
Abstract
Background:
The molecular basis of familial nonmedullary thyroid cancer (FNMTC) is still poorly understood, representing a limitation for molecular diagnosis and clinical management. In this study, we aimed to identify new susceptibility genes for FNMTC through whole-exome sequencing (WES) analysis of leukocyte DNA of patients from a highly informative FNMTC family.
Methods:
We selected six affected family members to conduct WES analysis. Bioinformatic analyses were undertaken to filter and select the genetic variants shared by the affected members, which were subsequently validated by Sanger sequencing. To select the most likely pathogenic variants, several studies were performed, including family segregation analysis, in silico impact characterization, and gene expression (messenger RNA and protein) depiction in databases. For the most promising variant identified, we performed in vitro studies to validate its pathogenicity.
Results:
Several potentially pathogenic variants were identified in different candidate genes. After filtering with appropriate criteria, the variant c.701C>T,
Conclusions:
WES analysis in one family identified SPRY4 as a likely novel candidate susceptibility gene for FNMTC, allowing a better understanding of the cellular and molecular mechanisms underlying thyroid cancer development.
Introduction
Thyroid cancer is the most common malignancy of the endocrine system (1). Although the majority of these tumors are sporadic, 3–9% of all follicular cell-derived thyroid tumors are familial, referred to as familial nonmedullary thyroid cancers (FNMTCs) (2). Approximately 95% of FNMTC cases are nonsyndromic (2,3). Nonsyndromic FNMTC is currently defined by the diagnosis of two or more first-degree relatives with differentiated thyroid cancer of follicular cell origin, without other predisposing hereditary syndrome, and in the absence of environmental causes (2,3). Compared with the sporadic forms of nonmedullary thyroid cancer (NMTC), FNMTC tends to occur more often at an early age (2). Papillary thyroid cancer (PTC) is the most common histologic subtype of FNMTC and sporadic NMTC (2,3). Up to 45–55% of the patients with FNMTC also have an increased rate of benign thyroid disease, including follicular adenoma and multinodular goiter (2). Several studies suggest a more aggressive clinical behavior of FNMTC, with a less favorable prognosis than sporadic forms. However, there are also studies reporting that the FNMTC is not more aggressive than the sporadic disease (4 –6).
Syndromic FNMTC has well-defined genotype–phenotype associations (5). These include several familial cancer syndromes such as Cowden syndrome, familial adenomatous polyposis, Carney complex, Werner syndrome, and DICER1 syndrome (7 –12).
It is thought that the inheritance pattern of FNMTC is autosomal dominant with incomplete penetrance and variable expressivity. Linkage analyses mapped several loci involved in nonsyndromic FNMTC genetic predisposition (2,13). The analysis of candidate genes, and the recent use of next-generation sequencing for whole exome and genome analyses, led to the identification of several susceptibility genes for FNMTC, such as NKX2.1 (14), DICER1 (15), SRGAP1 (16), FOXE1 (17), HABP2 (18), SRRM2 (19), RTFC (20), and, more recently, MAP2K5 (21) and MYO1F (22). In addition, studies conducted in patients with differentiated thyroid cancer and in patients with FNMTC, identified germline mutations in DNA repair genes (e.g., BRCA1/2, ATM, and CHEK2) (23 –25). Taken together, these findings support a genetic heterogeneity for FNMTC. Since most genes are mutated in single families or in a small fraction of the families, the etiology of FNMTC, as a distinct syndrome, continues to be poorly understood (4,5).
In this study, whole-exome sequencing (WES) analysis and functional studies, in a large family with FNMTC, provided evidence that a SPRY4 gene variant could play a role in FNMTC.
Materials and Methods
Ethics statement and patients
The collection of biological samples from all subjects involved in this study was performed after written informed consent. This study was approved by the Institutional Review Board (IRB) and the Ethical Committee of Instituto Português de Oncologia de Lisboa Francisco Gentil (IPOLFG).
The most informative family with FNMTC from our cohort [Family 2, Portuguese (European ancestry), previously reported (6,13,17,26,27)], followed in the Endocrinology Department from IPOLFG, was chosen for this study. In this family, benign and malignant thyroid lesions were the most common phenotypes, as follows: seven members were affected with NMTC (in some cases in concomitance with nodular goiter) (III.7, IV.2, IV.6, IV.8, V.3, V.8, and V.67) (Supplementary Table S1), and nine patients had nodular goiter only (I.2, III.3, IV.4, IV.13, IV.18, IV.25, IV.33, IV.60, and IV.63) (Fig. 1). In addition, of the seven cases affected with NMTC, five had benign colon lesions, and two had ovarian cancer, which may represent, or not, syndromic aspects related to the disease under study. Furthermore, several family members were affected with other types of cancer, such as bladder and testicular cancer. Biological sample information is described in the Materials and Methods section of Supplementary Data.
