
Abstract
Background:
Recent studies have reported a wide spectrum of ocular surface injuries in the context of autoimmune reactions in Graves' orbitopathy (GO). Increased expression of inflammatory mediators in tears of GO patients suggests that the lacrimal glands could be a target for immune responses. However, the immunophenotype for GO lacrimal microenvironment is not known. This study aimed to elucidate the pathological changes of GO lacrimal glands.
Methods:
In this case–control study, lacrimal glands were surgically collected from GO patients who underwent orbital decompression surgery and control subjects who underwent other ocular-related surgery. Bulk RNA-sequencing, flow cytometry with dimensional reduction, and immunohistochemical and multiplexed stainings were conducted. Western blotting and multipathway assays were performed in CD34+ fibroblasts derived from lacrimal and orbital tissues.
Results:
Increased expression of cytokines and chemokines accompanied by a variety of immune cell infiltrations mainly involving T cells, B cells, and monocytes was found in GO lacrimal glands. An in-depth investigation into T cell subsets revealed interferon (IFN)-γ-producing T helper (Th)1 and interleukin (IL)-17A-producing Th17 cell-dominated autoimmunity in the active GO lacrimal microenvironment. Both fibrosis and adipogenesis were observed in GO lacrimal tissue remodeling. IL-17A, not IFN-γ, stimulated transforming growth factor-β-initiated myofibroblast differentiation as well as 15-deoxy-Δ12,14-prostaglandin J2-initiated adipocyte differentiation in CD34+ lacrimal fibroblasts (LFs) and orbital fibroblasts (OFs), respectively. IL-17A activated many fibrotic and adipogenic-related signaling pathways in CD34+ LFs and OFs. A novel anti-IL-17A monoclonal antibody SHR-1314 could reverse the promoting effect of IL-17A on fibrosis and adipogenesis in CD34+ LFs and OFs.
Conclusions:
Our findings provide evidence for the infiltration of different lymphocytes into GO lacrimal microenvironment, where Th1 and Th17 cells characterize the onset of active lacrimal inflammation and contribute to tissue remodeling. These findings may have potential future therapeutic implications regarding the utility of anti-IL-17A therapy, which should be studied in future research.
Introduction
The primary lacrimal glands located within the upper temporal orbit produce the aqueous component of the tear film and play a key role in ocular surface health and visual quality (1). Graves' orbitopathy (GO), the most common extrathyroidal manifestation of Graves' disease, is usually associated with a wide spectrum of ocular surface changes (2,3). Lacrimal gland enlargement and dry eye syndrome in GO patients suggest ocular surface damage resulting from lacrimal gland dysfunction such as hyposecretion that may be caused by the inflammatory process and stimulation of lacrimal thyrotropin receptors by GO autoantibodies (4,5). The significant reduction in corneal surface epithelial cell density in GO patients is probably a secondary lesion due to dry eye syndrome, aggravating ocular surface inflammation (6).
Numerous studies have shown that T helper (Th)1-related cytokines such as interleukin (IL)-12 and tumor necrosis factor (TNF)-α, Th2-related cytokines such as IL-13, and Th17-related cytokines such as IL-1β, IL-6, and IL-17A appeared significantly higher in the tears of GO patients (7 –11). Furthermore, proteomics of GO tears demonstrated the imbalance between inflammatory and protective proteins (12,13). Some proteins, such as cystatin C, α-1-antichymotrypsin, retinal dehydrogenase, and lysozyme C (14,15), displayed different levels in GO tears, which may emerge as promising biomarkers for disease activity and ocular surface disease in GO. Hence, it is of great interest to acquire precise information on the immunopathological changes in lymphocytic distribution and function within the lacrimal glands in GO.
In this study, our aim was to investigate the immunophenotype for GO lacrimal glands and explore for interferon (IFN)-γ-producing Th1 and IL-17A-producing Th17 cell-dominated inflammation in the active GO lacrimal microenvironment. Understanding the altered regulation of autoimmunity in lacrimal glands will inform the potential future treatment for GO.
Materials and Methods
Study subjects
The study design is a case–control study. We screened 76 consecutive patients with GO, who underwent orbital decompression surgery at the Department of Ophthalmology, Shanghai Ninth People's Hospital, Shanghai JiaoTong University School of Medicine during the period from June 2020 to June 2021. Patients were eligible if they were 18 years or older, with active or stable GO, where the clinical activity score was evaluated by the same trained rater. The exclusion criteria were as follows: (i) autoimmune diseases, such as Sjögren's syndrome, asthma, and diabetes; (ii) history of exposure to chemical volatilization or strong light in their working environment; (iii) wearing contact lenses in nearly 15 days; (iv) other ocular surface diseases; or (v) current pregnancy or lactation.
All patients had stopped steroid or immunosuppressive agents for at least three months before surgery. We matched by age and sex 25 control subjects with lacrimal gland prolapse, blepharochalasis, or single eyelid, who underwent reconstructive ocular surgery.
The lacrimal glands and orbital connective tissues were collected intraoperatively. Blood samples were taken at the day before surgery. All participants signed informed consents, and the study was approved by the local institutional review board (SH9H-2019-T192-2) and conducted in accordance with the Declaration of Helsinki.
Statistical analysis
The data are presented as mean ± standard deviation or median with interquartile range. The two-tailed Student's t-test, one-way analysis of variance, nonparametric Kruskal–Wallis test, and chi-square test were used to perform comparison among groups, as appropriate. The SPSS Statistics (version 21.0) (International Business Machines Corporation) was used to analyze the data. A p-value <0.05 was considered statistically significant. A Bonferroni-corrected p-value <0.05 was considered to be a significant functional enrichment in the RNA experiments.
A detailed experimental scheme is provided in Supplementary Materials.
Results
A total of 51 eligible patients (26 active GO patients and 25 stable GO patients) were enrolled. The demographic and clinical characteristics of patients and controls are summarized in Table 1.
Patient and Control Demographics
F, female; GO, Graves' orbitopathy; IQR, interquartile range; M, male; SD, standard deviation; TRAb, thyrotropin receptor antibody.
