
Abstract
Objective:
Placental iodide transport is necessary for maintaining an adequate iodide supply to the developing fetus. We hypothesized that compounds from the placental barrier can compensate for decreases in maternal iodine intake and normalize fetal iodine levels.
Methods:
Pregnant rats administered different amounts of iodine (1.24, 2.5, 5, or 10 μg/day) were evaluated on gestational day (gd) 16 and 20. The iodine levels in maternal blood, amniotic fluid (AF), and placental tissue were estimated using As-Ce catalytic spectrophotometry. The protein and/or messenger RNA (mRNA) levels of sodium iodide symporter (NIS), pendrin, alpha-smooth muscle actin (α-SMA), and CD31 in the placental labyrinth, trophoblast cells isolated using laser capture microdissection (LCM), and/or fetomaternal thyroid were detected using immunoblotting, real-time polymerase chain reaction, and/or immunohistochemistry.
Results:
When iodine intake was reduced, iodine levels in maternal blood gradually decreased; however, placental iodine levels were not significantly different between groups on gd16 and gd20. Minimal changes were observed in AF iodine levels on gd16, and a mild decreasing trend was observed (iodine dose, 10 to 1.24 μg/day) on gd20. NIS protein, which was linearly distributed along the basolateral membrane of maternal–fetal thyroid follicles, gradually increased with decreasing iodine levels. Regarding iodine deficiency in the placental labyrinth on gd16 and gd20, pendrin and glycosylated NIS proteins were significantly upregulated in a dose-dependent manner. However, the mRNA levels were unchanged. Furthermore, the conversion of NIS protein from the nonglycosylated to the glycosylated form increased. In trophoblast cells isolated using LCM, PDS mRNA levels increased in the 1.24-μg/day group on gd16 but not NIS mRNA levels. There was a smaller α-SMA+ area in the labyrinth zone on gd16 and gd20; however, the proportional CD31+ area increased on gd16 and reduced on gd20 with decreased iodine levels.
Conclusions:
All mechanisms upregulating the expression of iodine transporters and changes in villous stroma and microvessel area in the placental labyrinth can promote iodide transfer from mother to fetus in iodine deficiency, especially before the onset of fetal thyroid function. Compensatory NIS protein regulation in the placenta against decreased iodine intake mainly occurs during translation and glycosylation modification after translation. Pendrin may be more important than NIS in the mediation of placental iodide transport.
Introduction
Iodine is an essential micronutrient for hormones produced by the thyroid gland and is especially crucial during pregnancy for fetal growth and development. Iodine deficiency during gestation leads to iodine deficiency disorders that result in severe and irreversible cognitive and physical impairments and in extreme cases, cretinism (1). The fetal thyroid gland can accumulate iodide and synthesize thyroid hormones by week 11 (2), but secretion of function thyroid hormone does not being until mid-second trimester, around 16–17 weeks of gestation (3). The available iodide for the fetal thyroid gland is mainly from the mother, and the capacity of the placenta to transport iodide from maternal blood to the fetus is very important (4).
Iodide transporters, such as the sodium iodide symporter (NIS) encoded by the SLC5A5 gene and pendrin encoded by the SLC26A4/PDS gene, can facilitate iodide uptake from blood into the thyrocytes and iodide release to follicles, which is important in thyroid hormone biosynthesis (5,6). When iodine intake is inadequate, transporter expression is upregulated to accumulate more iodide. In previous reports, both NIS and PDS messenger RNA (mRNA) were shown to be expressed in the human placenta, although at low levels compared with in the thyroid tissue (7,8), and during the accumulation of iodide in placental cell lines (9). The degree of iodine deficiency varied in parallel between mothers who smoke and their fetuses with no signs of worsening in the fetus, indicating that the autoregulation of placental iodide transport is similar to that in the thyroid (10).
Immunohistochemical analysis showed that pendrin is expressed at the brush border membrane of syncytiotrophoblast cells and NIS protein is mainly located in the villous cytotrophoblast layer (8). Therefore, placental iodine uptake from maternal blood is mediated by pendrin, which is different from thyroidal iodine uptake by NIS from blood. Reportedly, the upregulation of NIS mRNA in the fetal side of the placenta was also demonstrated in rats on a low-iodine diet (11), indicating the existence of an adaptive mechanism when iodine intake is inadequate to ensure that the fetus receives sufficient iodine during development. However, specific details, such as protein changes, remain unclear.
In addition to the trophoblast layer, iodide from maternal circulation has to traverse the villous stroma and capillary endothelium before entering fetal circulation. However, whether villous trophoblast cells, the villous stroma, and fetal vascular elements can efficiently regulate placental iodide transfer when iodine intake is changed is unclear. Therefore, after estimating the iodine status in a series of samples from rat dams and pups suffering from severe-to-mild iodine deficiency, we focused on the compensatory effects of NIS and pendrin expression in the placental labyrinth as well as the proportional area of villous stroma and vascular elements labeled using alpha-smooth muscle actin (α-SMA) or platelet endothelial cell adhesion molecule-1 (CD31) during mother-to-fetus iodide transfer.
Materials and Methods
Animals
Sixty-four specific pathogen-free (SPF) female Wistar rats (age, 4–6 weeks; body weight, 70–100 g) were purchased from Vital River Laboratory Animal Technology (Beijing, China). The animals were raised for one week with normal food (300–400 μg/kg iodine content) and tap water (8 μg/L iodine content) in a single SPF condition room (temperature, 26°C ± 3°C; humidity, 40–60%; 12-hour light/12-hour dark cycle).
