
Abstract
Background:
Maternal exercise (ME) improves fetal and offspring muscle development, but mechanisms remain to be established. Since the thyroid hormone (TH) is critical for cell differentiation during embryonic development, we hypothesized that ME elevates TH receptor (THR) signaling in embryos, which promotes embryonic myogenesis.
Methods:
Female mice were exercised daily on a treadmill or received a daily TH, triiodothyronine (T3) injection. Embryos (embryonic day 12.5 [E12.5]) and P19 cells were used for studying effects of TH on embryonic myogenesis. TH levels in serum and embryos after ME or T3I were analyzed. Expression of TH signaling related genes and myogenic genes was assessed. THRα binding to the promoters of myogenic genes was investigated by chromatin immunoprecipitation-qantitative polymerase chain reaction (ChIP-qPCR). A CRISPR/CAS9 plasmid was utilized to knock out THRα in P19 cells.
Results:
ME elevated TH levels in both maternal circulation and embryos, which were correlated with enhanced TH signaling and myogenesis. At E12.5, both myogenic determinants (Pax3, Pax7) and myogenic regulatory factors (Myf5, Myod) were upregulated in ME embryos. ME increased THRα content and elevated messenger RNA (mRNA) expression of TH transporter Slc16a2 and deiodinase Dio2. In addition, the THRα binding to the promoters of Pax3/7 was increased. In P19 embryoid bodies, T3 promoted myogenic differentiation, which was abolished by ablating THRα. Furthermore, maternal daily injection of T3 at a level matching exercised mothers promoted embryonic myogenesis.
Conclusions:
ME promotes TH delivery to the embryos and enhances embryonic myogenesis, which is partially mediated by enhanced TH signaling in ME embryos.
Introduction
While conflicting reports exist, increasing evidence points to the beneficial effects of gestational exercise on maternal and neonatal outcomes (1,2). The American College of Obstetricians and Gynecologist recommends that pregnant women participate in daily exercise during gestation (3). Unlike exhaustive exercise that causes adverse physiological responses (4), moderate exercise improves metabolic health (5,6). Similarly, gestational exercise also improves embryonic development. In mice, regular exercise during pregnancy remarkably reduced diet-induced obesity in their offspring (7), and suppressed metabolic dysfunctions (8 –10). Placenta mediates maternal and fetal exchange of nutrients (11), and maternal exercise (ME) enhanced placental vascularization and function, partially explaining the improvement of fetal development (7).
The thyroid hormone (TH) regulates muscle contraction, intestinal activity, and body temperature (12). Gestational thyroid dysfunction is quite common in women (13), which is associated with fetal growth retardation, adverse neurocognition (14), and miscarriage (15). Importantly, embryos do not produce TH and thus depend on maternal provision of triiodothyronine (T3) and thyroxine (T4) (16). The transcription of thyroid hormone receptor (THR) and transporter is initiated soon after fertilization in mice (17,18), but embryonic thyroid glands only function after embryonic day 17 (E17) in mice (19,20), emphasizing the importance of maternal TH in regulating embryonic development (21). Interestingly, recent studies showed that exercise could substitute TH treatment for hypothyroidism therapy (22 –25), because aerobic exercise increases circulatory TH levels (5,6,26,27).
While T4 is dominant in circulation, it is converted to active T3 in tissues locally by iodothyronine deiodinase 2 (D2 for protein and Dio2 for gene). The intracellular T3 level is regulated by D2 and D3. D2 converts T4 to T3 to activate THR signaling, while D3 inactivates both T4 and T3 through inner ring deiodination (28). D1 can catalyze both the outer and inner ring deiodination. The active T3 enters the cell nucleus and binds with THR, which, in conjunction with retinoid X receptor, further binds with its cognitive elements in promoters to initiate gene expression (29). There are two types of THR, THRα and THRβ, differentially expressed in tissues, with THRα predominantly expressed in skeletal muscle (30,31). However, despite a positive correlation between TH and muscle regeneration being reported (29,32,33), little is known about the roles of maternal TH in embryonic myogenesis.
Skeletal muscle, originated from the mesoderm, is one of the major metabolic organs (34). The number of skeletal muscle fibers is determined in utero, while after birth, the muscle growth mainly depends on hypertrophy (35). Prenatal muscle development is crucial for long-term postnatal function of skeletal muscle. In mice, the embryonic myogenesis occurs in the dermomyotome between E8.5 and 12.5, when embryonic Pax7+ myoblasts commit to a myogenic fate, and the myogenic factor Myf5 initiates myogenesis (34). The primary muscle fibers formed during the embryonic stage serve as scaffolds for the formation of fetal muscle fibers; these embryonic myogenic cells also contribute to the formation of satellite cells in offspring muscle (34,36,37). Thus, malformation of embryonic myogenic cells worsens fetal muscle development and offspring muscle function (38).