GO lacrimal glands were infiltrated by different immune cell subsets
Our RNA-sequencing data showed that the gene expression of CD14 (monocytes), CD19 (B cells), CD3E (total T cells), CD8A (cytotoxic T cells), PTPRC (total leukomonocytes), KIT (mast cells), CD38 (plasma cells), CD4 (Th cells), and NCAM1 (natural killer cells) was highly elevated in the active group compared with the stable group and normal controls (Fig. 1A). T cells, B cells, and monocytes were the most remarkably increased immune cell subsets in active lacrimal glands (Fig. 1A). Further analysis of cytokine and chemokine gene expression showed that they were more concentrated in active group than the stable and normal groups. TNF, IL-1B, IL-16, PDGF, CCL2, CXCL9/10/11, and IL-6 were increased in active lacrimal glands (Fig. 1B), consistent with previous findings in orbital connective tissues (16). Th1 and Th17 cytokine genes IFNG and IL-17A were elevated in active lacrimal glands, while the Th2 cytokine gene IL-4 was higher in normal lacrimal glands (Fig. 1B).

Immune cell infiltration and cytokine and chemokine expression in lacrimal glands from GO patients and control subjects. (
These data suggest that the inflammatory responses were stronger in active lacrimal glands and that both Th1 and Th17 cells would be the mainly involved in T cell subsets in GO lacrimal inflammation.
GO lacrimal inflammation was characterized by IFN-γ-producing Th1 and IL-17A-producing Th17 cells
To confirm the findings at transcription levels, we examined the main immune cell lineages using flow cytometry. Several clusters were formed in the TriMAP maps according to the defined lineage markers and all clusters contained populations found in active, stable, and normal lacrimal glands (Fig. 1C–E). There were two clearly separated clusters of CD3+ cells, one uniformly coexpressing CD4, and the other with more variable levels of CD8. IFN-γ was variably expressed in both of these two clusters, while IL-17A was expressed mainly in CD4+ cluster and IL-4 was not differentially expressed in all clusters (Fig. 1E). The cluster located in close proximity to the CD4+ cells was characterized by a high expression of CD14, indicating monocytes (Fig. 1E). At the right of CD14+ cluster was CD19+ cells, suggesting that these were B cells (Fig. 1E).
Moreover, our data demonstrated that the expression of CD3, CD4, CD8, CD19, CD14, IFN-γ, and IL-17A was higher in GO lacrimal glands than control ones, especially in the active stage. The expression of IL-4 was much lower and there were no apparent differences among the three groups (Fig. 1F).