All animal and experimental procedures were approved by the Animal Research Committee of Tianjin Medical University. The experiments and procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Study design
After adaptive feeding, all female rats were randomly allocated to four groups of eight rats each and fed for three months with iodine-deficient food and tap water with different doses of potassium iodide as in our previous report (12). Next, iodine intake in the four groups was as follows: 1.24 μg/day (severe iodine deficiency intake), 2.5 μg/day (moderate iodine deficiency intake), 5 μg/day (mild iodine deficiency intake), and 10 μg/day (adequate iodine intake, control group). The day when the vaginal plug was identified was designated gestational day (gd) 0 after mating with normal male Wistar rats (body weight, 250–300 g; purchased from Vital River Laboratory Animal Technology). The rats were anesthetized, euthanized using femoral artery bleeding on gd16 and gd20, and the uteri were dissected to harvest embryos and placentas from at least five dams.
Determination of iodine contents
Urine was collected in a metabolic cage before pregnant rats were sacrificed. Femoral arterial blood with heparin anticoagulant, amniotic fluid (AF), and placental tissues were collected on days 16 and 20 of pregnancy. All samples were kept at −80°C until use.
The AF samples were kept at room temperature for 30 minutes and centrifuged at 3000 rpm for 15 minutes (Sigma 3k15). The supernatant was then transferred into a new tube. The AF supernatant for each dam was pooled over six fetuses per litter for iodine determination.
Placental tissues were rinsed several times in cold deionized water until red blood cells were not observed in the wash buffer under a microscope at ×100 magnification and then dried with filter paper. The tissues were weighed and homogenized with 1.5 mL of cold water per sample. Results are expressed as μg iodine/g tissue wet weight.
Iodine levels in urine, blood, AF, and placental tissues were detected using As-Ce catalytic spectrophotometry according to the Health Industry Standard of the People's Republic of China (WS/T107-2006 or WS 302-2008). All samples were run in duplicate.
Histology and immunohistochemistry
For histomorphology, fetomaternal thyroid and placental tissues were fixed in 4% (w/v) paraformaldehyde and embedded in paraffin. Sections (4-μm thick) were cut, mounted on glass slides, and stained with hematoxylin and eosin (H&E).
For immunohistochemical analysis, sections were stained using a Polink-2 Plus Polymer HRP Detection System Kit (Zhongshan Golden Bridge Biotechnology, Beijing, China) according to the manufacturer's instructions. Thyroid tissues were incubated with rabbit-anti-rat NIS polyclonal antibody (1:50; Alpha Diagnostic Intl, San Antonio, TX). Antibodies against α-SMA (1:100; Abcam, Cambridge, MA) and CD31 (1:50; Boster Biological Technology co.ltd, Wuhan, China) were used to label placental villous stroma and fetal endothelial cells, respectively. Nuclei were counterstained with or without hematoxylin. Control sections were treated similarly, but the primary antibodies were omitted. Images were captured using an Olympus BX53 light microscope (Olympus Corporation, Tokyo, Japan) equipped with a DP72 microscope digital camera (Olympus Corporation). Morphometry was performed using the Image Pro Plus 7.0 software (Media Cybernetics, Rockville, MD), and the percentage of villous stroma or vascular elements (α-SMA- or CD31-positive) within the labyrinth was analyzed in five different fields per placenta from five dams per group at ×400 magnification.
Western blot analysis
As previously described, protein was extracted from the placental labyrinth using RIPA buffer (Cell Signaling Technology, Danvers, MA) supplemented with protease inhibitors (Sigma–Aldrich, St. Louis, MO) and quantified using the BCA protein assay (Pierce Biotechnology, Rockford, IL). Lysates were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA). The membranes were incubated with rabbit anti-NIS and anti-pendrin polyclonal antibodies at 1:200 dilution and anti-β-actin at 1:2000 dilution (Santa Cruz Biotechnology, Santa Cruz, CA). Densitometry was performed using the ImageJ 1.8.0 software.
Laser capture microdissection and reverse transcription–quantitative polymerase chain reaction
Trophoblast cells in the stem villi of the placental labyrinth in every group were isolated using laser capture microdissection (LCM). Briefly, 8-μm thick tissue cryosections of central placenta were immobilized on RNase-free slides, stained for 1 minute with 0.1% diethylpyrocarbonate-treated hematoxylin, and air-dried at room temperature for 10 minutes. Trophoblast cells in typical stem villi of the placental labyrinth were identified; proper parameters of energy, focal length, and number of points were established; and capture was performed by Laser Microdissection and Pressure Catapulting using a MicroBeam microscope (P.A.L.M. Microlaser Technologies). Isolated cells were placed directly into RNA extraction buffer and kept at −80°C until use.
As previously described (12), the total RNA in the trophoblast cells and placental labyrinth was extracted and reverse-transcribed to complementary DNA as well as quantitative polymerase chain reaction (qPCR) was performed. The following primers were used to generate the polymerase chain reaction (PCR) fragments: NIS, 5′-ACCAACGCTTCCAACGG-3′ and 5′-GAGATAGGAGATGGCGTAAA-3′; PDS, 5′-CATCAAGACACATCTCCGTTGGCCCT-3′ and 5′-GGTACTTCCGTTACCACTGGGC-3′; and UBC (as a housekeeping gene for normalization), 5′-TCGTACCTTTCTCACCACAGTATCTAG-3′ and 5′-GAAAACTAAGACACCTCCCCATCA-3′. All primers produced a PCR product that resolved as a single band with no primer dimer on agarose gels. All samples were run in triplicate and repeated on another day. The annealing temperature was 60°C for all primer pairs.
The initial template quantities of the genes NIS, PDS, and UBC were calculated using the “4- to 6-point method” as described by Ramakers et al. (13). The ratios of initial template quantities (NIS:UBC and PDS:UBC) were used as the relative expression levels for NIS and PDS mRNA, respectively.