Indeed, in our previous study, we found that ME enhanced muscle mass and endurance in offspring (39). Considering that ME enhances TH levels, we hypothesized that ME improves the delivery of maternal TH into the embryos, which activates TH signaling and enhances embryonic myogenesis.
Materials and Methods
Mouse exercise protocol
Animal studies were conducted according to the protocol approved by the Institute of Animal Use and Care Committee at Washington State University. A total of 32 eight-week-old C57BL/6J female mice (The Jackson Laboratory, Bar Harbor, ME) were randomly separated into control (CN) and ME groups (16 females/group). A detailed exercise protocol has been described previously, and moderate exercise intensity is defined as 70% maximal oxygen consumption (VO2max) (7). The ME mice were assigned to prepregnancy exercise for 24 days and then mated with age-matched males. Successful mating was confirmed by the presence of vaginal plug and designated as E0.5. To synchronize pregnancy, all female mice were only with males for one night. Pregnant mice continued to be exercised during pregnancy until embryo collection at E12.5.
All mice were housed at 22°C on 12-hour-light/12-hour-dark cycles, fed ad libitum. The food intake and body weight were measured. At E12.5, pregnant mice were anesthetized by carbon dioxide inhalation and euthanized by cervical dislocation. The somite number of embryos was counted under a stereomicroscope. The head of embryo was removed to minimize the noise from neurogenic tissue (40), except those embryos used for TH analysis, where the whole embryos were used.
T3 hormone injection
Sixteen 8-week-old C57BL/6J female mice (The Jackson Laboratory) were randomly separated into CN and thyroid hormone injection (T3I) groups (8 females/group). After mating, pregnant mice were subcutaneously injected with T3 (0.2 μg/g body weight) in saline solution daily (41,42), while CN mice received an equal volume of saline solution until dissection at E12.5.
Muscle function measurement
Two days before mating, the VO2max, acute exercise endurance, and total running distance were measured using a treadmill respiratory measurement system (Columbus Instruments, Columbus, OH). A grip strength meter for mice (Columbus Instruments) was used to measure the limb grip strength (7).
Metabolic studies
At gestational day (GD) 9.5, pregnant mice were subjected to a resting metabolic rate analysis, including oxygen consumption (VO2) and heat production (from 10 AM to 6 AM the next day) using an indirect open circuit calorimetry system (Comprehensive Lab Animal Monitoring System [CLAMS]; Columbus Instruments) (7).
TH analysis
Total T3, T4, and thyrotropin (TSH) in maternal serum and whole embryos were analyzed using a Thyroid Panel enzyme-linked immunosorbent assay (ELISA) Kit (Monobind, Inc.). For maternal serum separation, whole blood was clotted for 8 hours at 4°C, and then centrifuged at 2000 × g for 10 minutes to remove debris; the supernatant was transferred to a new tube. To detect the TH in the whole embryos, an extraction buffer included in the ELISA Kit was added to the embryos (300 μL/50 mg tissue) and homogenized at 4°C. The debris was removed by centrifuge (5424 R, Eppendorf, Enfield, CT) at 12,000 rpm for 5 minutes at 4°C (16).
P19 cell culture
An embryonic cell line, P19 (CRL-1825, ATCC), was used to examine the embryonic myogenesis, as described in a previous report (38). Briefly, P19 cells were cultured in a growth medium (alpha-minimal essential medium [a-MEM] with 7.5% bovine calf serum, 2.5% fetal bovine serum, and 0.5% penicillin–streptomycin; Sigma, Burlington, MA) at 37°C in 5% CO2. For embryoid body (EB) culture, a total of 5 × 106 P19 cells were subcultured into a Petri dish (VWR International, Irving, TX) with growth medium and 1% dimethyl sulfoxide, and placed in an incubator with shaker (Incu-Shaker Mini; Benchmark, Woburn, MA) at 70 rpm. In addition, 10 nM T3 (TOCRIS Bioscience, Minneapolis, MN) was added to assess the effects of TH on myogenesis (30,43).
Quantitative reverse transcription polymerase chain reaction analysis
RNA was extracted using TRIzol reagent (Invitrogen, Grand Island, NY) and reverse transcribed to complementary DNA (cDNA) by an iScript™ cDNA Synthesis Kit (Bio-Rad, Hercules, CA). SsoAdvanced™ Universal SYBR Green Supermix (BioRad, Hercules, CA) was used for polymerase chain reaction (PCR) with a Real-Time System (BioRad). All the primers used are listed in Table 1. 18S ribosomal RNA (rRNA) was used as a reference gene and relative RNA levels were calculated by the 2−ΔΔCt method (44).
Primers Used in This Study
Dio2/3, deiodinase 2/3; qRT-PCR, quantitative reverse transcription polymerase chain reaction; ChIP-qPCR, chromatin immunoprecipitation-quantitative polymerase chain reaction.