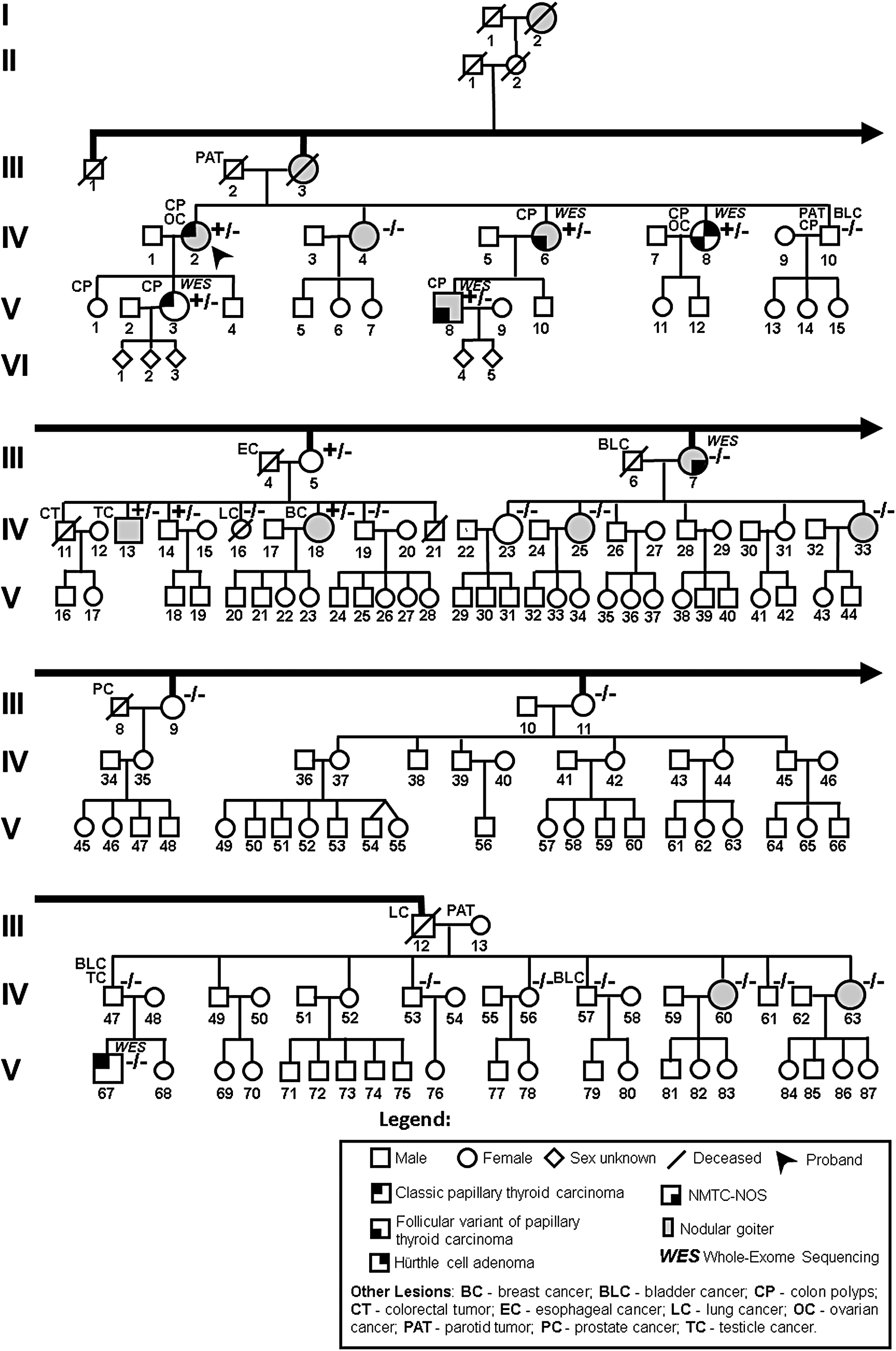
Pedigree of the family with FNMTC analyzed in this study. The carriers of the variant in the SPRY4 gene c.701C>T (
Probands of additional ninety-six FNMTC families were also included in this study. DNAs from 150 peripheral blood samples from Portuguese healthy controls (60 males and 90 females, average 63.9 years; supplied by Biobanco-iMM, Lisbon Academic Medical Center, Portugal) were also used.