In line with the flow cytometry data, a histological study showed that CD3+ cells infiltrated GO lacrimal glands compared with normal ones, especially in the active stage (Fig. 2A). In addition, those infiltrative T cells in GO lacrimal glands included both CD4+ (Fig. 2B) and CD8+ (Fig. 2C) subsets, which significantly gathered focally in the active stage. Furthermore, the expression of Th1 and Th17 cytokines IFN-γ and IL-17A was much higher in active lacrimal glands (Fig. 2D, E), suggesting an early phase of GO lacrimal autoimmunity characterized by Th1 and Th17 cells. This was in agreement with previous findings in orbital connective tissues (17,18).
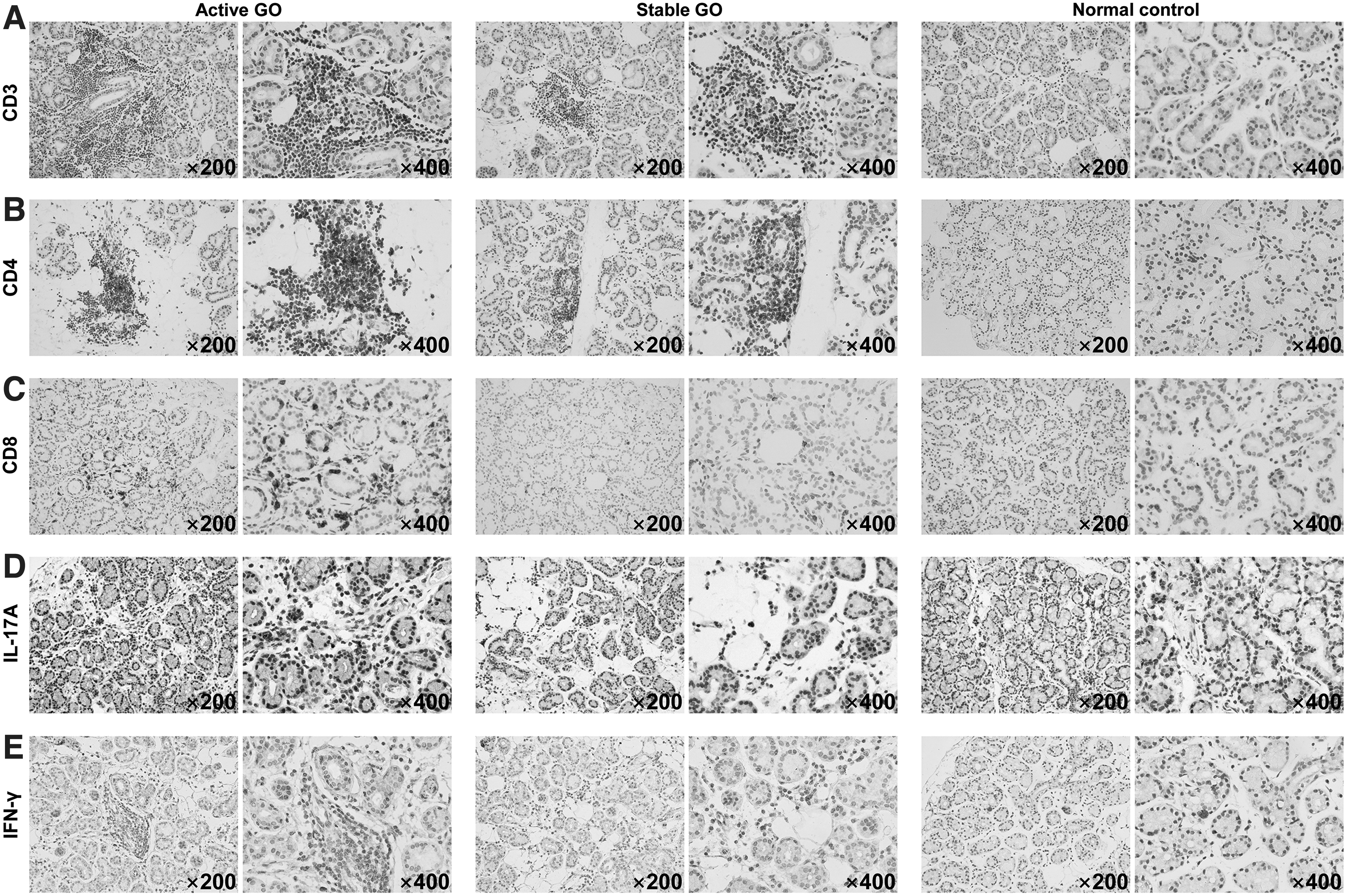
Immunohistochemical staining for lacrimal glands from GO patients and control subjects. The infiltration of CD3+ T cells (total T cells) (
Heterogeneous T cell subpopulations were observed in GO lacrimal glands
We next assessed the spatial distribution of T cell subpopulations in GO and control lacrimal glands by multiplexed immunohistochemistry (IHC). Our results showed the expected patterns of cell staining, with CD4 delineating cells morphologically consistent with Th cells, and CD8 delineating cells morphologically consistent with cytotoxic T cells (Fig. 3A). Tbet, GATA3, retinoic acid receptor-related orphan receptor (ROR)γt, and FOXP3 are key transcriptional factors for Th1, Th2, Th17, and regulatory T (Treg) cells, respectively (19). Their coexpression with CD4 and DAPI highlighted the presence of those T cell subsets (Fig. 3A, B) within GO lacrimal microenvironment, especially in the active stage (Fig. 3A). Both CD4+ and CD8+ T cells and the Th1, Th2, Th17, and Treg cell subsets were almost invisible in a normal lacrimal microenvironment (Fig. 3A).
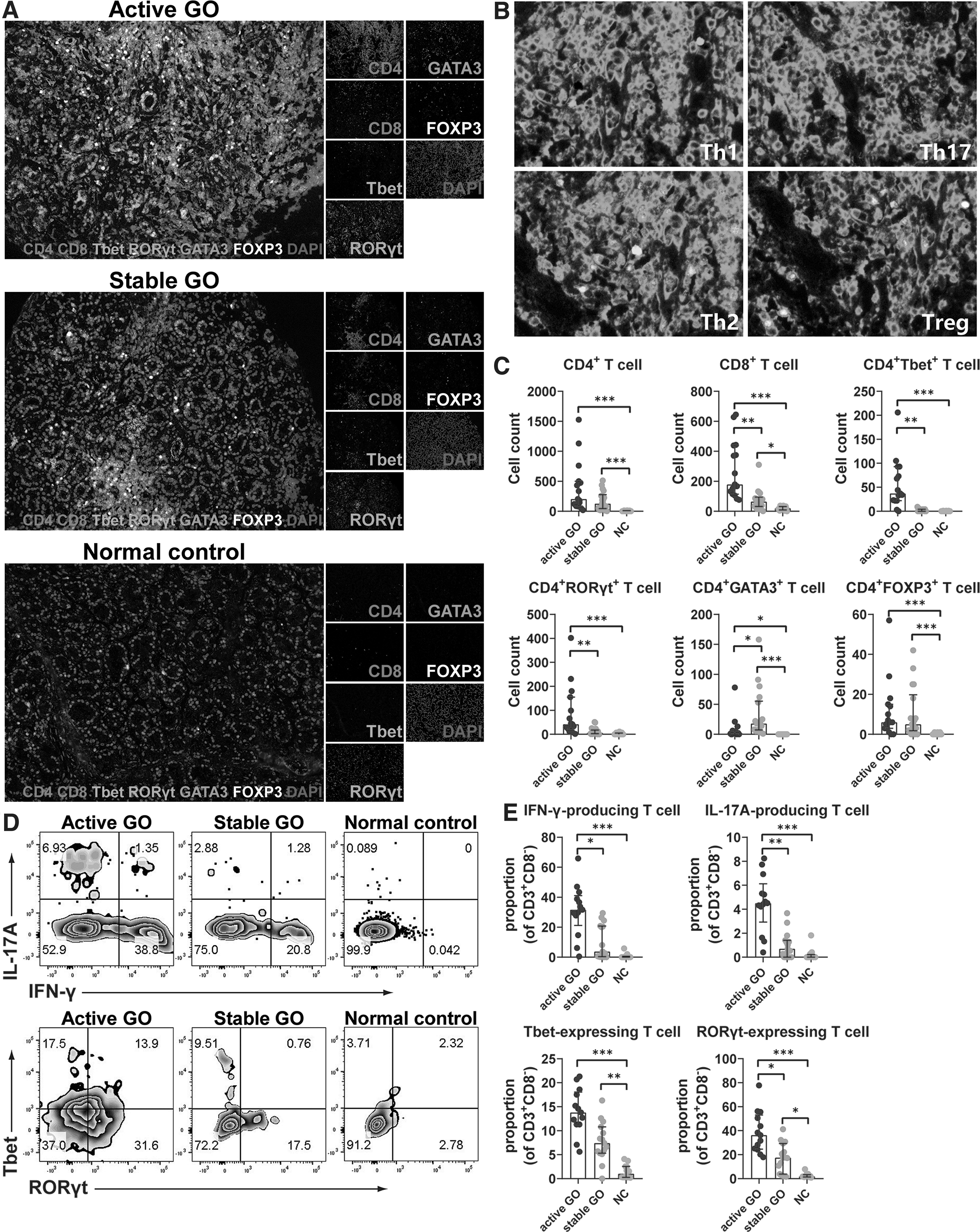
Spatial distribution of T cell subsets in lacrimal glands from GO patients and control subjects by multiplexed immunohistochemistry assay. (
Quantitative analysis demonstrated that there were a significantly increased number of CD4+ (active vs. control, 204.0 [85.0–500.0] vs. 1.0 [0.0–10.0], p < 0.001; stable vs. control, 128.0 [45.0–277.8] vs. 1.0 [0.0–10.0], p < 0.001) and CD8+ (active vs. control, 179.0 [114.0–439.0] vs. 20.0 [9.0–31.0], p < 0.001; stable vs. control, 64.5 [32.3–94.5] vs. 20.0 [9.0–31.0], p = 0.033) T cells in both the active and stable lacrimal microenvironments compared with normal control, and the number of CD8+ T cells in the active lacrimal microenvironment was higher than that in a stable one (active vs. stable, 179.0 [114.0–439.0] vs. 64.5 [32.3–94.5], p < 0.01) (Fig. 3C).