Western blot analysis
Proteins were extracted as previously described (39). The primary antibody against THRα was purchased from Abcam (Waltham, MA; ab53729), and the secondary goat anti-rabbit antibody (No. 926-32,211, RRID: AB_621,843) and goat anti-mouse antibody (No. 926-68,070, RRID AB_10,956,588) were purchased from LI-COR Biosciences (Lincoln, NE). Primary antibodies against β-actin (AB_528068) and tubulin (AB_2315513) were purchased from DSHB (Iowa City, IA) and used as references. Images were captured by an Odyssey Infrared Imaging System (LI-COR Biosciences) and quantified by ImageJ software (National Institutes of Health) (45).
Chromatin immunoprecipitation-quantitative polymerase chain reaction
Upstream sequences of each gene were obtained from the Swiss Institute of Bioinformatics Eukaryotic Promoter Database and analyzed by PROMO program (Version 8.3) (46,47). Chromatin immunoprecipitation (ChIP)-qPCR was carried out as previously described (48). Briefly, extracts were sonicated by a sonic dismembrator (Fisher Scientific, Waltham, MA) as follows: 30% intensity for 230 seconds with 5 seconds ON/5 seconds OFF. The sonicated DNA size was verified by agarose gel electrophoresis. Then, the THRα primary antibody (rabbit-anti-mice, ab53729; Abcam) or IgG (Sigma) was added, and immunoprecipitation (IP) was performed using EcoMagTM Protein A Magnetic Particles (No. MJA-102; Bioclone, Inc., San Diego, CA). Quantitative PCR was performed using SsoAdvanced Universal SYBR Green Supermix (BioRad, Hercules, CA) with System (BioRad). All the the primers used are listed in Table 1.
Immunocytochemical staining
P19 cells were incubated with a primary antibody against embryonic myosin heavy chain (MHC; F1.652-S; DSHB) and a fluorescent secondary antibody (6410-32, AB_2796308, donkey anti-mouse; SouthernBiotech, Birmingham, AL). A mounting medium with DAPI (Abcam) was added, and cells were observed by EVOS XL Core Imaging System (Mil Creek, WA) (49).
THRα knockout in P19 cells
A CRIPSR/Cas9 plasmid was used to knock out THRα in P19 cells. The crRNA (5′-CCTGGGCGTTAGTGCTGCGA-3′) targeting THRα was cloned into a plasmid with ampicillin bacterial resistance and puromycin selective marker (PX459; GenScript, Piscataway, NJ). The same vector with scrambled sequence was used as a CN. Plasmids were transfected into P19 cells using Lipofectamine 3000 (Invitrogen, Lenexa, KS). After transfection, P19 cells were selected with 3 μg/mL puromycin dihydrochloride (Fisher Scientific) for 24 hours (50,51).
Statistical analysis
All data were visualized and analyzed by Prism GraphPad (V.7.0, San Diego, CA), presented as mean ± standard error of the mean. Significance (p < 0.05) was identified using a two-tailed unpaired Student t-test. The correlation was analyzed by Pearson's chi-squared test. Embryos from a single pregnancy were pooled and a pregnancy was considered an experimental unit.
Results
Exercise improves metabolism and TH levels of pregnant mice
Mice were subjected to a 36-day moderate exercise training, including 12 days during gestation (Fig. 1A). Although the food intake did not change, the body weight gain was lower in the ME group before pregnancy (Fig. 1B). Interestingly, during gestation, starting from E8.5, the body weight gain (compared with E0.5) in the ME group was higher than in the CN group (Fig. 1B). After a 24-day exercise training, limb muscle maximal strength and muscle endurance of pregnant mice were significantly enhanced (Fig. 1C). Correspondingly, exercised mice had improved aerobic endurance capacity, indicated by increased VO2max, exercise duration, and running distance (Fig. 1D). In addition, exercised mice (at GD9.5) had increased VO2 and heat production during the active night phase (Fig. 1E, F). Importantly, ME had elevated T3 and T4 levels (Fig. 1G), whereas TSH level did not differ (Fig. 1H), suggesting that exercise increased circulatory TH levels.

Exercise improves metabolism and TH levels of pregnant mice. (
ME during pregnancy promotes TH delivery into embryos
We further analyzed the TH concentrations in embryos (E12.5). In agreement, both T3 and T4 levels were elevated in ME embryos (Fig. 2A). Accordingly, the messenger RNA (mRNA) contents of several TH-related genes, including TH transporter (Slc16a2), Thra, Dio2, and Dio3, were all elevated in the ME embryos (Fig. 2B) and positively related to the T3 level (Fig. 2C). Notably, the ratio of Dio2 to Dio3 was increased in the ME group, partially explaining the elevation of T3 in ME embryos (Fig. 2D). In addition, the THRα protein content in embryos was higher in ME embryos (Fig. 2E), consistent with elevation of TH-responsive genes, Pgc1a, Nrf1, and Myog (52 –54) in E12.5 (Fig. 2F). In short, ME improves TH delivery to the embryos, augmenting local TH signaling.