DNA extraction from blood, tumor tissues, and cells
The details of DNA extraction are described in the Materials and Methods section of Supplementary Data.
Whole-exome sequencing
Genomic DNA samples obtained from peripheral blood leukocytes from six representative members of this family, affected with thyroid cancer (individuals III.7, IV.6, IV.8, V.3, V.8, and V.67), were used for WES. This analysis was performed by the DNAVision s.a. company (Charleroi, Belgium) in individual IV.6. WES analysis of individuals III.7, IV.8, V.3, V.8, and V.67 was performed by the Erasmus Center for Biomics (Netherlands). The details of WES and bioinformatic analyses are described in the Materials and Methods section of Supplementary Data.
Polymerase chain reaction
Polymerase chain reactions (PCRs) were performed using the Taq DNA polymerase (Invitrogen™, California, USA) protocol. Primers information is listed in Supplementary Table S2.
Sanger sequencing and mutational analysis
Sanger sequencing was used to validate and analyze segregation of the variants identified by WES in the family, and also to analyze these and other variants in tumors and cell lines. The details are described in the Materials and Methods section of Supplementary Data. Primers information is listed in Supplementary Table S2.
Cell culture
Three distinct cell lines were used in this study: a human PTC cell line (TPC-1), Rattus norvegicus normal thyroid cells (PCCL3), and Mus musculus embryo fibroblasts (NIH/3T3). The details of cell culture conditions are described in the Materials and Methods section of Supplementary Data.
Plasmid preparation, transfection, and transduction of cell lines
Cell lines NIH/3T3, PCCL3, and TPC-1 stably overexpressing the candidate gene SPRY4 were prepared. The details of plasmid preparation, transfection, and transduction of cell lines are described in the Materials and Methods section of Supplementary Data.
Cell viability assay
Cell viability over time was analyzed by trypan blue (Gibco®; Life Technologies, Paisley, UK) exclusion assay. The detailed protocol is described in the Materials and Methods section of Supplementary Data.
Cell migration analysis
Cell migration was analyzed by wound healing assay. The detailed protocol is described in the Materials and Methods section of Supplementary Data.
Cell cycle analysis
Cell cycle was analyzed by flow cytometry. The detailed protocol is described in the Materials and Methods section of Supplementary Data.
Colony forming assay
The colony forming assay protocol is described in the Materials and Methods section of Supplementary Data.
Human phosphokinase assay
The Human phosphokinase assay was performed with the Proteome ProfilerTM Array (R&D Systems, Minneapolis, MN, USA), following the manufacturer's protocol. The details of the assay protocol are described in the Materials and Methods section of Supplementary Data.
Western blot
Protein lysates were resolved in sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE) mini gels. After protein transfer, membranes were probed using appropriate antibodies, and specific binding was detected through a secondary peroxidase-conjugated antibody, using a chemiluminescence assay. The details are provided in the Materials and Methods section of Supplementary Data.
Treatment with MEK1/2 inhibitor trametinib
Trametinib was obtained from Absource Diagnostics GmbH (München, Germany). Cell counting kit-8 (Dojindo, Munich, Germany) was used to analyze cell metabolic viability. Cell viability over time was analyzed by trypan blue (Gibco; Life Technologies) exclusion assay. The details of the assays are supplied in the Materials and Methods section of Supplementary Data.
Short interfering RNA gene silencing
SPRY4 gene silencing in TPC-1 cells was achieved using short interfering RNA (siRNA). Further details of the assay are supplied in the Materials and Methods section of Supplementary Data.
Quantification of gene expression by reverse transcription-quantitative real-time PCR
Protocols of RNA extraction, complementary DNA (cDNA) synthesis and quantification of gene expression by reverse transcription-quantitative real-time PCR (RT-qPCR) are described in the Materials and Methods section of Supplementary Data.
Statistical analyses
All experiments were performed at least in three independent assays, except for human phosphokinase assay, in which two independent assays were performed. The results are expressed as the mean ± standard deviation, and p-values <0.05 were considered statistically significant. The details are described in the Materials and Methods section of Supplementary Data.
Results
Selection of a candidate susceptibility gene through WES analysis
Six members of the FNMTC family, who had thyroid cancer, were selected for WES analysis (Fig. 1).
