all investigated candidate genes (35 –37). No differences in the Rpl19 expression between the groups/ZTs were observed using 2(−ΔCt) analysis (38) (data not shown).
Total T3 and TSH serum concentration measurements
The efficiency of the treatments was evaluated by measuring the total T3 and TSH serum concentrations by radioimmunoassay and with the specific rat pituitary panel from Milliplex (MAP Rat Pituitary Magnetic Bead Panel; Milliplex, Billerica, MA), respectively. For this, trunk blood was collected, and a pool of samples obtained during 24 hours from each of the experimental groups was assessed.
Statistical analysis
The data are presented as the mean ± standard error of the mean. The results were tested for normality using the D'Agostino-Pearson omnibus normality test. The T3 and TSH serum concentrations were evaluated using the Kruskal–Wallis test, followed by Dunn's multiple comparisons test. For the gene expression analysis, each temporal series was tested using one-way analysis of variance (ANOVA) to evaluate the influence of time, and two-way ANOVA was employed to verify the influence of time and treatment, followed by Bonferroni's post hoc analysis for pair-wise comparisons using GraphPad Prism v7 (GraphPad Software, Inc., San Diego, CA). The genes/groups that presented statistically significant one-way ANOVA results for the main effect of time were subjected to a second mathematical and statistical procedure to investigate the presence of a daily 24-hour rhythm (39). The theoretical cosine curve fit (cosinor method) was applied to each temporal series using the least-square calculation. The periodic fit was considered significant when p < 0.05. Thus, the mesor, amplitude, and acrophase were calculated for each group/gene (40) and compared by Student's t-test or one-way ANOVA, accordingly.
Results
Validation of the experimental procedures
The effectiveness of the treatments was confirmed by the TSH and total T3 serum concentrations from a pool of samples obtained at different ZTs (Table 2). As expected, the hypothyroid group presented a marked increase in TSH serum concentration and reduced total T3 serum levels when compared to the control group. In contrast, the hyperthyroid group displayed a reduced TSH serum concentration and an increased total T3 serum concentration when compared to both the control and hypothyroid groups.
TSH and Total T3 Serum Concentrations
Average ± SEM (n) from a pool of samples obtained from control, hypothyroid, or hyperthyroid rats. Kruskal–Wallis test followed by Dunn's multiple comparisons test. * p < 0.05 vs. control; # p < 0.05 vs. hypothyroid.
TSH, thyrotropin; T3, triiodothyronine; SEM, standard error of the mean.
Thyroid dysfunction altered the expression of core clock genes in the pituitary gland
A summary of the daily gene expression of core clock genes in the pituitary is shown in Figure 1. Significant changes in the expression of all of the core clock genes were observed in animals with thyroid dysfunction (hypo- or hyperthyroidism), and the main effect of time was observed for all of the investigated core clock genes in the pituitary gland, with the exception of Clock (Fig. 1B). Moreover, there were statistically significant interactions between the main effects of treatment and time for Bmal1, Per2, Nr1d1, and RAR-related orphan receptor A (Rora).

Clock gene expression in the pituitary in hypo- and hyperthyroid states. Control rats, black line; hypothyroid rats, red line; hyperthyroid rats, blue line. (
As shown in Figure 1A, hyperthyroid animals displayed significant increases in Bmal1 expression at ZTs 0/24, 3, and 12 when compared to control animals and at ZTs 15, 18, and 21 when compared to hypothyroid animals. The expression of Clock was higher in the hyperthyroid group when compared to both the control and hypothyroid groups at ZTs 0/24 and 15, and was also shown to be increased at ZTs 3, 12, and 21 when compared to hypothyroid animals (Fig. 1B). The Per2 mRNA content was elevated in the pituitary of hyperthyroid animals at ZTs 0/24 and 15 when compared to either control or hypothyroid animals, and was only significant at ZTs 12 and 18 when compared to the hypothyroid group. In addition, when compared to controls, hypothyroidism significantly reduced Per2 expression at ZT 12 (Fig. 1C). With regards to Cry1 expression in rat pituitaries, the hyperthyroid group presented increased gene expression levels at ZTs 12 and 15 when compared to the control and hypothyroid groups, respectively (Fig. 1D). The Nr1d1 transcript was increased at ZTs 0/24 and 3 in the hyperthyroid group when compared to the hypothyroid group. In contrast, the expression of this gene was decreased in the hypothyroid group at ZTs 6 and 12 when compared to the control group (Fig. 1E). The Rora mRNA content was elevated in hyperthyroid animals at ZTs 0/24 and 15 when compared to control and hypothyroid groups (Fig. 1F).