Meanwhile, the numbers of Th1 (CD4+Tbet+) (active vs. stable, 37.0 [22.0–93.0] vs. 1.0 [0.0–4.0], p < 0.01; active vs. control, 37.0 [22.0–93.0] vs. 0.0 [0.0–0.0], p < 0.001) and Th17 cells (CD4+RORγt+) (active vs. stable, 42.0 [23.0–155.0] vs. 6.5 [2.5–17.8], p < 0.01; active vs. control, 42.0 [23.0–155.0] vs. 0.0 [0.0–5.0], p < 0.001) were significantly elevated in the active lacrimal microenvironment compared with those in stable and normal ones. No differences in Th1 and Th17 cells were found between the stable and normal lacrimal microenvironment (Fig. 3C). There was the highest number of Th2 cells (CD4+GATA3+) (stable vs. active, 18.0 [7.3–55.3] vs. 2.0 [0.0–6.0], p = 0.019; stable vs. control, 18.0 [7.3–55.3] vs. 0.0 [0.0–0.0], p < 0.001) in the stable lacrimal microenvironment among the three groups (Fig. 3C).
In addition, there were more Treg cells (CD4+FOXP3+) (active vs. control, 6.0 [3.0–15.0] vs. 0.0 [0.0–0.0], p < 0.001; stable vs. control, 5.0 [1.8–19.8] vs. 0.0 [0.0–0.0], p < 0.001) both in active and stable lacrimal microenvironments than in a normal one (Fig. 3C), implying a possible endogenous protective role of immune regulation through the negative feedback loop of Treg cell activation.
We then accessed the frequencies of Th1 and Th17 cells in lacrimal glands by flow cytometry. Significantly increased proportions of CD3+CD8−IFN-γ-producing T (active vs. stable, 31.7 [21.3–41.1] vs. 3.8 [0.4–20.8], p = 0.015; active vs. control, 31.7 [21.3–41.1] vs. 0.1 [0.0–0.6], p < 0.001) and CD3+CD8−IL-17A-producing T (active vs. stable, 4.5 [2.9–6.1] vs. 0.7 [0.0–1.4], p < 0.01; active vs. control, 4.5 [2.9–6.1] vs. 0.0 [0.0–0.2], p < 0.001) cells were observed in active lacrimal glands compared with stable and normal ones (Fig. 3D, E), consistent with the TriMAP results.
Similarly, the proportions of CD3+CD8−Tbet-expressing T (active vs. control, 13.8 [11.3–18.1] vs. 1.0 [0.3–2.6], p < 0.001; stable vs. control, 7.4 [5.3–10.8] vs. 1.0 [0.3–2.6], p < 0.01) and CD3+CD8−RORγt-expressing T (active vs. control, 36.3 [24.7–53.1] vs. 1.9 [1.4–3.3], p < 0.001; stable vs. control, 17.5 [3.8–29.3] vs. 1.9 [1.4–3.3], p = 0.019) cells were significantly elevated in GO lacrimal glands compared with normal controls (Fig. 3D, E). The proportion of CD3+CD8−RORγt-expressing T cells (active vs. stable, 36.3 [24.7–53.1] vs. 17.5 [3.8–29.3], p = 0.032) was much higher in active lacrimal glands than stable ones (Fig. 3E).
Fibrosis was a hallmarked pathological change in GO lacrimal glands
Fibrosis, deposition of extracellular matrix (ECM), is an important pathological change of GO orbital connective tissues (20,21). We wondered whether fibrosis also occurred in GO lacrimal glands. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was performed to determine the biological and functional pathways of differentially expressed genes from our RNA-sequencing data. Genes involved in the ECM-receptor interaction pathway of KEGG database are demonstrated in Figure 4A. These ECM-related genes were highly enriched in active lacrimal glands compared with stable and normal controls (Fig. 4A). Since adipogenesis was another vital pathological change of GO orbital connective tissues (20), we examined genes involved in the antiperoxisome proliferator-activated receptor (PPAR) signaling pathway of KEGG database. Likewise, they were mainly elevated in active lacrimal glands (Fig. 4B).