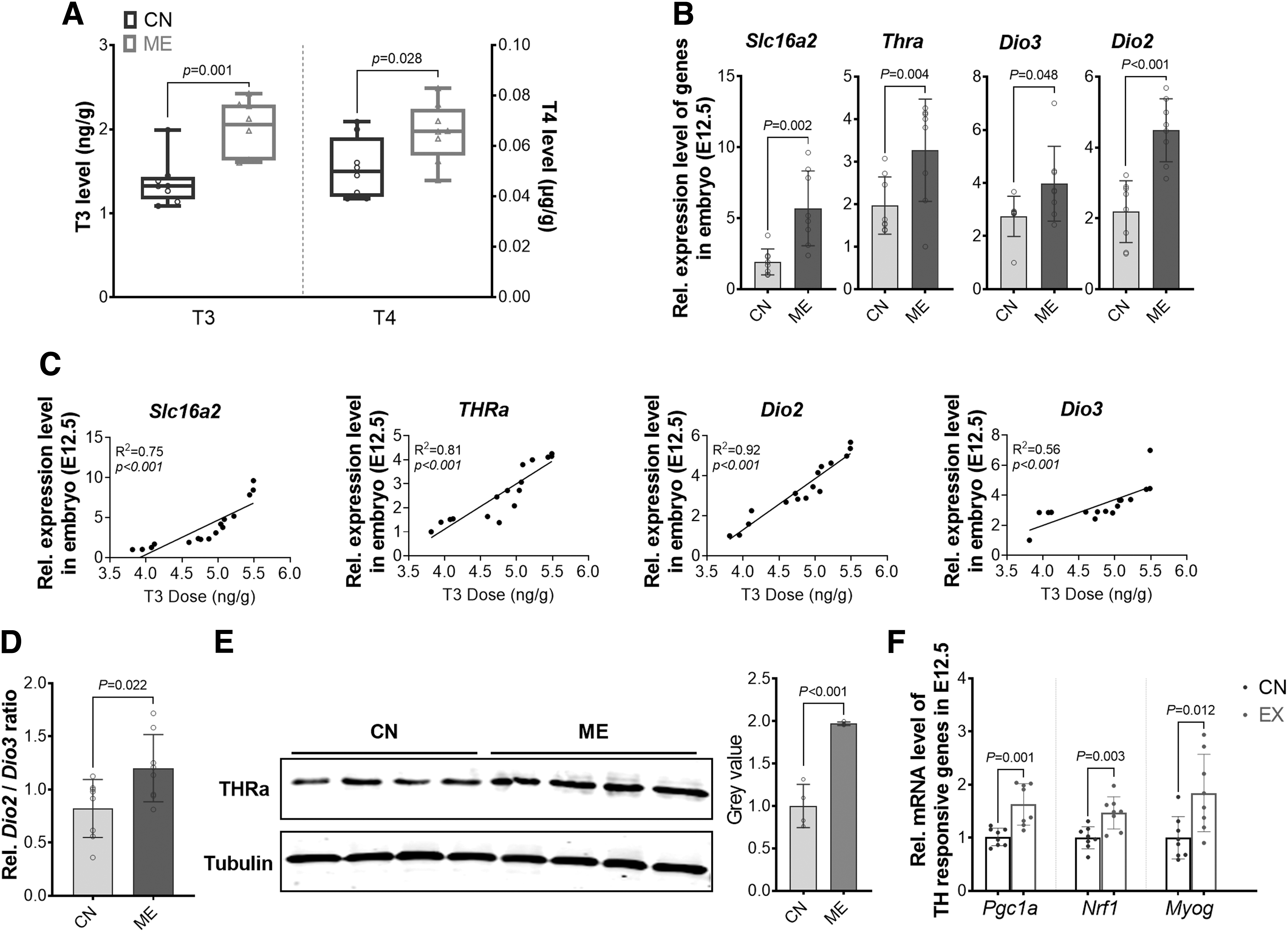
ME during pregnancy promotes TH delivery into embryos. (
ME during pregnancy promotes embryonic myogenesis
At E12.5, ME embryos had higher mass (Fig. 3A), consistent with the increased body weight gain of pregnant mice during gestation (Fig. 1B). The somite count, placenta weight, and number of embryos per litter did not differ between groups (Fig. 3B–D). For embryonic myogenesis, the expression of myogenic genes, including Myf5, Myod, Pax3, and Pax7, was upregulated in ME embryos (Fig. 3F) and positively correlated with T3 level (Fig. 3E). Based on bioinformatic prediction, four and six potential THRα binding sites (dissimilarity <5%) were predicted in the promoters of Pax3 and Pax7, respectively (Fig. 3G). By ChIP-PCR analysis, one THRα binding site located at −7574 bp of the transcription start site (TSS) of Pax3 and one THRα binding site located at −7519 bp of the Pax7 TSS were identified (Fig. 3H).