Thyroid disorders induced marked changes in the circadian rhythmicity of clock genes in the pituitary
The cosinor analysis of the core clock genes Bmal1, Per2, and Nr1d1 in the pituitary is presented in the Figure 2. In the control group (black lines), a circadian pattern was observed for Bmal1, Per2, and Nr1d1 gene expression (Fig. 2A–C, respectively). Hypothyroidism abolished the circadian expression patterns of Bmal1 (Fig. 2A) and Per2 (Fig. 2B) and reduced the amplitude and mesor of Nr1d1 expression (Fig. 2C, red line) when compared to control values. In contrast, the hyperthyroid condition increased the mesor of Bmal1 and obliterated the Per2 circadian pattern (Fig. 2A). With regards to Nr1d1 mRNA expression, hyperthyroidism increased the mesor, reduced the amplitude, and phase advanced the acrophase when compared to the control group (Fig. 2C, blue line). The control group presented no observable alterations in the mRNA expression of Clock or Cry1 over the experimental period, and no rhythmicity was detected for Rora mRNA expression (Supplementary Table S1).

Cosinor rhythmic analysis of clock genes in the pituitary. Bmal1 (
Daily expression patterns of pituitary hormones in hypo- and hyperthyroidism
The expression of pituitary hormone transcripts was evaluated over a 24-hour period under hypo- and hyperthyroid conditions (Fig. 3). The gene expression of all the pituitary hormones was significantly altered with regards to the main effect of treatment (hypo- and hyperthyroid), while the main effect of time was only found to be statistically significant for proopiomelanocortin (Pomc). Furthermore, interactions were detected between the main effects of treatment and time for Gh, luteinizing hormone beta (Lhb), and Pomc.
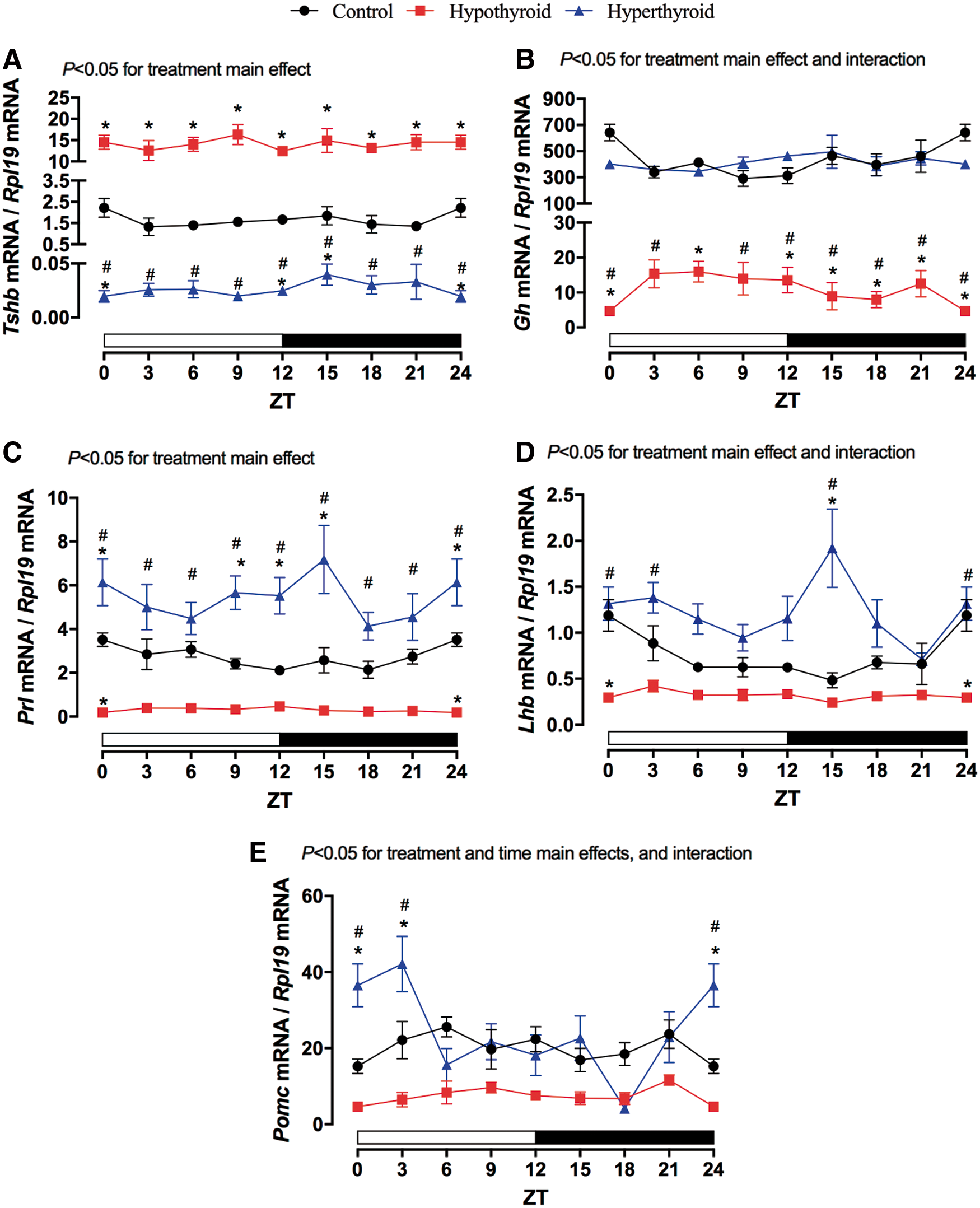