Measurement of rat chorionic gonadotropin
Serum samples of pregnant rats were analyzed using enzyme-linked immunosorbent assay kit (USCN Life Science, Wuhan, China) according to the instructions of the kit. The detected concentration range is 1.56–100 mIU/mL. Briefly, 100 μL of Standards, Blank, or serum samples per well were added to a 96-well plate coated by purified anti-rat chorionic gonadotropin (rCG) antibody. Then, the anti-rCG antibody, detection reagent, substrate solution, and stop solution was added to each well in turn, and the absorbance was measured at 450 nm. All assays were performed in duplicate. rCG concentrations were calculated using optical density values against standard curves calibrated with known concentration values.
Statistical analyses
Statistical analyses were performed using the SPSS 24.0 statistical package (IBM Corporation, Armonk, NY). Iodine levels are expressed as medians and compared using the nonparametric Kruskal–Wallis test. The other quantitative data are expressed as the mean ± standard deviation. Differences between groups were evaluated using one-way analysis of variance (ANOVA) and the F-test. If the ANOVA revealed significant differences among groups, individual groups were compared with the control group using the least significant difference test or Tamhane's T2-test. A p-value <0.05 was considered to denote statistical significance.
Results
Iodine content in different samples
The median urinary iodine levels in dams with iodine intakes of 1.24, 2.5, 5, and 10 μg/day were 0, 26.9, 115.9, and 353.7 μg/L, respectively. The iodine levels in maternal blood, fetal AF, and placental tissues are shown in Figure 1A.
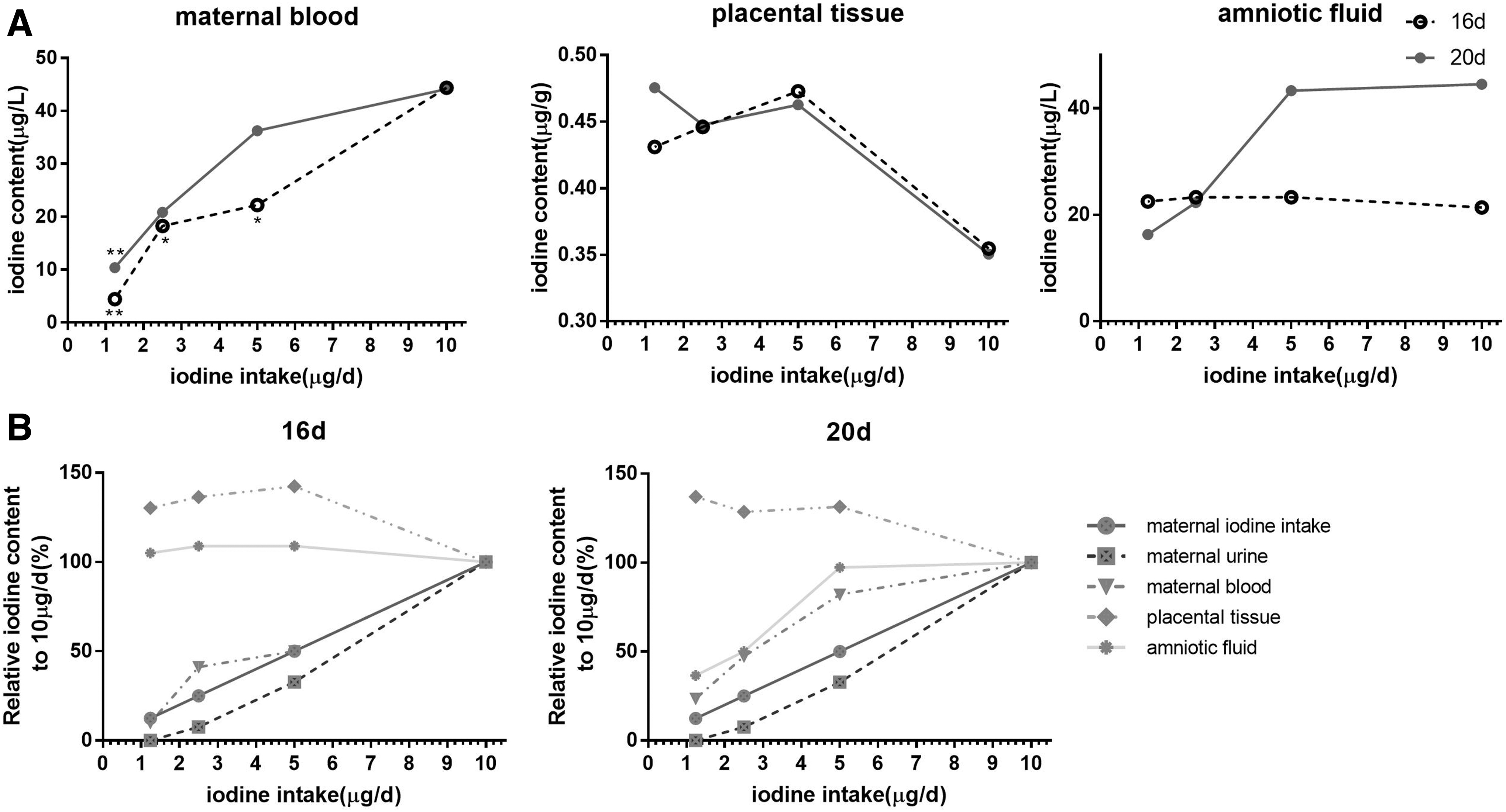
Iodine contents in blood of pregnant rats, placental tissue, and amniotic fluid at two gestational days (
The iodine levels in the urine and blood of pregnant rats in each group increased as the amount of iodine administered to the pregnant rats increased. Furthermore, as the amount of iodine administered to the pregnant rats increased, the iodine content in AF did not change significantly on gd16 but increased on gd20. Compared with the 10-μg/day control group, the iodine level in placental tissue did not differ between gd16 and gd20. However, the amplitudes of the various iodine levels differed among the groups as follows: urine iodine in the dams > dietary iodine intake of the dams > blood iodine in the dams > iodine in AF > iodine in placental tissue (Fig. 1B).