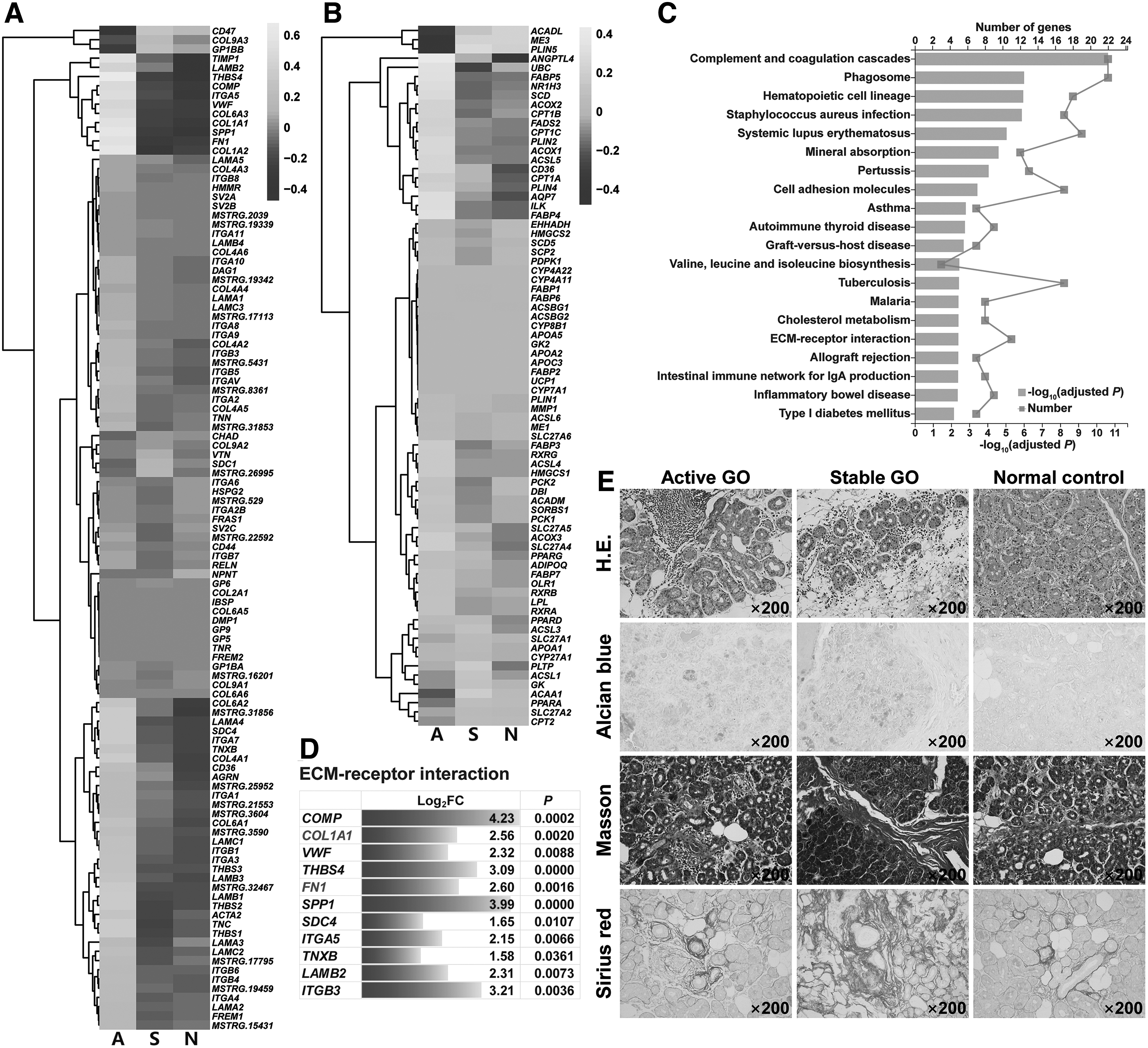
Fibrosis and adipogenesis in lacrimal glands from GO patients and control subjects. (
It is not surprising that the fibrotic and adipogenic transcriptome was higher in active stage than in stable stage, implying that there was a sequential process where the fibrotic and adipogenic genes were increased in active disease and gradually faded along with their protein deposition in stable disease marked by tissue remodeling.
Moreover, the ECM-receptor interaction pathway was significantly upregulated (adjusted p < 0.01) in active lacrimal glands compared with normal controls, which was among the top 20 significantly enriched KEGG pathways (Fig. 4C). Within the ECM-receptor interaction pathway, we found 11 differentially expressed genes that were significantly increased, including FN1 and COL1A1 (Fig. 4D). The PPAR signaling pathway was not significantly different in lacrimal glands between GO patients and normal controls.
We also explored lacrimal fibrosis and adipogenesis by the IHC study. Hematoxylin–eosin staining revealed that glandular parenchyma and stroma were infiltrated with a large number of mononuclear cells in active lacrimal glands, indicating robust inflammatory injury. Yet, stable lacrimal glands were destroyed with glandular atrophy, interstitial fibroplasia, and adipocyte proliferation, suggesting vanishing of the inflammation and enhanced tissue remodeling caused by fibrosis and adipogenesis (Fig. 4E). Furthermore, increased ECM deposition with enlarged collagen fiber bundles in GO lacrimal glands was displayed by Alcian blue, Masson, and Sirius red stainings, especially in stable stage (Fig. 4E).
IL-17A promoted fibrosis and adipogenesis while IFN-γ attenuated them in GO fibroblasts
Previous studies showed that circulating CD34+ fibrocytes appear to infiltrate GO orbital connective tissues and differentiate into CD34+ orbital fibroblasts (OFs) that are the main target cells in GO pathogenesis (22,23). Furthermore, CD34+ cells were observed in the fibrous septa of GO lacrimal glands compared with normal ones (Fig. 5A), suggesting that CD34+ fibrocytes might possibly infiltrate GO lacrimal glands as well.