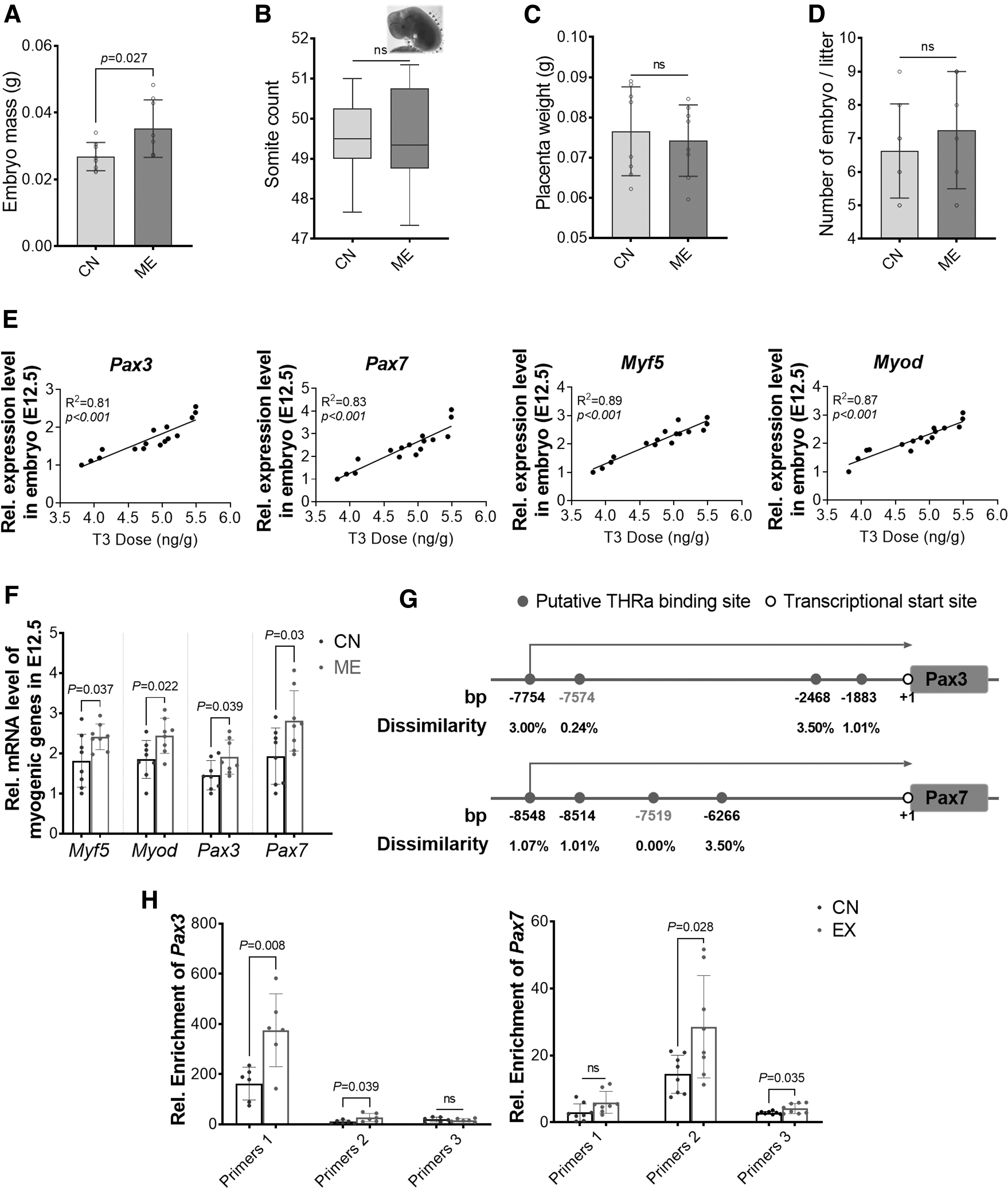
Maternal exercise during pregnancy promotes embryonic myogenesis. (
To sum up, ME facilitates myogenesis, which correlates with the upregulation of THRα signaling and expression of myogenic genes in E12.5 embryos.
TH facilitates myogenesis in P19 EB
P19 cells are capable to form EB and simulate embryogenesis in vitro (55). Here, T3 treatment (Fig. 4A) enhanced myogenic differentiation in P19 EB, indicated by immunostaining of embryonic MHC, where more myoblasts and myofibers were visible in T3-treated cells (Fig. 4B). Consistently, TH signaling was highly activated by T3 during myogenic differentiation, as shown by an increased expression of Thrα, Slc16a2, Dio2, and Dio3, as well as an elevated Dio2/Dio3 ratio (Fig. 4C). Furthermore, myogenic genes, Pax3, Pax7, Myf5, and Myod, were expressed at higher levels in the T3 group (Fig. 4D).