Location and expression of NIS protein in maternal–fetal thyroids
Representative positive photomicrographs of the NIS protein in maternal–fetal thyroids on gd20 are shown in Figure 2A. Whether on gd16 or gd20, maternal thyroid NIS protein in the 10-μg/day iodine intake group was found mostly in the cytoplasm and rarely at the cell membrane between follicle epithelial cells. However, when the pregnant rats suffered from iodine deficiency, most of the NIS protein was linearly distributed along the basolateral membrane of thyroid follicles, indicating that these cells could transport iodide. Furthermore, a positive correlation was observed between thyroid NIS expression and degree of iodine deficiency.
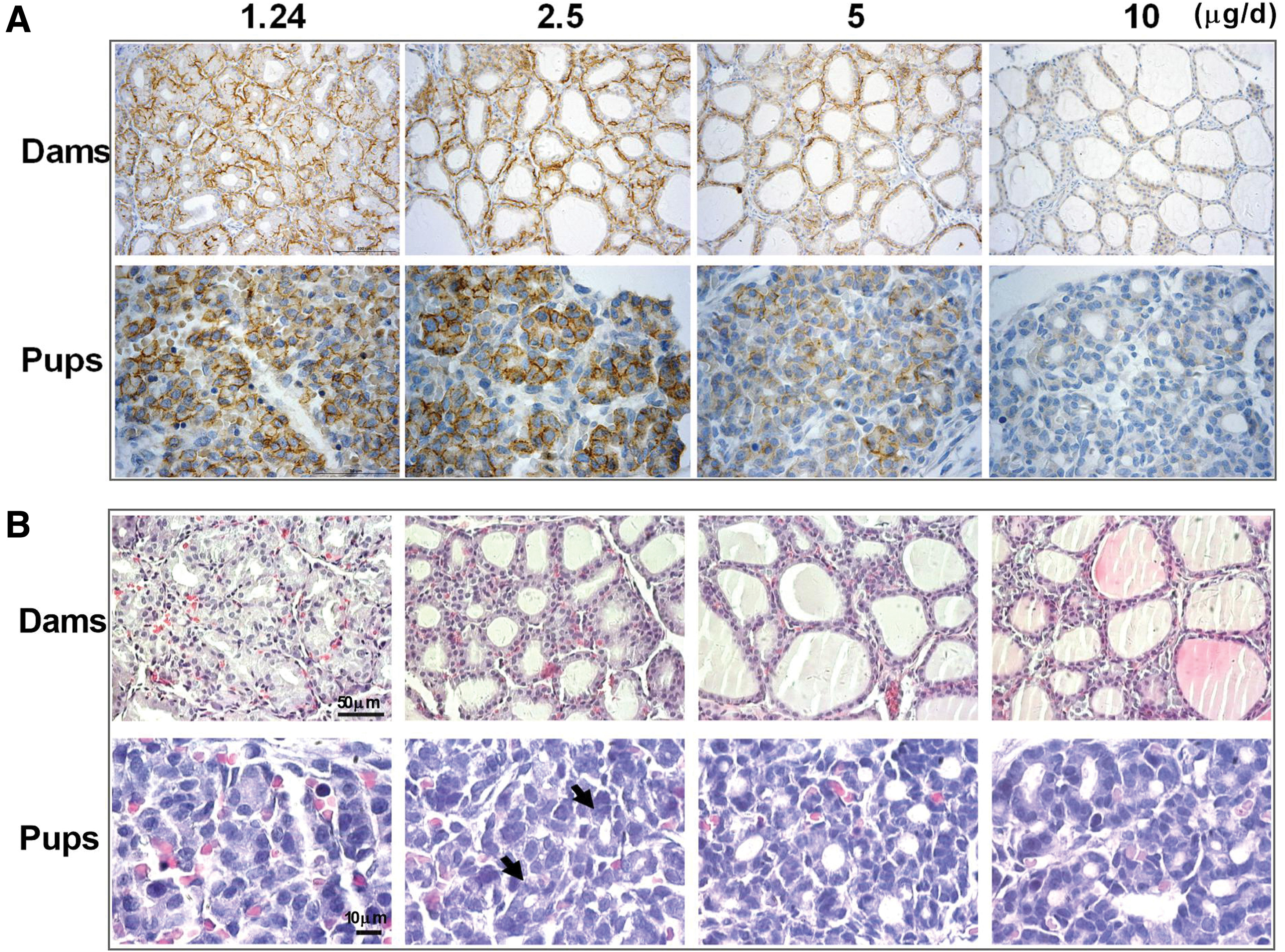
Representative photomicrographs of NIS protein (
In fetal thyroids on gd20, most of the NIS protein was located at the cell membrane in the 1.24-, 2.5-, and 5-μg/day iodine intake groups, and increased immunostaining was observed with increased maternal iodine intake. Normal fetal thyroid follicles did not appear in the severe iodine deficiency group (1.24 μg/day), and NIS immunoreactivity was observed along the whole cell membrane of thyrocytes.
Maternal–fetal thyroid histology
On gd16 and gd20, the thyroids of dams were increasingly enlarged when iodine intake was reduced from 10 to 1.24 μg/day. Furthermore, the absolute and relative thyroid weights in the severe iodine deficiency group (1.24 μg/day) were significantly higher than those in the adequate iodine intake control group (10 μg/day) on gd16 and gd20 (all p < 0.01; Supplementary Fig. S1).
H&E staining of maternal thyroids revealed the presence of typical goiter with smaller follicles, fewer follicular colloids, and more vessels that increased with decreasing iodine intake on gd16 and gd20 (Fig. 2B in dams). Smaller thyroid follicles and hyperemia were observed in pups in the 2.5- and 5-μg/day iodine groups, respectively, compared with pups in the 10-μg/day control group; similar findings were observed in dams. Notably, normal thyroid follicles were not observed in the fetal thyroids of pups in the 1.24-μg/day iodine group, indicating that thyroid development during the embryonic period was inhibited by severe iodine deficiency (Fig. 2B in pups).