One family branch had a stronger evidence of hereditary transmission of thyroid cancer (core family), having three siblings (IV.2, IV.6, and IV.8) and two of their offspring (V.3 and V.8) affected. Two family members with thyroid cancer (III.7 and V.67), who did not belong to the core family, and did not have ascendants or descendants with thyroid cancer, were evaluated as possible phenocopies in the bioinformatic analysis. Candidate gene variants were identified, using stringent criteria and filtering procedures (detailed information in the Materials and Methods section of Supplementary Data). The most suggestive variants were found in four candidate genes (CASP8AP2, KCTD16, SPRY4, and TBC1D14), and these were selected for further studies. Results of segregation and tumor loss of heterozygosity analyses (LOH) are detailed in the Results section of Supplementary Data and Supplementary Table S3. Characterization of the four genes (expression in thyroid, protein functional domains, and signaling pathways) and respective variants (conservation in evolution, frequency in the Portuguese population, and in silico predicted damaging effects) is also detailed in the Results section of Supplementary Data and Supplementary Table S4.
The missense variant NM_001127496.1:c.701C>T, located in exon 3 of SPRY4 (sprouty RTK signaling antagonist 4) is predicted to change the encoded amino acid from threonine to methionine (
Functional characterization of SPRY4 c.701C>T (p.Thr234Met ) variant
To investigate the biological consequences of SPRY4
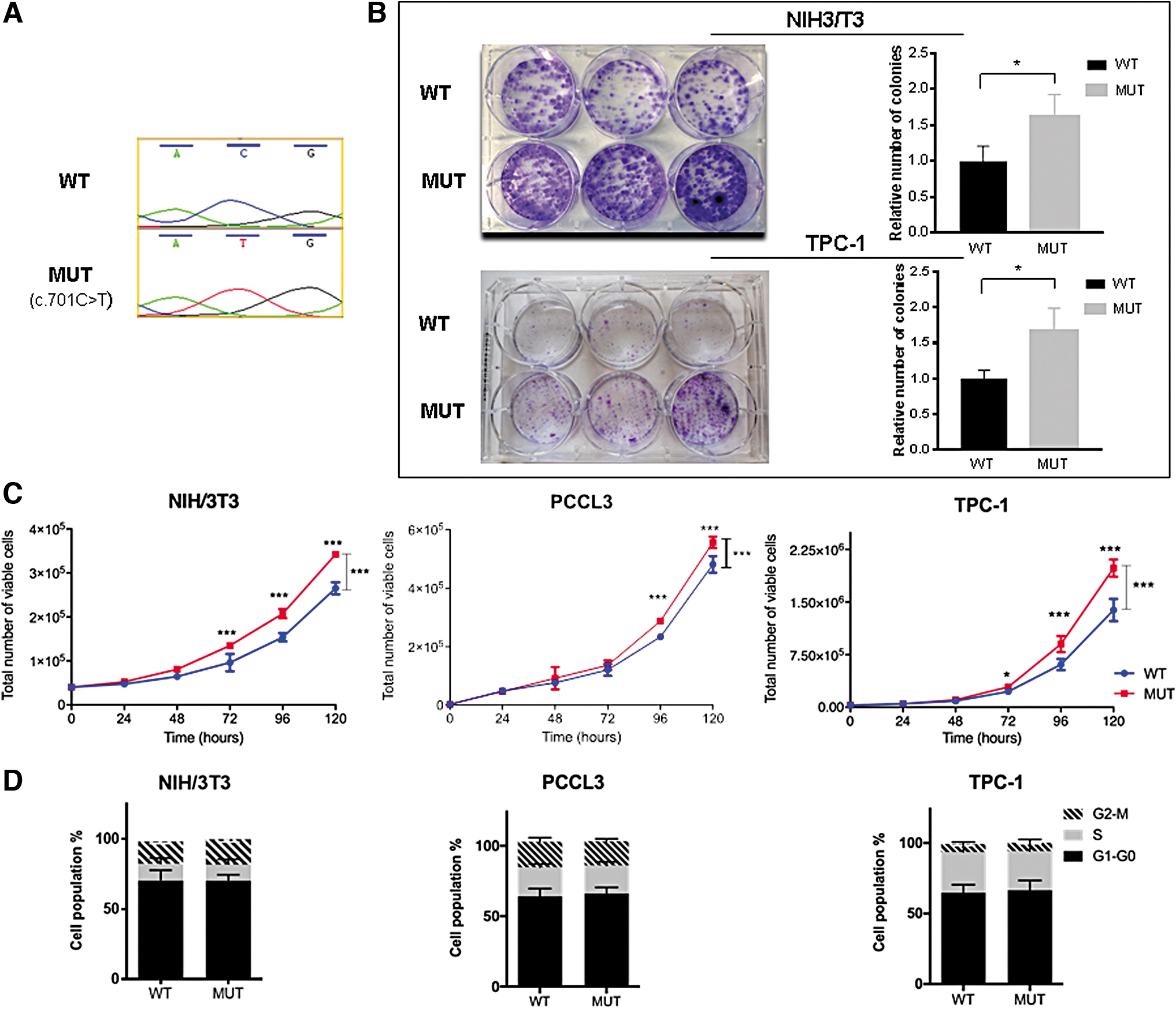
Functional studies of SPRY4
NIH/3T3 and TPC-1 cells expressing MUT SPRY4 showed a significant increase in colony formation, compared with WT cells, suggesting that the variant promotes colony-forming ability (p < 0.05; Fig. 2B). PCCL3 cells did not form colonies under these assay conditions. A significant increase in cell viability over time was observed for MUT SPRY4-expressing cells, compared with WT cells, at 96 and 120 hours, in all cell lines (p < 0.001; Fig. 2C).
