
Abstract
Background:
The gut is a target organ of thyroid hormone (TH) that exerts its action via the nuclear thyroid hormone receptor α1 (TRα1) expressed in intestinal epithelial cells. THs are partially metabolized via hepatic sulfation and glucuronidation, resulting in the production of conjugated iodothyronines. Gut microbiota play an important role in peripheral TH metabolism as they produce and secrete enzymes with deconjugation activity (β-glucuronidase and sulfatase), via which TH can re-enter the enterohepatic circulation.
Summary:
Intestinal epithelium homeostasis (the finely tuned balance between cell proliferation and differentiation) is controlled by the crosstalk between triiodothyronine and TRα1 and the presence of specific TH transporters and TH-activating and -inactivating enzymes. Patients and experimental murine models with a dominant-negative mutation in the TRα exhibit gross abnormalities in the morphology of the intestinal epithelium and suffer from severe symptoms of a dysfunctional gastrointestinal tract. Over the past decade, gut microbiota has been identified as an essential factor in health and disease, depending on its compositional and functional profile. This has led to a renewed interest in the so-called gut-thyroid axis. Disruption of gut microbial homeostasis (dysbiosis) is associated with autoimmune thyroid disease (AITD), including Hashimoto's thyroiditis, Graves' disease, and Graves' orbitopathy. These studies reviewed here provide new insights into the gut microbiota roles in thyroid disease pathogenesis and may be an initial step toward microbiota-based therapies in AITD. However, it should be noted that cause-effect mechanisms remain to be proven, for which prospective cohort studies, randomized clinical trials, and experimental studies are needed.
Conclusion:
This review aims at providing a comprehensive insight into the interplay between TH metabolism and gut homeostasis.
Introduction
It has now been established that there is essential crosstalk between thyroid hormones (THs) and the gut. TH controls the intestinal epithelium homeostasis via the interaction between triiodothyronine (T3 or L-3,5,3′-triiodothyronine) and thyroid hormone receptor α1 (TRα1), the dominant TR isoform expressed in intestinal epithelial cells, the presence of specific TH transporters, and TH-activating or inactivating enzymes. In turn, the gut microbiota, consisting of 1013 to 1014 bacterial cells, has been identified as an important regulator of health and disease, including autoimmune thyroid diseases (AITDs). This review addresses the different aspects of this so-called gut-thyroid axis: the role of THs in intestinal epithelium homeostasis, the important function of the gut microbiota in peripheral TH metabolism, and the interaction between these gut microbiota and thyroid autoimmunity.
Intracellular TH Metabolism
THs are tyrosine-based molecules that contain iodine atoms at three or four positions of the aromatic rings. The two principal THs are thyroxine (also known as T4 or L-3,5,3′,5′-tetraiodothyronine) and T3. The THs are produced and secreted by the thyroid gland under strict control of the hypothalamus-pituitary-thyroid-axis via a negative feedback loop. 1,2 The thyroid mainly produces T4, the prohormone, and to a lesser extent T3, the active hormone.
T4 requires conversion into T3 in the peripheral tissues to become biologically active as T3 is the only form that binds the TR. The conversion of T4 occurs at the cellular level by specific enzymes, the so-called deiodinases (DIOs), that can remove an iodine atom from the inner or outer ring of TH. Before TH can be metabolized, it must be transported into the cell by active TH transporters. Several TH transporter families have been described; monocarboxylate transporters (MCT8 and MCT10), the organic anion transporter polypeptides (OATP), large neutral amino acid transporters (LAT), and recently, SLC17A4 of the solute carrier family.