Gene expression of pituitary hormones. The mRNA content of pituitary hormones of the control group (black line), hypothyroid group (red line), and hyperthyroid group (blue line). (
The pair-wise analysis showed that the hypothyroid group had markedly increased Tshb gene expression levels (Fig. 3A, red line), while hyperthyroidism strongly reduced the Tshb levels at all time points (Fig. 3A, blue line). The Gh mRNA content was reduced in hypothyroid rats throughout the analyzed period (Fig. 3B, red line), but no significant differences were observed under hyperthyroid conditions (Fig. 3B, blue line). The hyperthyroid group displayed increased Prl expression levels (Fig. 3C, blue line), and a significant reduction was observed in the hypothyroid group at ZT 0/24 (Fig. 3C, red line) when compared to the controls (black lines). In addition, the pituitaries of hypothyroid rats contained less Lhb mRNA at ZT 0/24 when compared to control animals (Fig. 3D, red line). On the other hand, hyperthyroidism promoted significant increases in Lhb mRNA expression at ZT 15 when compared to both the control and hypothyroid groups, as well as at ZTs 0/24 and 3 when compared only to the hypothyroid group (Fig. 3D, blue line). Lastly, Pomc gene expression was augmented at ZTs 0/24 and 3 in the hyperthyroid state when compared to the control condition (Fig. 3E).
Prl and Lhb expression in the pituitary presented a circadian rhythmicity pattern that is altered by thyroid hormone levels
The expression of Prl and Lhb in the control group presented a circadian expression pattern (Fig. 4). Hypothyroidism reduced the mesor and amplitude and induced a phase delay in the acrophase of Prl mRNA expression when compared to the control condition (Fig. 4A). As shown in Figure 4B, it was also found that both hypo- and hyperthyroidism abolished the circadian rhythmicity pattern of Lhb mRNA expression in the rat pituitary.