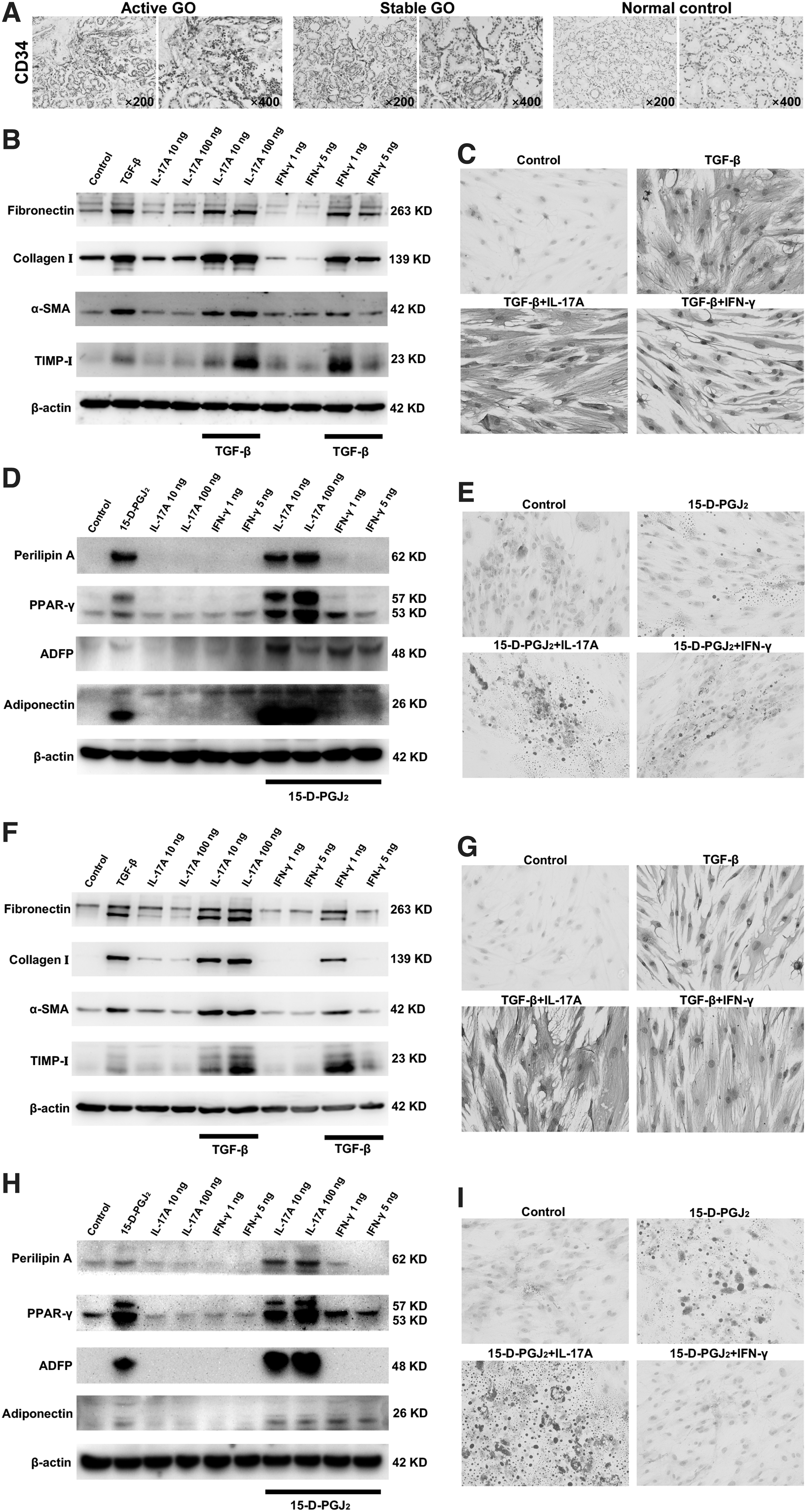
Effects of IL-17A and IFN-γ on CD34+ LFs and OFs derived from GO patients. (
Our data revealed that transforming growth factor (TGF)-β increased the expression of fibronectin, collagen I, α-smooth muscle actin, and tissue inhibitor of metalloproteinase-I in CD34+ lacrimal fibroblasts (LFs) and OFs (Fig. 5B, F). No obvious fibrotic changes were seen if only recombinant human (rh)IL-17A or rhIFN-γ was used to treat CD34+ OFs and LFs (Fig. 5B, F). Yet, IL-17A showed a profibrogenic effect in the presence of TGF-β, especially at a high concentration (100 ng/mL) (Fig. 5B, F). In contrast, IFN-γ exerted an inhibitory effect on fibrosis induced by TGF-β in CD34+ LFs and OFs (Fig. 5B, F). Our results were further confirmed by vimentin staining in which 100 ng/mL rhIL-17A promoted fibrosis while 5 ng/mL rhIFN-γ attenuated fibrosis in TGF-β-treated CD34+ LFs and OFs (Fig. 5C, G).
For adipogenesis induction of CD34+ LFs and OFs, the expression of perilipin A, PPAR-γ, antiadipose differentiation-related protein (ADFP), and adiponectin was increased by 15-deoxy-Δ12,14-prostaglandin J2 (15-D-PGJ2) treatment (Fig. 5D, H). Neither rhIL-17A nor rhIFN-γ upregulated those proteins, indicating that IL-17A or IFN-γ alone could not affect the adipogenic differentiation of CD34+ LFs and OFs (Fig. 5D, H). On the basis of 15-D-PGJ2, IL-17A enhanced the expression of perilipin A, PPAR-γ, ADFP, and adiponectin in CD34+ LFs and OFs, especially at a high concentration (100 ng/mL) (Fig. 5D, H). However, IFN-γ restrained adipogenesis induced by 15-D-PGJ2 in CD34+ LFs and OFs (Fig. 5D, H).
Likewise, more lipid droplets were observed by oil red O staining in CD34+ LFs and OFs in the presence of 15-D-PGJ2 under the stimulation of 100 ng/mL rhIL-17A. Yet, 5 ng/mL rhIFN-γ decreased lipid droplets in 15-D-PGJ2-treated CD34+ LFs and OFs (Fig. 5E, I). Collectively, our data suggested that IL-17A had both profibrogenic and proadipogenic effects on GO fibroblasts.
IL-17A activated fibrosis and adipogenesis-related pathways in GO fibroblasts
To explore the potential mechanisms of IL-17A on CD34+ LFs and OFs, we examined IL-17A-mediated intracellular signaling. The modified protein mass spectrometry of CD34+ LFs treated with 100 ng/mL rhIL-17A showed that JAK-STAT, mTOR, and PPAR-γ signaling pathways were activated (Fig. 6A). In CD34+ OFs, IL-17A activated JAK-STAT, mTOR, MAPK, adipocytokine, and insulin signaling pathways (Fig. 6B).