The expression of the different TH transporters is cell-specific, and the affinity of these transporters for other TH metabolites also differs; for example, MCT8 prefers T4 while MCT10 prefers T3. 2 –5
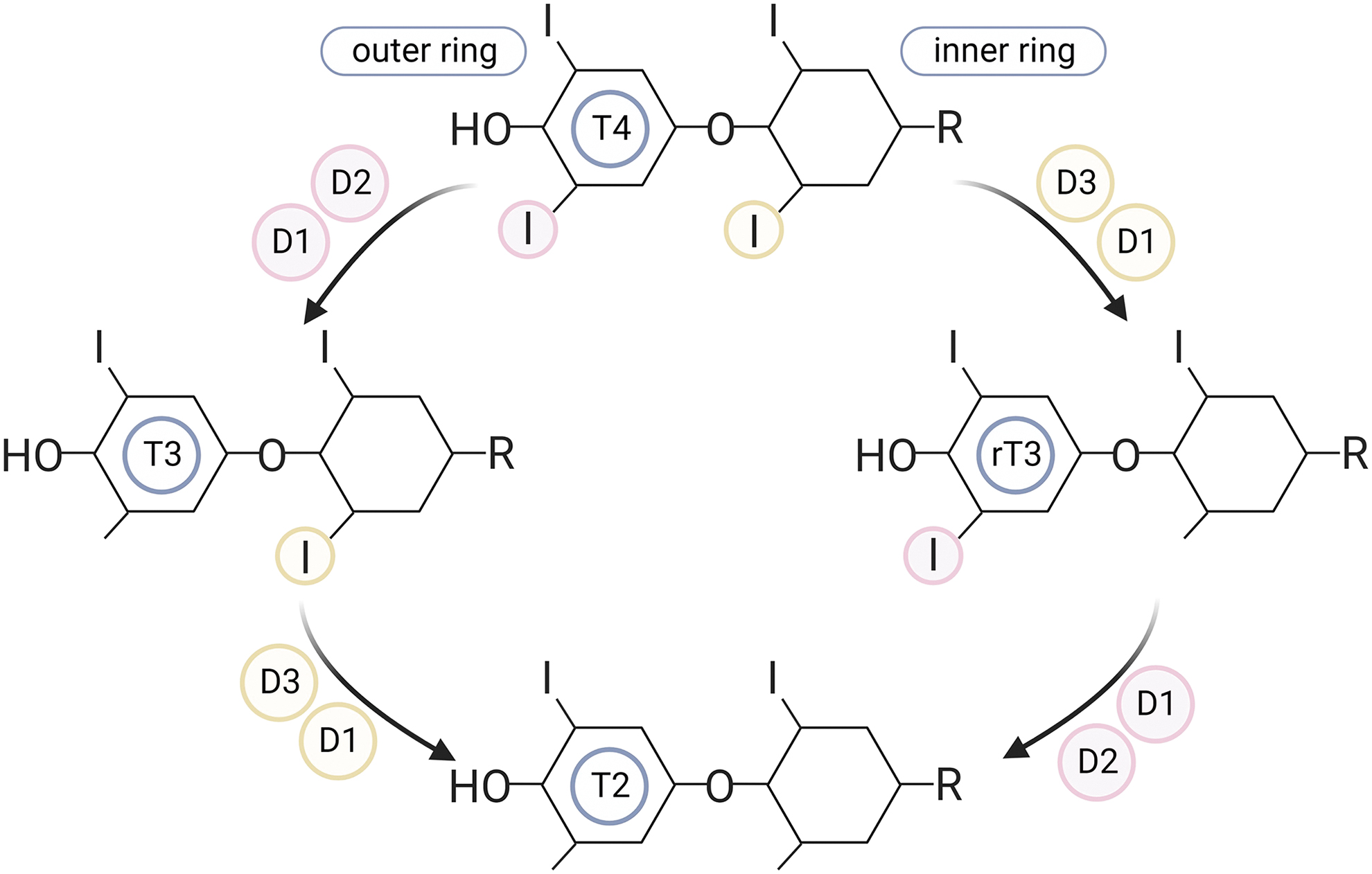
Three types of DIOs can be distinguished: type 1 deiodinase (DIO1), type 2 deiodinase (DIO2), and type 3 deiodinase (DIO3) (Fig. 1). DIO1 is capable of both inner and outer ring deiodination and is highly expressed in the liver, where it is believed to be the main source of local T3 and is important for the clearance of rT3. 6 DIO2 selectively removes an iodine atom of the outer ring of TH, resulting in the production of T3 out of T4. DIO2 is important in generating local T3 in specific tissues and cells and plays a significant role in negative feedback regulation as it is expressed in the hypothalamus and pituitary. 7
Different types of deiodinase enzymes.
DIO3 removes an iodine atom of the inner ring, thereby converting T4 and T3 to their respective inactive metabolites rT3 and diiodothyronine (T2). 5,8 DIOs are differentially expressed between various cell and tissue types. 5 The interplay between the transporters and DIOs determines the local TH availability in cells and tissues.