Cosinor rhythmic analysis of pituitary hormone expression. Prl (
With regards to other pituitary hormones, Tshb mRNA content in all the groups remained unchanged throughout the experimental period (no statistical differences in one-way ANOVA), and any differences were due exclusively to the treatment. Furthermore, the cosinor analysis failed to detect any circadian patterns associated with the mRNA content of Gh and Pomc (see Supplementary Table S1 for additional details).
Thyroid dysfunction altered the expression and circadian pattern of clock-controlled genes
The effects of hypo- and hyperthyroidism on Dio2 and Tef expression in the rat pituitary gland were also evaluated during the 24-hour period (Fig. 5). Overall, the expression of both Dio2 and Tef was significantly altered in terms of the main effects of treatment and time. However, an interaction was only detected for Dio2 mRNA content. Hypothyroidism resulted in increased Dio2 mRNA content, with levels two- to threefold higher during the light phase (ZTs 3–12; Fig. 5A, red line). In contrast, the hyperthyroid group did not display any observable alterations in Dio2 gene expression when compared to the control group (Fig. 5A blue line). With regards to Tef expression, hyperthyroid animals presented increased levels at ZTs 0/24, 3, and 12 when compared to control animals, and increased levels at ZTs 0/24, 3, 12, 15, and 18 when compared to hypothyroid animals (Fig. 5B).

Gene expression of clock controlled genes and cosinor rhythmic analysis in the pituitary. The mRNA content of clock controlled genes of the control group (black line), hypothyroid group (red line), and hyperthyroid group (blue line). (
Additionally, the cosinor rhythmic analyses of Dio2 and Tef mRNA expression levels revealed that in the pituitary of the control and hypothyroid groups, the expression of these genes fits a circadian rhythmicity pattern (Fig. 5C and D, respectively). Hypothyroidism was found to increase the mesor and amplitude of Dio2 and to reduce these parameters in Tef expression. Additionally, hypothyroidism advanced the acrophase of both transcripts when compared to the control group, while hyperthyroidism abolished their circadian expression pattern (Supplementary Table S1).
Discussion
In this study, the repercussions of hypo- and hyperthyroidism on the pituitary circadian clock, CCG, and daily pituitary hormone expression profiles were investigated. Hypothyroidism was confirmed by the presence of higher levels of TSH and reduced total T3 concentrations in the serum, while the opposite was observed in hyperthyroid rats. Overall, the expression of all investigated core clock components, pituitary hormones, and CCG were markedly altered in response to hypo- and hyperthyroidism, and their circadian oscillations were strongly changed according to the thyroid hormonal levels.
Recently, it was shown that these thyroid conditions alter the expression of circadian clock genes in the heart, with more prominent effects observed in the hypothyroid state (34). Similar to the heart, the pituitary is an important target of transcriptional, posttranscriptional, and posttranslational thyroid hormone–mediated regulation. The consequences of thyroid disorders on pituitary hormone synthesis/secretion have been investigated, mainly during the light phase, and were shown to modulate the synthesis and secretion of TSH (23,24,26), GH (29,41), LH and FSH (27,28), as well as others. Moreover, posttranscriptional mechanisms regulating Gh mRNA poly(A) tail length have been shown to be related to daily variations in GH synthesis (42). Based on the fact that there are a wide range of effects observed in all pituitary endocrine cell types due to imbalances in thyroid hormone levels and that the pituitary hormone secretion displays a circadian rhythm, it was hypothesized that these effects could be due to a direct and/or indirect modulation of the pituitary clock by thyroid hormones.