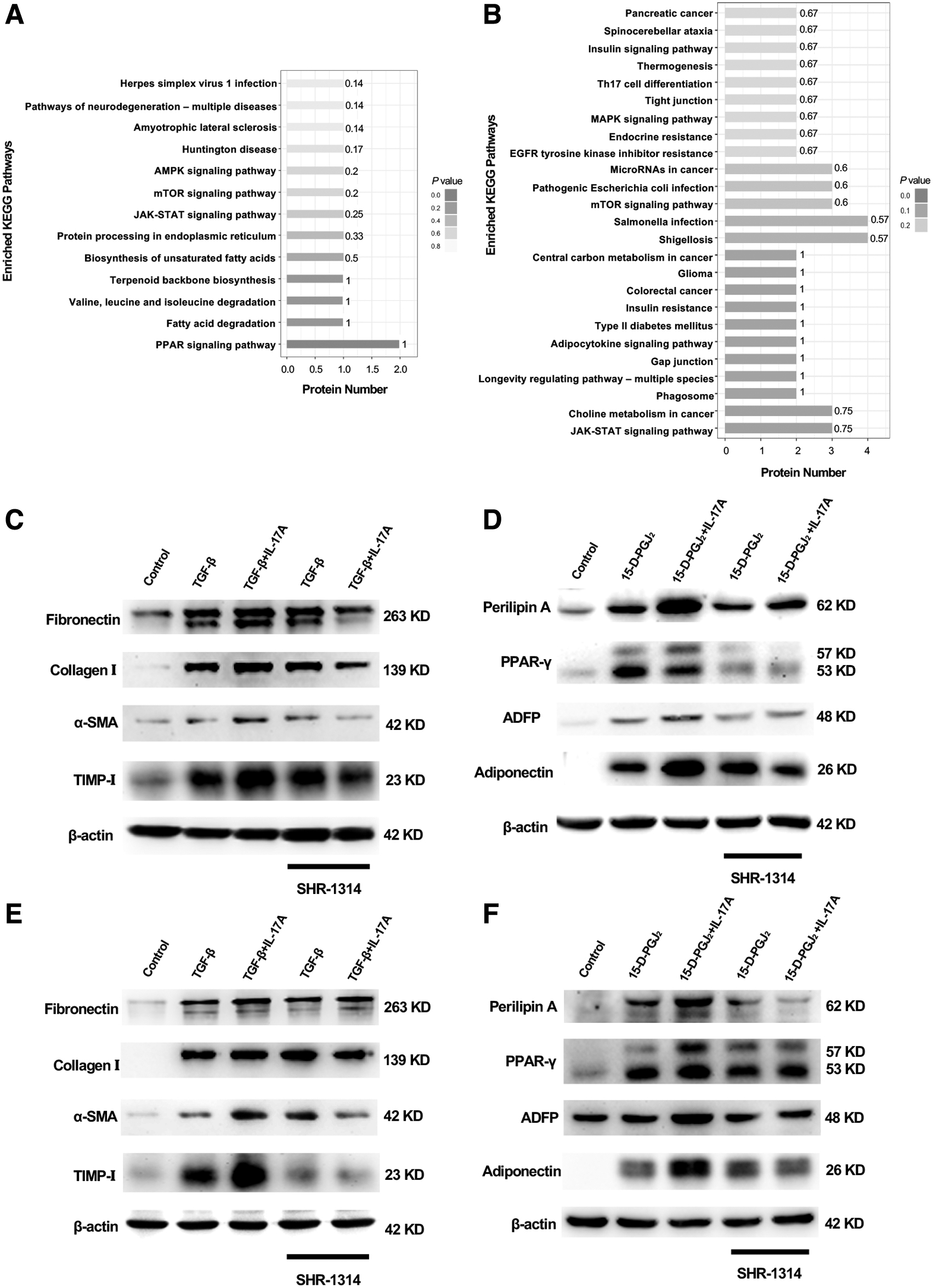
IL-17A signaling pathways in CD34+ LFs and OFs. (
The effect of SHR-1314, a novel monoclonal antibody of IL-17A
SHR-1314 (vunakizumab; Hengrui Pharma, China) is a humanized IgG1 anti-IL-17A monoclonal antibody composed of two identical heavy chains and two identical light chains. It can selectively bind to IL-17A and block its interaction with receptors.
We investigated its blocking effect in our in vitro GO models. One hundred nanograms per milliliter IL-17A was used as a profibrogenic or proadipogenic agent for TGF-β- or 15-D-PGJ2-activated CD34+ LFs and OFs, respectively. SHR-1314 did not affect TGF-β-induced fibrosis in CD34+ LFs and OFs (Fig. 6C, E, and Supplementary Fig. S1A, C), nor did SHR-1314 in 15-D-PGJ2-induced adipogenesis (Fig. 6D, F and Supplementary Fig. S1B, D). However, SHR-1314 stopped IL-17A-dependent enhancement of fibrogenic and adipogenic processes in CD34+ LFs and OFs (Fig. 6C–F). Taken together, our data warranted that the novel monoclonal antibody SHR-1314 could be a promising therapeutic agent for GO.
Discussion
The lacrimal glands are understudied in GO. Although xerostomia and ocular surface damage usually occur in active GO, many studies have confirmed that ocular surface inflammation and dry eye symptoms are quite common not only in the active phase but also in the stable phase (24). The lacrimal gland expressing thyrotropin receptors is histologically identical to the thyroid that is susceptible to immunological damage (2,5), and is likely to be involved in the same autoimmune response in active GO, causing reduced tear secretion and increased inflammatory factors in tear components (7 –11). Therefore, lacrimal glands are expected to be another target organ of GO autoimmune reactions, in addition to orbital connective tissues.
In this study, we demonstrated a complicated GO lacrimal immune microenvironment, mainly consisting of T cells, B cells, and monocytes, characterized by fibrosis and adipogenesis. The increased number of CD8+ T cells and monocytes in active lacrimal glands was in conformance with the observation of an early wave of CD8+ T cell and macrophage recruitment in orbital connective tissues from GO murine models during disease onset (25). It was previously reported that CD4+ Th cells were the most abundant orbital-infiltrating lymphocytes in GO (16,26). We found that the lacrimal CD4+ T cells produced higher levels of IFN-γ and IL-17A, especially in the active stage, indicating that both Th1 immunity and Th17 immunity were involved in GO lacrimal inflammation.
Our findings give a clue to the origin of elevated IL-17A levels in GO tears (8 –11). Although gene and protein expressions of IL-4 were not changed in GO lacrimal glands, distinct from orbital connective tissues, the number of GATA3-expressing T cells was significantly increased in stable lacrimal glands, implying a less important role of IL-4 or that other cytokines produced by GATA3+ Th2 cells may participate in the pathological process of stable lacrimal glands.