T3, the active hormone, exerts its action via binding to the TR, a nuclear receptor expressed in a wide variety of cell types. Several TR isoforms have been described, but only three isoforms can bind T3: TRα1, which is widely expressed in cardiac and skeletal muscle, the central nervous system, hematopoietic cells, bone, and intestine; TRβ1, which is mainly present in the brain, liver, and kidney; and TRβ2, which is expressed in the retina, inner ear, hypothalamus, and pituitary and thereby involved in negative feedback regulation. 1 These TR isoforms are encoded by the THRA and THRB genes, respectively.
The TRs act as transcription factors and are primarily necessary to establish the actual effect of T3 either with or without (in)direct DNA binding (canonical vs. non-canonical pathways) and can be classified into four types of TH signaling pathways. 9 In the canonical pathway, activated TRs behave as ligand-dependent transcription factors. They directly bind to specific DNA sequences named TH response elements, after which they promote the expression of the targeted genes.
In the non-canonical pathways, TRs can bind indirectly to DNA (tethered to DNA by other proteins) or participate in signaling pathways without the requirement of DNA binding and rapidly mediate second messenger signaling (by participating in the PI3K pathway, among others). Lastly, TH can exert its effect independently of TR through binding to integrin αvβ3. Integrin αvβ3 is a transmembrane receptor for T4 and to a lesser extent T3 and has been shown to result in rapid non-genomic signaling pathways, including extracellular signal-regulated kinases, with direct cellular responses. 10,11
Other TH-Metabolizing Pathways
Two major classical pathways of TH metabolism are sulfation and glucuronidation of iodothyronines. About 20% of daily T4 production appears in feces, predominantly via biliary excretion of conjugated iodothyronines. 12 Conjugation of iodothyronines occurs predominantly in the liver, leading to increased water solubility of the substrates, thus enhancing their biliary and urinary clearance. Sulfation of iodothyronines is performed by sulfotransferases that catalyze the sulfate conjugation of the phenolic hydroxyl group, resulting in the sulfated iodothyronines substrates T4S, T3S, rT3S, and 3,3′T2S.
This sulfation is a primary step toward rapid and irreversible inactivation of THs (T4 and T3), as these sulfated iodothyronines are highly efficient substrates of DIO1 (and are not processed by DIO2 and DIO3). 13,14 The rapid clearance of sulfated iodothyronines by DIO1 explains the low serum, bile, and urine levels of sulfated TH in adults. However, under conditions with low DIO1 activity (i.e., hypothyroidism, selenium deficiency, and non-thyroidal illness), the inactivation of TH by sulfation is reversible due to the expression of sulfatases in different tissues 15,16 and by gut microbiota, 17 which converts T3S back to T3.
Hepatic UDP-glucuronyltransferases are responsible for catalyzing the glucuronidation of the phenolic hydroxyl group of iodothyronines, as glucuronic acid is the first step in the enterohepatic cycle, resulting in the production of T4G, T3G, and T2G. 2,18 In contrast to the sulfates, glucuronidated iodothyronines are rapidly eliminated via biliary excretion in the intestine. After the biliary excretion of the glucuronidated iodothyronines (mainly T4G in humans) in the intestine, gut microbiota, specifically obligatory anaerobes, can hydrolyze T3G and T4G back to T3 and T4 by using the bacterial enzyme β-glucuronidase. 2,18 This process promotes the intestinal reabsorption of free iodothyronines into the enterohepatic circulation, where they are again available to the liver. These findings suggest that glucuronidated iodothyronine may serve as an intestinal TH reservoir, thereby preventing fluctuation of serum TH levels.