The results show that the gene expression of the core clock, CCG, and pituitary hormones were altered in hypo- and hyperthyroidism. The expression levels of Bmal1, Per2, and Nr1d1 oscillated with a period of 24-hour in euthyroid animals (control group), while Clock, Cry1, and Rora did not present a circadian pattern in the pituitary gland—a result that is consistent with a previous study (20). The absence of thyroid hormones (hypothyroid group) reduced the expression of core clock genes when compared to the control group, abolished the circadian pattern of Bmal1 and Per2, and reduced the mesor and amplitude of Nr1d1. On the contrary, hyperthyroidism increased the expression of all core clock genes at distinct ZTs, abolished the Per2 circadian rhythmicity, increased the mesor and amplitude of Bmal1, and reduced the amplitude and phase advanced Nr1d1 expression.
Previous studies have provided evidence for a correlation between the intrinsic clock and pituitary function (13). In fact, it was demonstrated that NR1D1 is involved in recruiting Nuclear Corepressor 1 (NCOR1), which leads to transcriptional repression of basal Tshb expression by a mechanism independent of thyroid hormones (21). While the molecular mechanism involved in the circadian secretion of pituitary hormones has yet to be elucidated, it is almost certainly related to the circadian clock of the pituitary and can be modulated by several neuroendocrine factors (i.e., hypothalamic hormones, glucocorticoids, and thyroid hormones).
In other words, hypo- and hyperthyroidism alter or abolish the pituitary clock rhythmicity, which could impact pituitary metabolism and function. As observed for the core clock genes, the expression of all pituitary hormones and CCGs was altered by the treatments. As expected, hypothyroidism increased Tshb mRNA content throughout the 24-hour experimental period and reduced the expression levels of Gh, Prl, Lhb, and, apparently, Pomc. In contrast, hyperthyroidism reduced Tshb mRNA content, increased the Prl expression at all time points, and increased the Lhb and Pomc mRNA content at certain ZTs. Additionally, the hyperthyroid animals did not display any observable alterations in Gh expression when compared to control animals.
It is well known that the endogenous levels of thyrotropin-releasing hormone (TRH) are elevated in the hypothyroid state. TRH is the main regulator of TSH synthesis/secretion and also increases PRL expression/secretion in a sex-specific manner. For example, female rats have higher levels of TRH in the anterior pituitary compared to male rats (43), and PRL release has been shown to be important during the first days of lactation (44). On the other hand, in male rats, the endogenous TRH synthesis/secretion is not required for the physiological control of PRL secretion (45), which is in agreement with the current data and previous studies (46,47), which showed a reduction of Prl and Gh expression in the hypothyroid condition. In addition to the sexual dimorphism of Prl modulation by TRH, a central inhibitory action of TRH on prolactin secretion through the activation of dopaminergic pathways in the arcuate hypothalamic nucleus was previously described in rats (48,49). Besides dopamine inhibiting PRL secretion, it also suppresses the adenylyl cyclase activity and reduces the inositol phosphate metabolism, resulting in the suppression of Prl gene expression (50). Thus, it is suggested that the elevated levels of TRH increase the dopaminergic inhibitory effect on Prl expression and secretion in hypothyroid male rats.
Furthermore, an important ontogenetic modulation of Lhb expression by thyroid hormones was observed. Previous studies have shown that the induction of hypo- and hyperthyroidism in adulthood promotes an increase and a decrease in Lhb mRNA content, respectively (27,28). On the contrary, the induction of hypothyroidism during gestation, postnatal development, and/or puberty (experimental model of the present study) reduces LH expression/secretion, which can be effectively reversed with T3 treatment (51,52). All these data indicate that thyroid hormones play a critical role in the maturation of the hypothalamus–pituitary–gonads axis, and that any alterations in these hormone levels during early development could lead to different reproductive disorders later in life.
With regards to the rhythmic expression patterns of pituitary hormones, Prl and Lhb showed a circadian oscillation pattern. On the other hand, the levels of Pomc and Gh mRNA content varied over the 24-hour period, but the cosinor analysis failed to detect any circadian oscillations. The Tshb gene expression in control rats did not present a daily pattern contrasting in vitro experiments using TαT1 cells synchronized by serum shock challenge (21). These data suggest that the circadian oscillations in TSH, GH, and ACTH secretion observed in several species could be modulated by posttranscriptional or posttranslational events or even by the activation of direct pathways, which can regulate the secretion rates independently of transcriptional mechanisms. It is highly plausible that all these steps might be modulated by hypothalamic factors (15,17 –19).