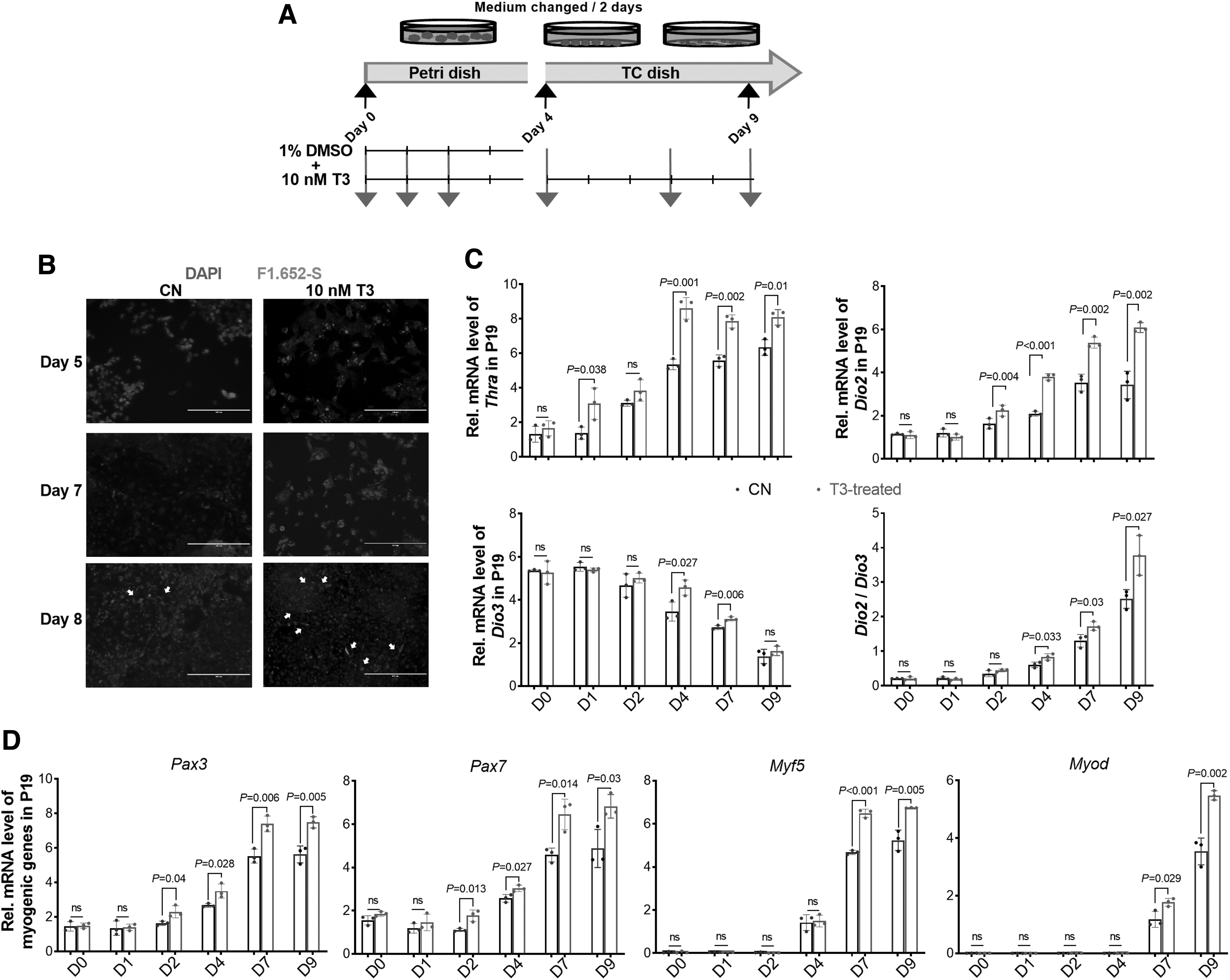
TH facilitates myogenesis in P19 EB. (
Knocking out THRα in P19 cells abolishes myogenic differentiation stimulated by T3
To analyze the mediatory role of THRα, we knocked out THRα, which was verified at the mRNA and protein levels (Fig. 5A, C). Interestingly, THRα-knock out (KO) P19 cells displayed impaired formation of EB, indicated by less EB numbers and smaller sizes (Fig. 5B). Furthermore, myogenic differentiation was almost abolished in THRα-KO P19 cells after an 8-day differentiation, shown by less myofiber formation (Fig. 5B) and decreased expression of myogenic genes (Fig. 5D). Despite THRα KO, the expression of Dio2 and Dio3 did not differ. Taken together, THRα expression is required for embryonic myogenesis.