A small fraction of TH is metabolized by oxidative deamination or decarboxylation of the alanine side chain of TH. Deamination of T4 and T3 produces 3,3′,5,5′-tetraiodothyroacetic acid and 3,3′,5-triiodothyroacetic acid, respectively. Decarboxylation followed by deiodination results in the formation of thyronamines, including 3-T1AM 2 . Interestingly, the entire molecular machinery required for this T1AM biosynthesis is expressed in the (murine) intestinal tissue. 19
Another minor TH metabolizing pathway (<5% of TH disposal) is ether-link cleavage, resulting in the formation of diiodotyrosine. 20 For more information on these forms of TH metabolism, the reader is referred to a review by Wu et al, discussing these alternate pathways of TH metabolism. 20
The Role of TH in the Gut
The interplay of TH transporters, DIO enzymes, and TRs is of great importance for the bioavailability of TH in cells and tissues and, therefore, for the effect of TH. The integrated action of these TH signaling cascade components is cell type-specific, meaning that circulating TH concentrations do not necessarily reflect intracellular TH bioavailability. 21 The solute carrier family (SLC17A4) transporters are the predominant transporters in small intestinal and colonic epithelial cells, followed by LAT- and OATP-transporters. 3 All DIO enzymes are present in the gastrointestinal tract, with expression rates depending on different stages of embryonic and adult life. 22 –24 Both TRα and TRβ1 isoforms are present in the gastrointestinal tract, although TRα1 is the predominant subtype in the intestinal tissue to which T3 can bind. 25
The TH is involved in several processes in the gut. The intestinal epithelium consists of rapid and continuous cell renewal and is a well-established TH target. 26 Epithelial lineages derived from intestinal stem cells consist of rapidly proliferating progenitor cells in the crypts (invaginations of the intestinal wall) in both the small intestine and colon. The small intestine consists of differentiated cells in the villi (extensions of the intestinal wall), followed by apoptosis in the apex of the villi, whereas the colon consists of a flat surface with highly differentiated cell types in the upper part (Fig. 2).
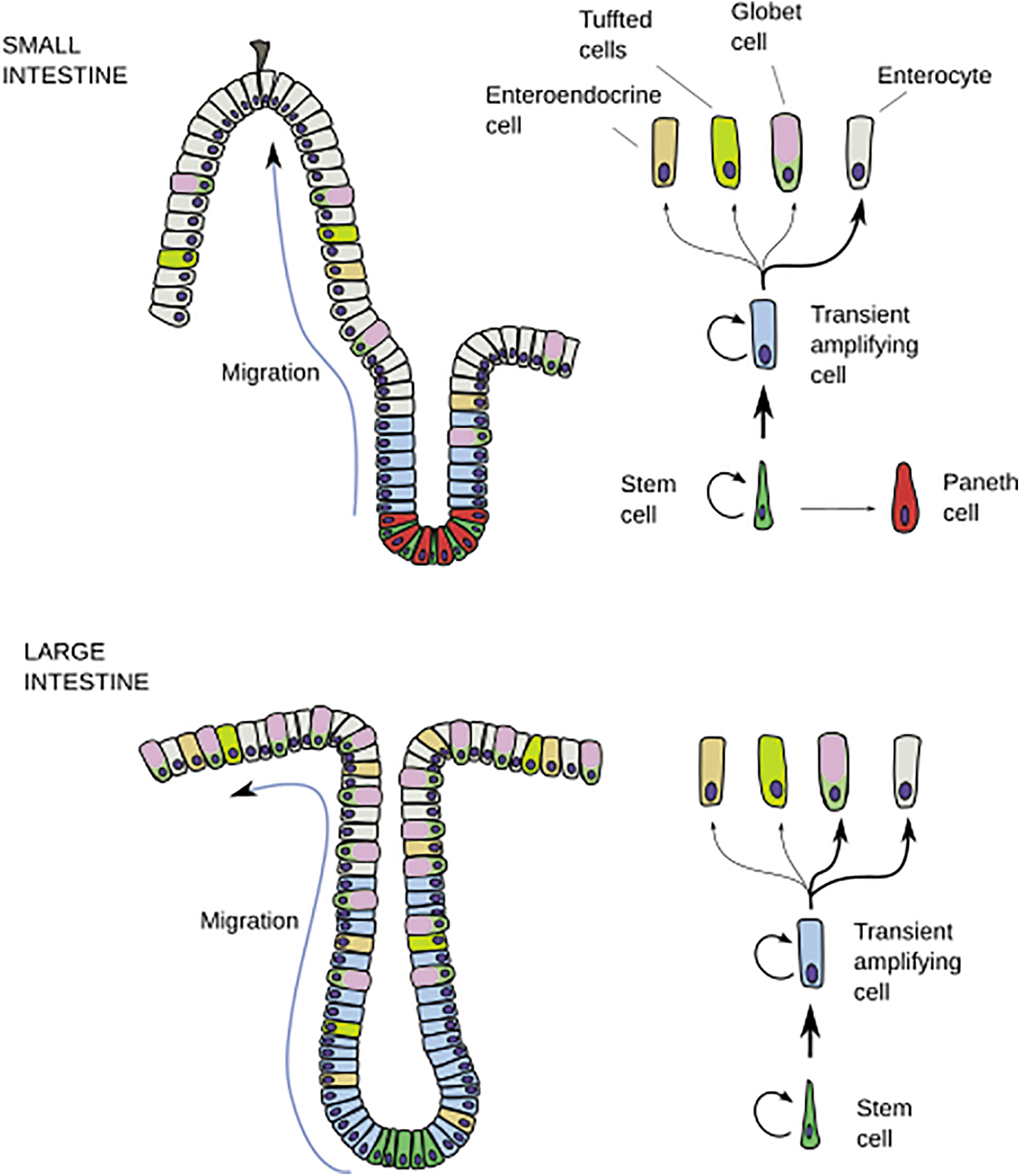
Drawing of the main cell lineages of the small intestine (upper part) and large intestine (bottom part). Thick arrows indicate a larger population. (Reproduced with permission from
Several studies have shown that the intestinal epithelium's developmental and physiological functions are controlled by TH signaling. 26 –28 The first evidence that supported this hypothesis derived from the observation that intestinal epithelium remodeling in amphibians completely depends on TH. 29 The observation that Dio3 is highly expressed in the intestinal epithelium of the human fetus fits within the idea that the regulation of intracellular TH concentrations is highly relevant during embryogenesis. 22