NIS and pendrin protein expression in placental labyrinth tissues
NIS is a membrane glycoprotein and its function is regulated by post-translational glycosylation as reported in cancer studies (14). Although NIS reportedly has a hyperglycosylated form with a molecular weight of ∼70–90 kDa that represents the mature and main iodide-pumping form (15), based on immunoblot analysis of the fetal rat placenta on gd16 and gd20 in the present study, NIS was in a nonglycosylated form with a molecular weight of ∼50 kDa and a glycosylated form with a molecular weight of 60 kDa (Fig. 3A). The 60 kDa bands on gd16 and gd20 increased with reduced iodine intake, and statistically significant differences were observed in rats with lower iodine intakes of 1.24 and 2.5 μg/day (Fig. 3B, C; p < 0.05 and p < 0.01, respectively) compared with the control. However, the 50 kDa bands were different on gd16 and gd20, indicating downregulation in the lower (1.24 and 2.5 μg/day) iodine intake groups (all p < 0.05) on gd16; however, on gd20, upregulation was observed in the 2.5- and 5-μg/day groups (Fig. 3B, C; all, p < 0.05). However, the ratio of 60-kDa to 50-kDa NIS protein on gd16 and gd20 increased in the two lower iodine intake groups (1.24 and 2.5 μg/day), with a significant difference observed on gd16 (Fig. 3D; all, p < 0.05). This indicates that there was an increase in the conversion from the nonglycosylated to the glycosylated form to enhance iodide-pumping ability.
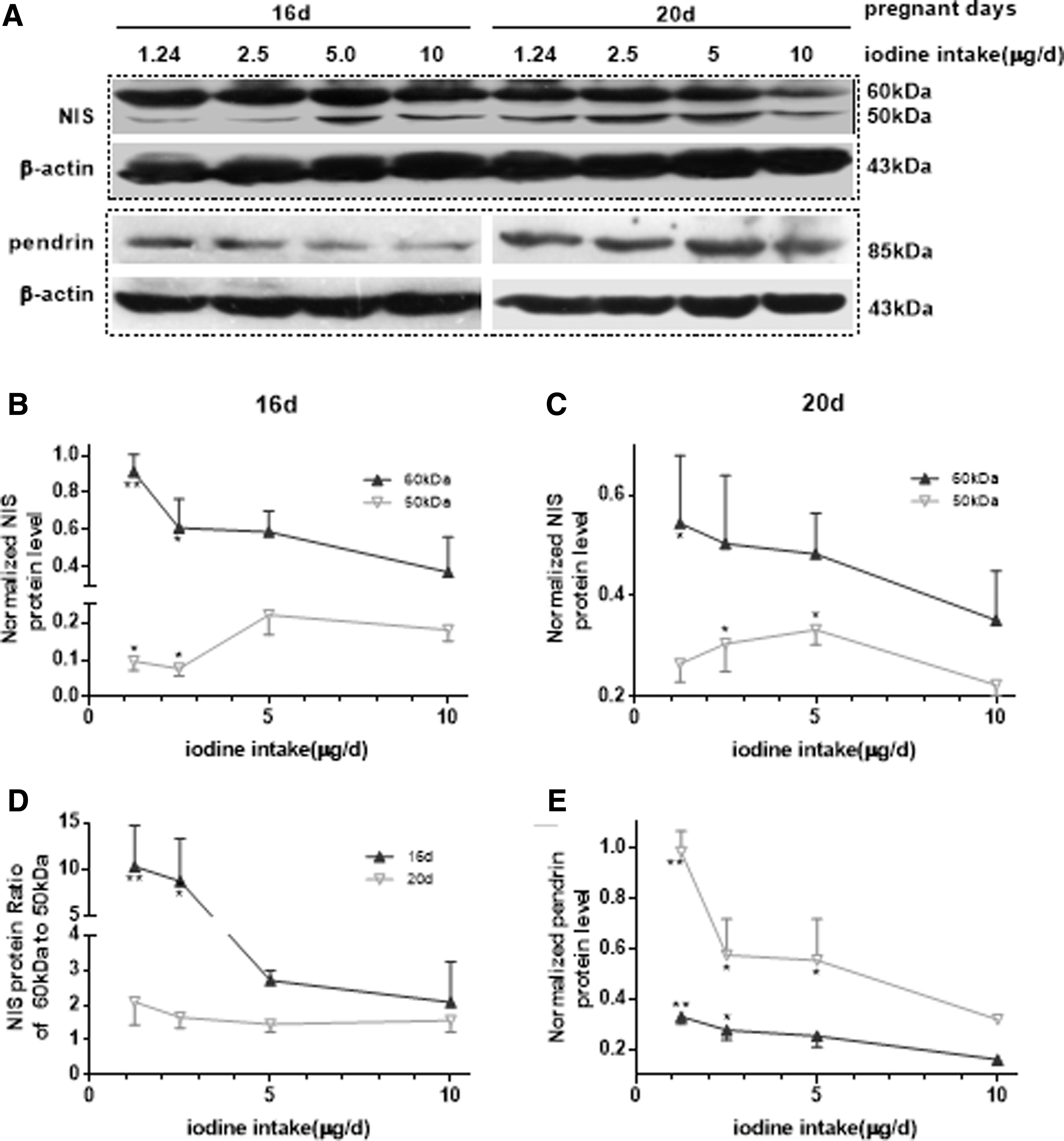
Expression of NIS and pendrin protein levels in the rat placenta labyrinths. (
Pendrin protein levels increased on gd16 and gd20 with decreased iodine intake and were significantly different between the 1.24- and 2.5-μg/day groups and the control on gd16 and between the 1.24-, 2.5-, and 5-μg/day groups and the control on gd20 (Fig. 3E; p < 0.05 and p < 0.01, respectively).