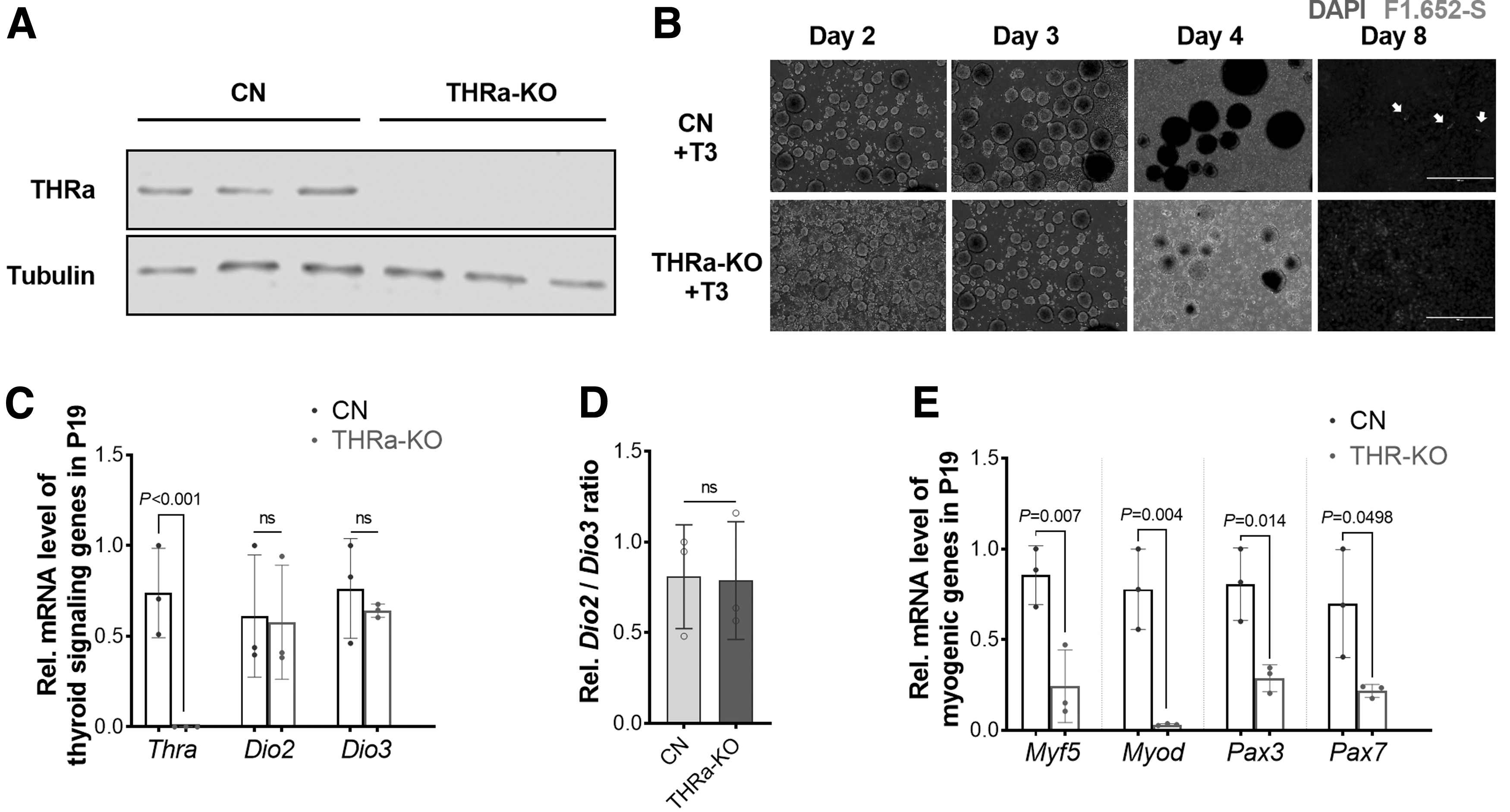
Knocking out THRα in P19 cells abolishes myogenic differentiation stimulated by T3. (
Daily injection of T3 during pregnancy enhances embryonic myogenesis
To further verify the mediatory roles of THR signaling in enhanced myogenesis due to ME, we injected pregnant mice with T3 daily (Fig. 6A). T3I did not alter food intake and body weight gain (Fig. 6B). As expected, T3 level in maternal serum increased after 3 days of T3I (Fig. 6D). Also, 12 days later, T3 maintained higher, while T4 and TSH levels retreated to the CN level (Fig. 6E). Correspondingly, embryos from T3-injected mice had a higher T3 concentration (Fig. 6F). There were no differences observed in embryo mass, placental weight, embryo number, and somite count (Fig. 6C). Similarly, the THRα protein was expressed higher in the T3I group (Fig. 6G), along with enhanced TH signaling (Fig. 6H). In agreement, the expression of myogenic genes was also elevated (Fig. 6I).

Daily injection of T3 during pregnancy enhances embryonic myogenesis. (
These data showed that maternal T3I replicates the beneficial effects of ME on embryonic myogenesis, confirming the mediatory roles of TH signaling.