Hypothyroidism reduced the amplitude and mesor of Prl expression and induced a delay in its acrophase at around 7.5 hours, while the circadian oscillation of Lhb mRNA content was abolished in the absence of thyroid hormones. Additionally, hyperthyroidism suppressed the rhythmicity for both Prl and Lhb. Thus, it is becoming clear that thyroid disorders can alter the rhythmicity of the reproductive axis through direct or indirect effects on the pituitary clock, which can in turn influence the gonadotroph and lactotroph activities.
The expression of Dio2 and Tef is under the control of the circadian clock and presents a circadian oscillation in the pituitary gland of euthyroid rats. However, in the hypothyroid condition, both transcripts were found to be phase advanced. Hypothyroidism also increased the expression and amplitude of Dio2 (which converts thyroxine [T4] to T3) mainly during the light phase, which could ensure T3 availability for the active phase of the animals (dark phase). In hypothyroid animals, the expression of Tef showed a reduction in the mesor and amplitude. In the hyperthyroid animals, the circadian expression patterns of Dio2 and Tef were abolished. The observed alterations in the expression patterns of both of these CCGs can be directly related to the presence or absence of thyroid hormones modulating the transcription of these genes or indirectly through thyroid hormone-mediated modulation of the pituitary circadian clock.
In brief, it has been shown that the absence or excess of thyroid hormones can significantly alter pituitary clock function, changing the circadian patterns of CCG and pituitary hormone gene expression. Similar modulation of the circadian clock was shown in rat hearts, which led to the hypothesis that the peripheral clocks of other thyroid hormone targeted tissues might be altered under conditions of hypo- and/or hyperthyroidism.
It is well known that the HPT axis presents a dynamic and adaptive response to several illnesses and/or conditions. The adaptive characteristic of thyroid homeostasis to critical illness is referred to as low-T3 syndrome, nonthyroidal illness syndrome, euthyroid sick syndrome, or thyroid allostasis in critical illness, tumors, uremia, and starvation (53). In starvation, for instance, a differential modulation between central (increased) and peripheral (reduced) deiodinase activities results in lower TRH and TSH secretion levels and reduced concentrations of serum T3 and T4. In the same condition, other studies have also reported a marked reduction in LH, GH, and PRL synthesis and secretion (54 –56), as well as a reduction in the amplitude of GH pulses and the duration/length of GH secretory episodes (57).
The present findings suggest that the adjustments that the HPT axis makes in an attempt to ensure thyroid homeostasis during critical illness could interfere with the circadian clock of the pituitary gland, triggering a chronodisruption of the hypothalamic–pituitary axes and consequently their target tissues. Such a situation could directly promote reproductive, growth, and metabolic disorders, frequently observed in patients who present thyroid dysfunctions, therefore being extremely relevant in a clinical setting. Further studies to characterize the mechanisms by which thyroid hormones directly or indirectly regulate the clock in pituitary cells, as well as the circadian clock regulation on pituitary hormone synthesis and secretion, may provide a better understanding of the underlying mechanisms in the future.
Footnotes
Acknowledgments
The authors are grateful to Julieta H. Scialfa Falcão and Erika Lia Brunetto for the excellent technical assistance. This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP: 2013/05629-4) awarded to M.T.N. P.B.-S. is the recipient of a FAPESP fellowship (2014/25888-7 and 2016/24941-7). M.T.N. is the recipient of a Conselho Nacional de Pesquisa e Desenvolvimento (CNPq—309437/2017-2) fellowship and CAPES Foundation.
Author Disclosure Statement
The authors declare no conflict of interest.
Supplementary Material
Supplementary Table S1