NIS and PDS mRNA levels in placental labyrinth tissues and trophoblast cells isolated using LCM
To investigate whether the transcriptional levels of NIS and PDS genes were activated during inadequate iodine intake, qPCR was performed using labyrinth tissue samples. NIS mRNA under an adequate iodine intake decreased substantially, ∼80-fold (0.036 ± 0.011 on gd16 vs. 0.00044 ± 0.00003 on gd20), and PDS mRNA levels remained unchanged (0.0021 ± 0.0001 on gd16 vs. 0.0013 ± 0.0002 on gd20) with the progression of pregnancy from gd16 to gd20. However, significant differences were not found among the four groups on gd16 and gd20 (Fig. 4A).

mRNA expressions of NIS and PDS in the rat placental labyrinth (
Next, to confirm whether activated transcriptional levels of NIS and PDS genes only occurred in trophoblast cells where they are mainly expressed, the distinguishable trophoblast cells from the villi stem were isolated using LCM. In general, with the progression of pregnancy, NIS mRNA levels in trophoblast cells under a normal iodine intake decreased approximately fivefold (1.32 ± 0.30 on gd16 vs. 0.25 ± 0.10 on gd20); however, PDS mRNA levels increased approximately ninefold (0.0056 ± 0.0028 on gd16 vs. 0.051 ± 0.026 on gd20). Similar to the placental labyrinth tissues, trophoblast NIS mRNA was unchanged under a deficient iodine intake on gd16 and gd20. However, PDS mRNA in trophoblast cells was gradually upregulated with decreased iodine intake, especially on gd16, and mRNA levels were significantly higher in the 1.24-μg/day group compared with the 10-μg/day group (Fig. 4B; p < 0.05).
Determination of villous stroma and vascularity in the labyrinth zone
Immunohistochemistry and image analysis showed an obvious reduction in α-SMA+ proportional area in the 1.24-μg/day iodine group on gd16 and gd20 (both, p < 0.05; Fig. 5). Additionally, a slight reduction in α-SMA+ proportional area was observed in the 2.5- and 5-μg/day groups on gd20 (both, p > 0.05). However, increased CD31 immunoreactivity was observed with a decreased iodine intake from 10 to 1.24 μg/day (both in the 2.5- and 1.24-μg/day groups; p < 0.01) on gd16. However, on gd20, the CD31+ proportional area was significantly reduced in the 1.24-μg/day group (p < 0.01).
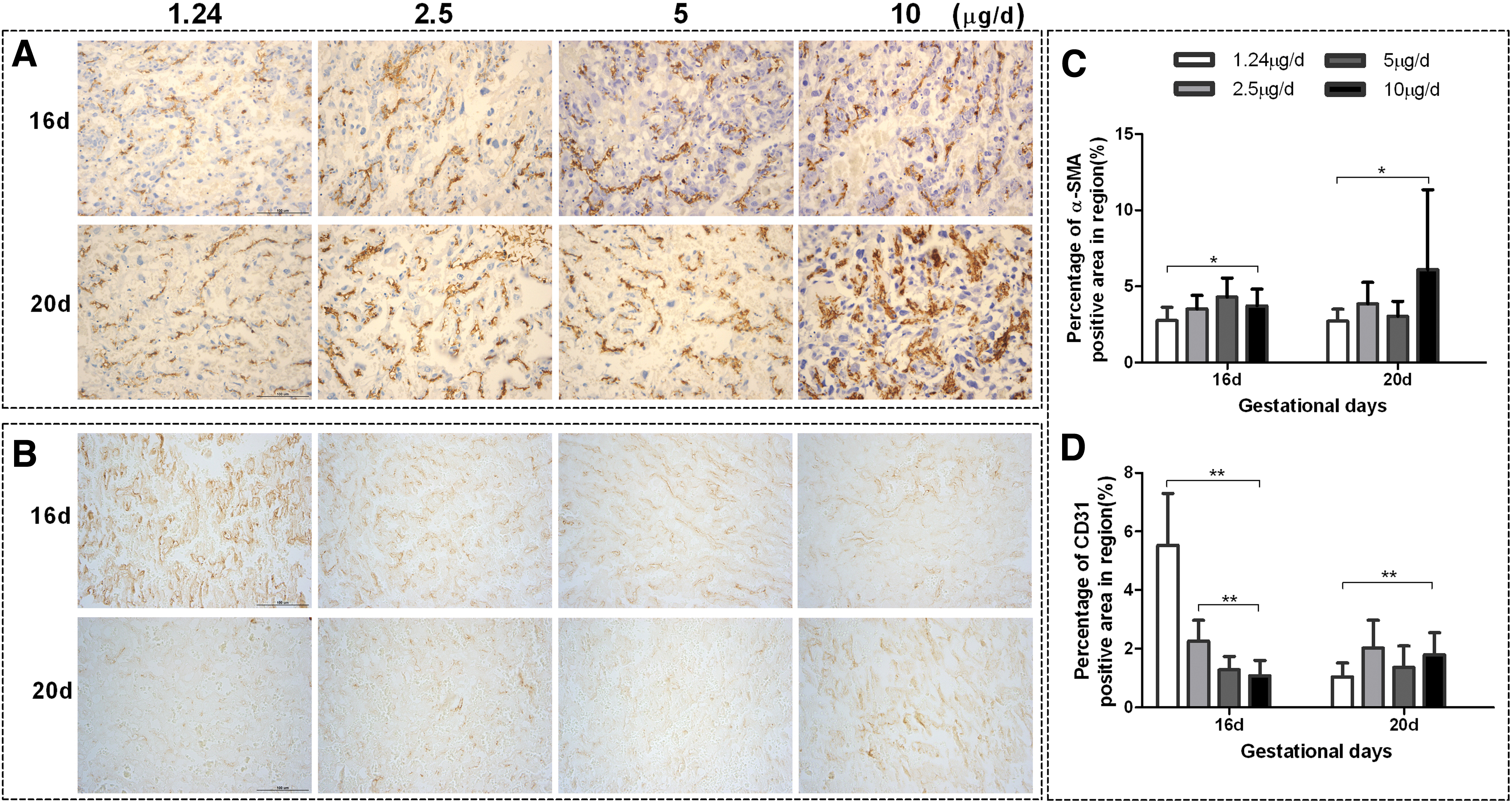
Representative images of α-SMA and CD31 immunoreactivity in placental labyrinth zone on two gestational days. The distribution of α-SMA (
Maternal serum rCG concentration
Significant differences in serum rCG concentration were not observed among the four groups (Supplementary Fig. S2).