The influence of MUT SPRY4 on cell cycle was analyzed by flow cytometry in NIH/3T3, PCCL3, and TPC-1 cells, but no significant differences were observed (Fig. 2D). Likewise, no significant differences were observed in cell migration ability (Supplementary Fig. S3).
Effect of SPRY4 p.Thr234Met variant in the phosphorylation of cancer-related kinases
The phosphorylation profiles of cancer-related kinases in TPC-1 cells expressing WT and MUT SPRY4 was explored, using a human phosphokinase antibody array. Overall, the fold changes, evaluated by the ratio MUT/WT, showed an increase in kinase phosphorylation levels (Fig. 3; Supplementary Fig. S4). Phosphorylation levels of activating residues in some signaling proteins and targets of ERK1/2 MAPK cascade, such as STAT3, STAT5b, MSK1/2, and p53 (31), were significantly increased (≥1.5-fold change) for MUT SPRY4-expressing cells compared with WT. In addition, this analysis showed a significant increase in PYK2 and PLC-γ1, also related with activation of MAPK signaling (32). Accordingly, Western blot analysis showed an increase in ERK phosphorylation levels (p-ERKT202/Y204) in TPC-1 MUT SPRY4 protein lysates, compared with WT, while there were no corresponding changes in total ERK abundance, and SPRY4 levels were similar, suggesting that an increase in ERK activity could be associated with SPRY4 mutated status (Fig. 4).