Discussion
Regular moderate exercise benefits the mental and physical health of pregnant women (56,57) and improves the long-term metabolic health of offspring (2,58). A meta-analysis showed that ME improves fetal growth and prevents preterm birth (17,59,60). Improvement in VO2max due to regular exercise depends on exercise type, intensity, and duration (61), which correlates with metabolic health (62).
In our study, a 24-day regular treadmill exercise before mating increased the relative VO2max, along with improved muscle performance; the resting metabolic rate was also elevated, showing an overall improvement in the metabolic health of female mice. During gestation, these well-trained mice showed elevated body mass gain, indicating no stress reaction to exercise. Consistently, embryo mass was higher in the ME group. In summary, ME before and during pregnancy promotes healthy embryonic development.
TH secretion by the thyroid gland is under the regulation of the hypothalamus–pituitary–thyroid axis (63). Exercise stimulates TH secretion (64). In aged rats, exercise improved their TH secretion to a rate similar to young rats (65). Notably, elevation of TH secretion due to ME should be distinguished from pathological hyperthyroidism and thyroid disorders. ME-induced TH secretion is caused by elevated metabolism, which is a healthy self-adaptation (66). To mimic TH elevation due to exercise, we used T3I. In rodent studies, T3I was used to induce hyperthyroidism (67,68). The decreased T4 and TSH after a 3-day T3I suggested a risk of hyperthyroidism, although their levels mostly recovered after 12 days of injection. Consequently, T3I pregnant mice did not show symptoms of hyperthyroidism, nor altered body weight or food intake. No abnormal activities such as anxiety and weakness were observed.
However, T3I did not increase the embryo mass as ME did, suggesting that ME improved embryonic development not entirely dependent on the elevation of TH signaling. Indeed, regular moderate exercise enhances placental functions (7,69), which might improve nutrient delivery and embryonic development (70). Overall, our data showed that maternal T3I elevated T3 in embryos to a comparable level with ME, which facilitated embryonic myogenesis. The molecular mechanism of increased TH secretion due to exercise remains unclear (64), but exercise enhances thyroid gland activity and increases iodine utilization (71,72). Other metabolic organs such as the brown adipose tissue, liver, and muscle also regulate circulatory TH levels (27). Our data show that a 36-day moderate exercise elevates serum T3 and T4 concentrations in pregnant mice, as well as in embryos.
TH is essential for myogenesis, a process regulated by a hierarchical expression of transcription factors (54). Among them, Pax3, Pax7, and Myf5 commit progenitor cells to the myogenic fate. After that, Myf5 activates the expression of other myogenic regulatory factors, including Myod, Myog, and Mrf4, which induce terminal myogenic differentiation (34,73). During myogenesis, a sufficient reservoir of TH is required for proper cell differentiation (30,74,75). T3 activity is positively related to the expression of myogenic genes (76), while KO of THRα dramatically inhibited myogenic differentiation and muscle regeneration (77). The intracellular concentration of T3 is regulated by extracellular TH, transmembrane transporters, and T4 to T3 conversion (19,78). It is worthy to mention that the ratio of Dio2 to Dio3 was increased after ME, consistent with an increased T3 level in embryos, which facilitates myogenesis. In addition, our data showed that THRα binds to the Pax3 and Pax7 promoters, which was enhanced due to ME. Similarly, previous studies found that THRα binds with the Myod and Myog promoters in C2C12 myogenic cells (29,73).
Taken together, we discovered a TH-mediated mechanism in enhancing embryonic myogenesis due to ME. ME enhances maternal TH delivery into embryos, upregulating TH signaling and promoting embryonic myogenesis (Fig. 7). Due to the high accessibility of exercise and the pervasiveness of a sedentary lifestyle in pregnant women, our findings have important implications related to improving fetal development and offspring muscle health.

Summary of schematic diagram. Maternal treadmill exercise elevates the circulatory TH level in pregnant mice and embryos (left panel). The increased TH provides sufficient reservoir for proper embryo development (right panel). In E12.5 somite (black line encircled), where myogenesis initiates, ME increases the expression of Dio2, which elevates T3 level, and enhances THRa binding with the promoters of myogenic genes, facilitating embryonic myogenesis. The schematic art pieces used in this figure were provided by Servier Medical art, under a Creative Commons Attribution 3.0 Unported License. .
Footnotes
Authors' Contributions
Y.G. designed and performed the experiments, analyzed the data, and wrote and edited the article. L.Z. was involved in collecting the embryos. J.S.S. provided the main idea for exercise intervention. X.L. and Y.C. were involved in analyzing and interpreting the data. J.M.D. performed animal care and metabolic measurement. M.-J.Z. and G.K.M. interpreted the data and revised the article. M.D. designed the studies, interpreted the data, and edited the article.
Acknowledgment
We thank Servier Medical Art for the artwork.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This study is supported by the National Institute of Health R01-HD067449 to M.D.