Discussion
The fetus receives iodide from maternal circulating iodide and from deiodination of iodothyronines within the placenta. In our previous study, iodine was excreted through urine to maintain a relatively stable blood iodine content under mild-to-severe iodine deficiency (12). In the present study, increased thyroid weight, many smaller follicles, hyperemia, and increased active NIS membrane protein expression in a dose-dependent manner were observed in the thyroids of pregnant rats in the 1.24-, 2.5-, and 5-μg/day iodine groups on gd16 and gd20, indicating that the dams were in a state of severe-to-mild iodine deficiency. The association of placental iodine level with dietary iodine intake was inconsistent in previous reports (16,17). In the present study, differences in iodine levels in placental tissue were not observed among the four groups on gd16 and gd20. Thus, the iodine supply to the fetus depends on maternal blood iodine levels under severe-to-mild iodine deficiency; rats in the 1.24-, 2.5-, and 5-μg/day iodine groups on gd16 and gd20 exhibited ∼10–21%, 41–42%, and 50–76%, respectively, of iodine levels in the control group at the same time points.
Estimating fetal iodine intake by detecting iodine levels in the blood and urine of fetal rats is difficult; thus, the AF iodine level was used as a substitute for fetal iodine intake in the present study. In rats, day 16 of pregnancy is equivalent to the early stages of pregnancy in humans. Because the placenta and fetal vessels develop during this period, water and solutes from maternal plasma pass across the placenta to the fetus, and AF composition is similar to that of fetal plasma (18). The results of the present study show that the AF iodine levels in all the groups were similar on gd16, indicating that the fetal iodine level was minimally affected when iodine supply decreased by 10–50% in maternal blood.
The rat fetal thyroid begins to function at the end of the embryonic day 17. The capacity of thyroid tissue to concentrate 131I rapidly increases between days 17 and 22 after fertilization (19,20). Therefore, regardless of maternal iodine intake on gd16, the fetus attempts to maintain relatively stable iodine levels, and the placental barrier plays a crucial role in this process.
In the present study, NIS identified in rat placenta was observed to be ∼60 kDa in size compared with the ∼100 and ∼75 kDa form previously observed in rat thyroids and in lactating breast, respectively (21). However, the unglycosylated forms observed in above tissues were found at ∼50 kDa, suggesting differential glycosylation in thyroidal, lactating breast, and placental NIS (22). The upregulated pendrin and glycosylated NIS proteins facilitate mother-to-fetus iodide transfer in the placental labyrinth. The NIS protein transforms from a nonglycosylated to a glycosylated form, indicating that there is enhanced iodide-pumping activity in placental tissue during iodine deficiency. This change in form is critical for the NIS protein to perform its function, as reported in several cancer studies (23,24).
NIS protein and mRNA levels were reportedly stimulated by human chorionic gonadotropin in vitro in a previous study (25). Increased NIS and pendrin proteins in the present study were not associated with rCG levels because serum rCG did not change across the different groups. The iodide level is the main regulating factor in placental iodine transport (6). The NIS compensation for iodine shortage represents post-transcriptional regulation because its mRNA level was unchanged in the placental labyrinth.
However, NIS mRNA expression was reportedly induced in the fetal side of the placenta under a low-iodine diet (11). In addition, NIS is expressed in various placental and ovular tissues (26). Due to differing proportions of trophoblastic and nontrophoblastic cells in placental tissues and previously published results, we further investigated NIS mRNA levels in a trophoblast cell population isolated using LCM. However, mRNA levels in the trophoblast cells did not differ significantly, although a slight upregulation was observed in the lowest iodine intake group. In addition to the increased pendrin protein levels in three experimental groups, PDS mRNA was notably upregulated in trophoblast cells isolated using LCM under severe iodine deficiency but not in the placental labyrinth in the inadequate iodine intake groups. In other words, pendrin was significantly activated at the transcriptional level with gradually decreasing iodine intake.
Furthermore, more vascular elements labeled by CD31 and less villous stroma labeled by α-SMA were detected on gd16 in the fetal side of the placenta when iodine intake was reduced to 2.5 or 1.24 μg/day. CD31, an endothelial marker, was used for the study of villositary vascularization (27). The embryonic vascularization of the placental labyrinth layer can increase the exchange area to transmit more essential nutrients besides iodide under moderate-to-severe iodine deficiency.
Upon immunostaining for α-SMA in normal placental specimens, α-SMA was found in myofibroblasts located around the majority of fetal vessels, smooth muscle cells, pericytes of fetal blood vessel wall, and in the villous stroma mainly of stem and peripheral villi (intermediate immature and mature villi) (28 –30). The contraction/relaxation of α-SMA+ cells in placental villi would have implications not only in the regulation of blood flow within the placenta but also in the regulation of the length and width of the villi in the intervillous space (31). Thus, as shown in the present study, the deficient expression of both stromal and vascular α-SMA within the labyrinth can shorten the crossing distance of iodide and regulate placental blood flow to accelerate mother-to-fetus iodide delivery under severe iodine deficiency. Notably, decreased α-SMA expression and increased CD31 expression also indicate defective myofibroblastic differentiation and fetal vessel development in the villous stroma and stromal vessels, implying that the ability to regulate nutrient exchange is limited under iodine-deficient conditions.