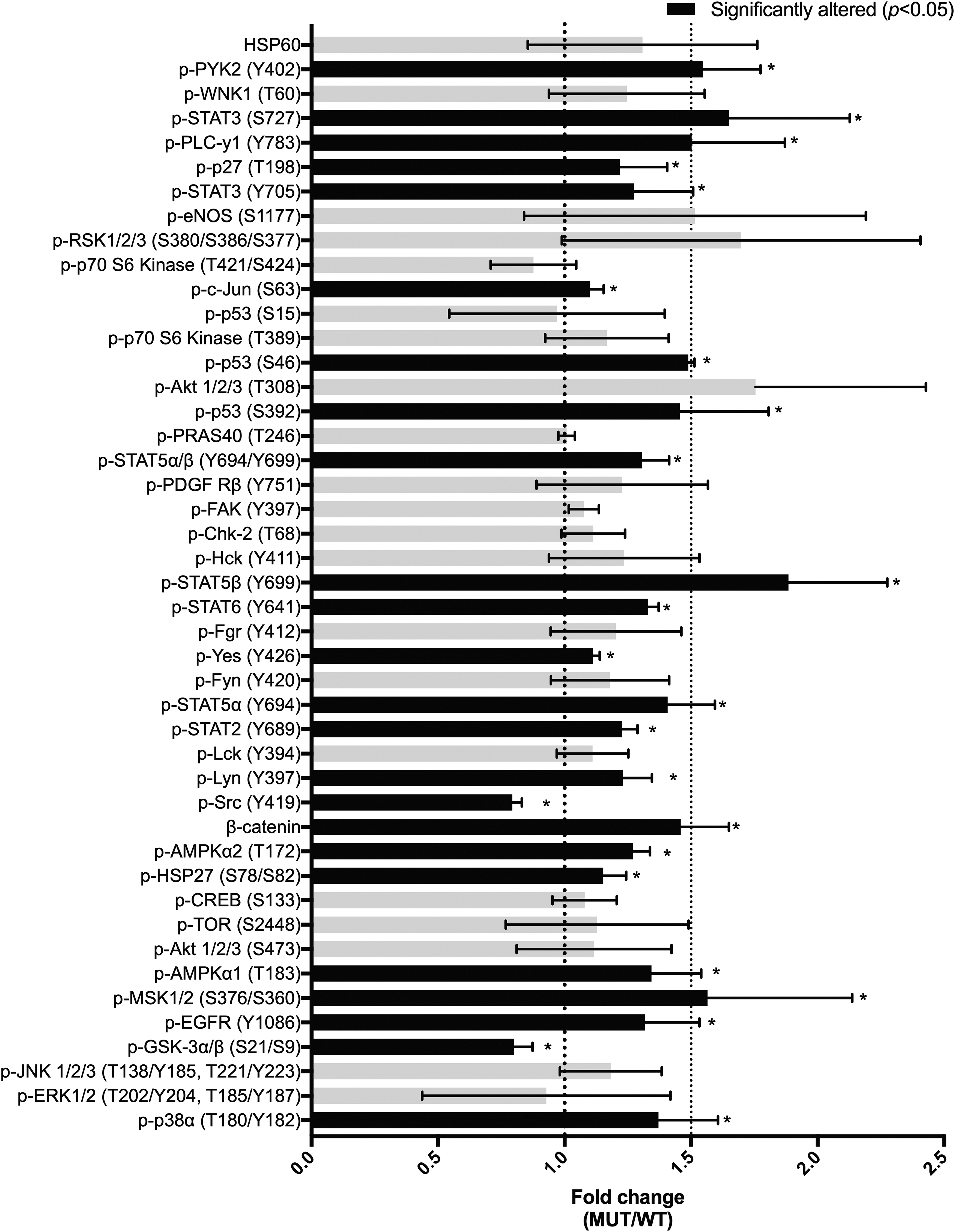
SPRY4
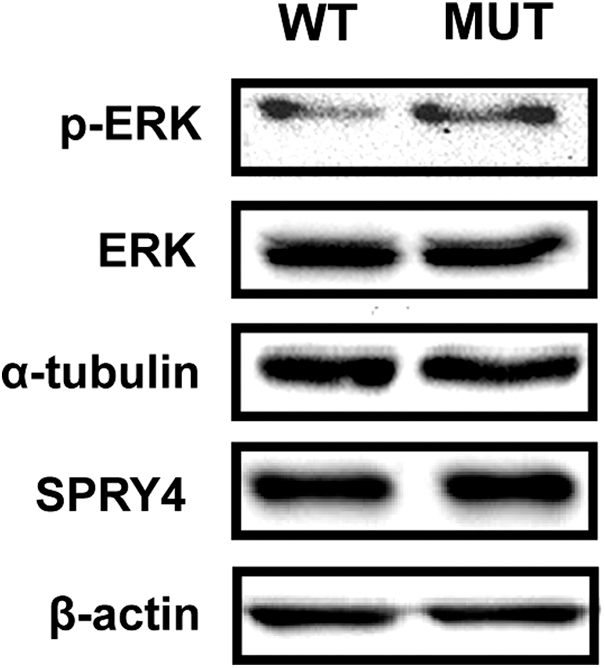
Effect of WT and MUT SPRY4 in ERK kinase activity in TPC-1 cells. Western blot analysis of phospho-ERK (p-ERK) and total ERK, in TPC-1 cells expressing WT and MUT (
Treatment of TPC-1 cells expressing WT and MUT SPRY4 with MEK1/2 inhibitor trametinib
Trametinib, a highly specific and potent MEK1/2 inhibitor, at nanomolar doses (0.5–100 nM) decreased metabolic cell viability in a dose-dependent manner, and this effect was more pronounced in TPC-1 cells expressing MUT SPRY4 compared with WT [trending with 0.5–5.0 nM trametinib, and statistically significant with 10 (p < 0.05), 50 (p < 0.01), and 100 nM (p < 0.01)] (Supplementary Fig. S5). The use of 5.0 nM trametinib induced a low number of viable TPC-1 WT and MUT cells over time (data not shown). Treatment with 0.5 nM trametinib induced a significant decrease in cell viability over time in both TPC-1 WT and MUT cells, compared with the corresponding nontreated cells (p < 0.001), but this decrease was significantly more pronounced in MUT cells compared with WT (p = 0.003) (Fig. 5).