Given the fact that CD34+ OFs derived from fibrocytes are the main target cells in GO orbital connective tissues (23), one could speculate that those CD34+ fibroblasts may also play a pathogenic role in GO lacrimal glands. As expected, we observed increased generation of CD34+ cells in GO lacrimal glands. Our in vitro experiments further confirmed that CD34+ LFs differentiated into myofibroblasts or adipocytes, similar to CD34+ OFs, under IL-17A stimulation. This could explain lacrimal tissue remodeling formed by fibrosis and adipogenesis attributing to those CD34+ LFs. Fibrosis may be due to the activation of JAK-STAT pathway that can promote liver and left atrial fibrosis (27,28) and mTOR pathway that can promote liver and pulmonary fibrosis (29,30) in CD34+ LFs and OFs.
The MAPK pathway, which has been proven to mediate GO fibrosis (31,32), was activated in CD34+ OFs. Adipogenesis may lie in the activation of PPAR-γ pathway in CD34+ LFs, which plays an important role in adipocyte differentiation (33). Yet, in CD34+ OFs, the insulin signaling pathway was activated, which we previously reported as enhancing adipocyte differentiation of OFs (34). Altogether, IL-17A contributed to GO fibrosis and adipogenesis via numerous mechanisms, and further confirmatory research is needed.
Shih et al. reported fibrosis in GO Müller's muscles (35). We recently reported an elevated number of Th1 and Th17 cells in GO paranasal sinus mucosa (36). Hence, tissues adjacent to orbital fat may have the same damage mechanisms characterized by inflammation-mediated fibrosis and adipogenesis. More research is needed to determine whether IL-17A or other cytokines participate in those processes.
It is widely accepted that Th17 cells play an important role in human autoimmune diseases, which have been confirmed in disease-associated animal models and clinical practice (19,37). IL-17A blockade has been successful in treating psoriasis and ankylosing spondylitis (38,39). We previously reported that intravenous methylprednisolone therapy significantly reduced the proportion of circulating IL-17A-producing Th17 cells in active, very severe GO patients (40). Mu et al. demonstrated decreased serum IL-17A levels in active, moderate-to-severe GO patients receiving intravenous methylprednisolone either on a weekly or daily regimen at the 12-week visit (41).
However, Siomkajło et al. found no obvious changes of circulating Th17 cells in active, moderate-to-severe GO patients after intravenous methylprednisolone treatment on the same weekly scheme (42). Yet, Siomkajło et al. defined Th17 cells as CD161+RORγt+ cells and they did not examine the levels of IL-17A. When we defined Th17 cells as CXCR3−CCR6+ cells, they did not change after intravenous methylprednisolone treatment (40). Owing to the similar proinflammatory, profibrotic, and proadipogenic mechanisms of IL-17A in both the lacrimal glands and orbital connective tissues, targeting Th17 cells may have a therapeutic effect on both tissues.
SHR-1314 is a recombinant humanized monoclonal antibody against IL-17A. At present, 10 clinical trials at different stages have been launched using this drug (e.g., NCT03710681, NCT04840485, NCT05055934). In this study, SHR-1314 appeared to reverse the profibrotic and proadipogenic effects of IL-17A on CD34+ LFs and OFs, suggesting a potential future therapeutic benefit.
Our study is subject to several limitations. First, although patients were consecutively recruited, not all of the surgical explants from patients and controls were subject to all of the testing and some of the experiments were performed on small numbers of specimens, and so, there is potential for selection bias. Some barriers to inclusion of all of the samples in all of the analyses included the following: insufficient available tissue volume and the experimental cost. Since the experimental conditions of basic research are relatively controllable, the sample size in each study group of in vitro experiments usually depends on the experimental type and its empirical evidence, with special consideration to the biological repetitions. Different sample sizes of patients and controls were applied for different biological experiments in our study, but the risk of the potential for selection bias still exists.
Second, our study design is exploratory, as we did not have a clear hypothesis and we did not identify an a priori primary outcome. Thus, confirmatory research is needed. Third, the gating strategies of Th17 cells may have influenced the results, which may partially explain the differences in our findings relative to previous studies (40 –42). Since IL-17A may induce fibrosis and adipogenesis both in CD34+ OFs and LFs, it would have been preferable and more accurate to focus on the cytokine instead of cell surface marker or transcriptional factor.
In conclusion, our study reveals key features of GO lacrimal immunophenotype in which the occurrence and development of inflammation and tissue remodeling partially depend on IL-17A and IFN-γ axes. Our study suggests T cell-based GO pathogenesis, which will help to inform future research in new therapies for GO.
Footnotes
Authors' Contributions
Research idea: S.F. and H.Z.
Experimental design: S.F., H.Z., and J.S.
Sample collection: S.Z., Y.W., Y.L., and J.S.
Performance of experiments: Y.H., Y.W., S.Z., Y.L., and Y.W.
Data analysis: Y.H., Y.W., and X.L.
Writing and revising: Y.H., Y.W., S.Z., S.F., and H.Z.
Acknowledgment
We thank Mr. Haotian Yu (Majorbio Bio-Pharm Technology) for his help with RNA-sequencing data analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Our study was supported by the National Natural Science Foundation of China (81930024, 81761168037, 81770974, 81800695, 82071003, and 82000879), the National Key R&D Program of China (2018YFC1106100 and 2018YFC1106101), the Research Grant of the Shanghai Science and Technology Committee (20DZ2270800, 17DZ2260100, 19410761100, and 19DZ2331400), the Clinical Research Plan of SHDC (SHDC2020CR3051B), and the Project of Medical Robots (IMR-NPH202002). From the Clinical Joint Research Center of the Institute of Medical Robots, the Shanghai Municipal Education Commission—Gaofeng Clinical Medicine Grant Support (20152228), the Shanghai JiaoTong University Translational Medicine Crossed Research Grant (ZH2018ZDA12, ZH2018QNA07), the Sample Database Project of Shanghai Ninth People's Hospital (YBKB201901), and the Joint Innovation Team for Young Physicians of Shanghai Ninth People's Hospital (QC202002). Innovative Research Team of High-Level Local Universities in Shanghai (SHSMU-ZDCX20210901, SHSMU-ZDCX20210601).
Supplementary Material
Supplementary Materials
Supplementary Figure S1