In rats with a normal iodine intake, the placental expression of NIS mRNA and protein decreased and that of pendrin mRNA and protein increased from gd16 to gd20, which were consistent with results of previous in vitro experiments (7,32). Compared with rat placentas before the onset of fetal thyroid function, the regulatory effects of the fetal side of an iodine-deficient placenta on iodine transfer from mother to fetus in late pregnancy are still partly effective. For example, the expression of pendrin and glycosylated or nonglycosylated NIS proteins remained high, and α-SMA+ area was mildly to severely reduced in the three inadequate iodine dose groups.
However, the NIS protein transformation from the nonglycosylated to the glycosylated form was not significant and different from the increases observed in early pregnancy, whereas the CD31+ area was obviously reduced in the severely deficient iodine intake group. These differences indicate that the protective mechanisms of NIS and embryonic vascularization against the effects of iodine deficiency in late pregnancy are weaker than in early pregnancy.
Syncytiotrophoblast cells, rather than cytotrophoblast cells, are the major cellular components responsible for the exchange of substances in the placental barrier during late pregnancy (33). Thus, pendrin may be more important in late pregnancy than in early pregnancy regardless of whether the iodine intake is reduced or not. The results in the present study are consistent with previous findings—the expression of human PDS gene is higher in term than in first trimester pregnancy samples, and in early villous trophoblast, pendrin expression increases with syncytiotrophoblast formation (8,34). Furthermore, NIS gene expression is the limiting step for the onset of thyroid function in the human fetus (35). Thyroid NIS, rather than placental NIS, plays an important role in regulating iodine levels in the fetus after the maturation of the fetal thyroid. Therefore, the expression of placental NIS decreases with increasing gestational age, and the compensatory defense against iodine deficiency weakens.
Under mild and moderate iodine deficiency, α-SMA+ areas were smaller in late pregnancy than in early pregnancy, similar to those observed under severe iodine deficiency. Furthermore, the proportion of fetal capillaries within the labyrinth zone decreased due to a maternal diet severely deficient in iodine compared with the control or other treatment groups on gd16, which could impair angiogenesis in that layer. In a previous study, hypothyroidism induced by propylthiouracil reduced the area of vascular endothelial growth factor expression in the placental labyrinth on gd19 (36). In other words, the development of α-SMA+ and CD31+ cells in the placental labyrinth was inhibited by the duration and severity of maternal iodine deficiency. Therefore, placental function affected iodide transfer after the establishment of fetal thyroid function.
As the pregnancy progressed to gd20 and fetal thyroid function was established, AF iodine levels in the 10-μg/day iodine group increased approximately twofold from gd16 to gd20. In addition, AF iodine levels on gd20 in the 1.24-, 2.5-, and 5-μg/day groups were ∼39%, 73%, and 97%, respectively, of those in the control group. The range of change in AF iodine was much greater compared with that on gd16 and much lower compared with maternal blood iodine on gd20. Furthermore, AF and maternal thyroid iodine changed similarly, and the increase in fetal thyroidal NIS protein levels in the plasma membrane with decreased iodine doses was accompanied by smaller thyroid follicles, indicating that the pups were in a state of iodine deficiency.
In a previous study, NIS mRNA increased approximately twofold in fetal thyroids due to a low-iodine diet on gd21 (11). Furthermore, increased stimulation of the pituitary–thyroid axis and a relatively larger thyroid size was observed from gd18–19 through the first postnatal day in the fetuses of rats fed a low-iodine diet (37). A relatively larger thyroid size was also observed in fetal thyroids in the severe iodine deficiency group on gd20 in our previous study (35).
Terminal differentiation of the thyroid tissue is characterized by follicle formation and onset of thyroid hormone synthesis. As our published data (12), no matter what thyroid dysfunction of dams experienced, serum total thyroxine and free thyroxine levels of pregnant rats gradually declined as a result of mild-to-severe iodine deficiency at both gd16 and gd20. In pup AF, however, hypothyroidism and subclinical hypothyroidism were observed in the fetus of the severe iodine deficiency group on gd20 and gd16, respectively, while no obvious changes were observed in fetal thyroid function from the mild-to-moderate experimental groups at these two stages. Thyroid hormone transporters and deiodinases of placental tissue participated in the compensatory regulation to maintain the normal thyroid function of the fetus when maternal thyroid function was abnormal. Therefore, the formation of fetal thyroidal follicles was blocked in rats fed the lowest iodine diet, and thyroid development was inhibited due to severe fetal hypothyroidism, during which thyroidal iodine uptake further deteriorated.
Although numerous protective mechanisms in the placental barrier and fetal thyroid contribute to the effects against iodine deficiency from gd16 to gd20, the available iodine for the fetal thyroid is still insufficient for the synthesis of thyroid hormone even under mild iodine deficiency (11). The gradual increase in thyrotropin levels in the AF at gd16 and gd20 after a decrease in the dose of iodine supplements (12) and inhibition of fetal neuronal and synaptic growth on gd20 also confirm the above statement (38).
In conclusion, extensive adaptive and/or protective mechanisms in the placental barrier against the effects of iodine deficiency are already present before the functional establishment of iodine uptake in the fetal thyroid. Among these, the glycosylation modification of placental NIS protein and increased fetal capillary area play a crucial role in maintaining fetal iodine levels. When the fetal thyroid is fully functioning, it controls the direction of iodine flow, and the placenta may partly lose control of iodine delivery to the fetus in early pregnancy. Therefore, increased pendrin protein, rather than NIS, may play a pivotal role in the mediation of placental iodide transport.
Footnotes
Acknowledgment
We thank Tianjin Lung Cancer Institute at Tianjin Medical University General Hospital for assistance with acquired trophoblastic cell populations by Laser Microdissection and Pressure Catapulting.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (grant numbers: 31570178 and 81573103).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2