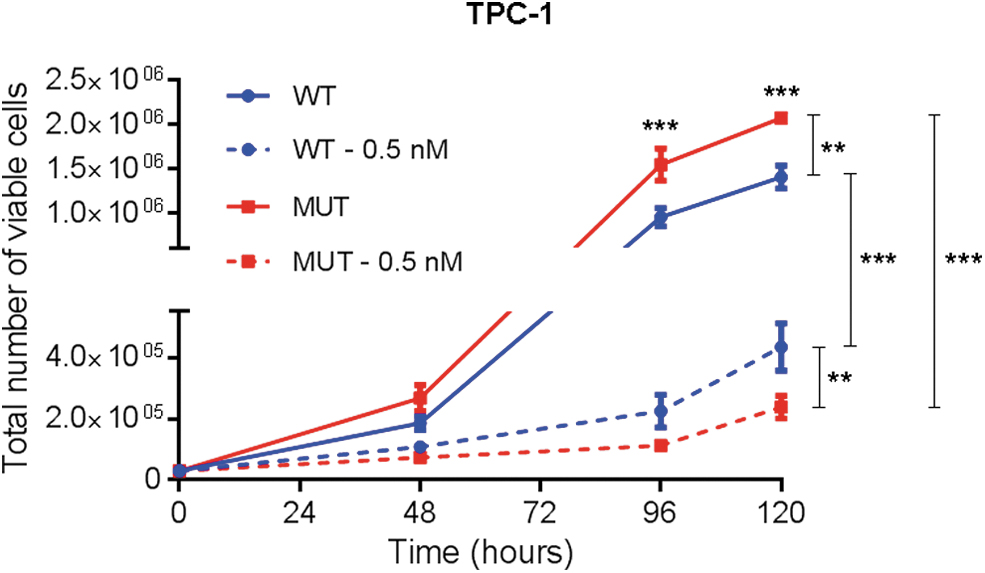
Treatment of TPC-1 cells expressing wild-type and MUT SPRY4 with MEK1/2 inhibitor trametinib. Cell viability over time of TPC-1 cells expressing WT and MUT (
Effect of SPRY4 silencing in TPC-1 cell viability
The effect of silencing SPRY4 in TPC-1 cells was analyzed using siRNA. Acceptable levels of silencing were obtained (Supplementary Fig. S6). A significant decrease in the total number of viable cells was observed in SPRY4 silenced cells (siSPRY4), compared with control (siCTR) or original TPC-1 cells (p < 0.001); siCTR and original TPC-1 viabilities over time were similar (Supplementary Fig. S7).
Screening of SPRY4 gene mutations in FNMTC patients
The entire coding sequence and splicing boundaries of SPRY4 were analyzed in probands from 96 Portuguese families with FNMTC. However, no potentially pathogenic variants were detected.
Screening of somatic BRAF and RAS mutations in familial tumors
To explore the role of somatic mutations in HRAS, NRAS, KRAS, and BRAF proto-oncogenes, in tumor progression in this family, the mutational hotspots of these genes were sequenced in the thyroid cancers of 6/7 affected members, as tumor sample from patient III.7 was not available [results partially reported earlier in (26)]. We found that 5/6 familial thyroid cancers had a somatic HRAS mutation, whereas only one patient had a BRAF mutation (Table 1).
Mutations in HRAS and BRAF Genes in Thyroid Carcinomas from Patients of the Family with Familial Nonmedullary Thyroid Cancer
RefSeqs: NM_005343.4 (HRAS) and NM_004333.6 (BRAF).
Transversion.
Transition.
c, classic; fv, follicular variant; PTC, papillary thyroid carcinoma.
Discussion
Identification of cancer-predisposing mutations is a critical step in cancer risk assessment and can help in cancer screening and early detection strategies.
In this study, we selected one of the most informative families with FNMTC from our cohort and performed WES analysis in the patients with thyroid cancer.
Four variants in the candidate genes KCTD16, CASP8AP2, TBC1D14, and SPRY4 were initially selected. However, CASP8AP2, KCTD16, and TBC1D14 were not prioritized for further studies. Still, it cannot be ruled out the possibility that a combination of these gene variants, which would most likely have medium/low penetrance, might contribute to thyroid cancer predisposition.
SPRY4 gene was the main candidate for FNMTC predisposition in this family. The SPRY4 gene variant, which consisted of a cytosine to thymine transition (c.701C>T,
Sprouty proteins have conserved motifs, which include a C-terminal cysteine-rich domain (CRD) known as sprouty domain. SPRY4 c.701C>T is predicted to change a threonine located in the sprouty domain. Somatic SPRY4 mutations have been described in several cancers, such as uterine (2.3%), stomach (1.4%), and colorectal (1.0%), and, particularly, the mutation found in this study was described in a large intestine cancer [The Cancer Genome Atlas—PanCancer Atlas (33)]. A comprehensive analysis of 496 PTCs showed absence of SPRY4 mutations in these tumors (34).
Sprouty proteins are modulators of ERK signaling acting on different points of the pathway, which also influence other pathways to control proliferation, differentiation, motility, and survival (28,35). Mammalian Sprouty4 inhibits vascular endothelial growth factor (VEGF)-induced activation of ERK, in a Ras-independent manner, by binding through the CRD domain to Raf1 (28,30). Aberrant activation of ERK and deregulation of Sprouty occurs in distinct cancers, in a cancer cell-specific and context-dependent manner, promoting or inhibiting tumor progression (28). The suppressive effect of SPRY4 in cell migration and cancer progression and/or its underexpression, has been described in melanoma, lung, prostate, breast, colorectal, and medullary thyroid cancers (36 –41). In contrast, SPRY4 knockdown inhibited testicular germ cell tumor growth (42). Thus, SPRY4 has a critical role in cancer, mainly as modulator of the MAPK/ERK pathway, a major pathway involved in thyroid carcinogenesis (29).
In this study, we observed that MUT SPRY4 (c.701C>T,
In this study, siRNA-mediated SPRY4 gene silencing induced a significant decrease of TPC-1 viable cells, suggesting that this gene may have an oncogenic activity in follicular cell derived thyroid cancer, which has not been reported yet. This is consistent with absence of LOH in six thyroid tumors from affected members of this family. Similarly, in cells from human testicular germ cell tumor and from ovarian cancer, SPRY4 siRNA-mediated knockdown resulted in decreased cell migration and invasion (42,48).
In a series of 54 FNMTC tumors (93% were PTCs), we previously identified somatic mutations in BRAF (41%), NRAS (2%), and HRAS (2%) (26,27). Characterization of RAS and BRAF mutations in tumors from affected members from the present family revealed that 5/6 had HRAS mutations. These findings were surprising since HRAS mutations prevalence is low in both familial (frequency referred earlier) and sporadic PTCs (3.2%) (33). In this family, because the SPRY4 mutation was found to be germline, we postulate that it represents an initial genetic event. Subsequent somatic alterations, such as HRAS mutations, could contribute to neoplastic transformation of thyroid follicular cells. Studies with HRAS transformed human fibroblasts revealed that SPRY2 (member from the sprouty proteins family) and HRAS interact, and that SPRY2 plays a critical role in the ability of these cells to form tumors in athymic mice (49). Hence, additional studies are needed to unveil the molecular mechanisms of HRAS somatic mutation, and its possible interplay with SPRY4 in tumor development.
In summary, WES analysis and functional assays identified a novel candidate susceptibility gene, SPRY4, in the FNMTC family studied. Further studies of SPRY4 may provide insights into its function and whether it is a susceptibility gene for other cases of FNMTC.
Footnotes
Acknowledgments
Authors are thankful to Ana T. Pinto for support in phosphokinase arrays data analysis, and to Rafael Luís for sequence data submission to Sequence Read Archive. The authors are grateful to the patients and their families for their co-operation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Liga Portuguesa Contra o Cancro—Núcleo Regional do Sul (LPCC-NRS), Televisão Independente, Instituto Português de Oncologia de Lisboa Francisco Gentil, iNOVA4Health—UIDB/04462/2020, a program financially supported by Fundação para a Ciência e Tecnologia/Ministério da Educação e Ciência, and Associação de Endocrinologia Oncológica. The authors are thankful to LPCC-NRS that granted the researcher Marta Pojo. Inês J. Marques was a recipient of a PhD scholarship from the PhD Programme ProRegeM (Mechanisms of Disease and Regenerative Medicine) approved by Fundação para a Ciência e Tecnologia (FCT) - PD/BD/108086/2015, and Carolina Pires was granted with a PhD scholarship by FCT - 2020.07120.BD.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4