Interestingly, Dio3 is re-expressed in intestinal epithelium in adult life upon neoplastic transformation (intestinal adenomas and carcinomas) compared with healthy intestinal tissue, 24,30 indicating an essential role for TH in the homeostasis of intestinal cell proliferation and differentiation. It must be noted that TH and TRα1 are also coordinated and integrated with other signaling pathways in both mammalian and amphibian intestine. 31,32 As such, TH directly affects the Wnt/β-catenin pathway and Notch signaling, regulators of self-renewal and differentiation of stem/progenitor cells. 26,33
Intestinal alkaline phosphatase (IAP) is a brush border enzyme secreted by intestinal epithelial cells, of which the highest expression is located in the duodenum. 34,35 The primary function of IAP consists of a gut mucosal defense factor. It dephosphorylates the proinflammatory bacterial endotoxin lipopolysaccharide (LPS), thereby preventing the translocation of LPS into the systemic circulation and subsequent TLR4 responses of host cells. 36 The IAP gene is a T3-responsive gene, as shown by an increased IAP gene transcription in enterocytes in response to T3. 37 –40
Diminished IAP levels were seen in hypothyroid rats 41 displaying marked hypoplasia of crypts and villi. Interestingly, fecal samples of IAP-knock out (C56BL/6) mice showed a significantly reduced diversity of gut microbiota composition compared with their wild-type counterparts, 39 which was restored after oral supplementation of IAP. The IAP-deficient zebrafish were highly susceptible to LPS toxicity, resulting in a steep influx of intestinal neutrophils, whereas the intestines of germ-free zebrafish lacked neutrophils. 42
Overall, these studies indicate that THs are not only an important regulator of intestinal development but also promote mucosal tolerance to the commensal gut bacteria and may preserve gut microbiota composition via their action on IAP. 42
In this regard, the availability of transgenic mouse models made it possible to study the role of TH in the gut in more detail. TRα-/- mice (in which the TRα isoforms are abolished but the TRΔα isoforms remain) showed postnatal growth arrest with delayed maturation of the small intestine and bones. The intestine was the most affected organ in TRα0/0 mice (lacking TRα1, TRα2, and the shorter TRΔα transcripts). 43 These mice showed a reduced mucosal thickness compared with wild-type mice and a reduction of villus height, as well as decreased levels of digestive enzymes (lactase, sucrose, aminopeptidase) and gut transcription factors (Cdx-1 and Cdx-2) in the small intestine, indicating a diminished intestinal function. In contrast, no intestinal development retardation was seen in mice lacking the TRβ isoform (TRβ-/- mice), demonstrating that the proliferation of intestinal cells (crypts) is enhanced by T3 in a TRα1-dependent manner. 43
Moreover, TRα0/0 mice have an altered TH metabolism, indicated by a lower serum rT3/T4 ratio compared with their wild-type counterparts (2.63 ± 0.18 vs. 5.09 ± 0.22, p < 0.001, respectively). 44 These findings align with results from ex vivo experiments with 3D intestinal epithelium organoids, which showed a reduced development and stem cell activity in organoids prepared from crypt cultures of TRα0/0 mice compared with crypt cultures from either wild-type or TRβ-/- mice. In addition, accelerated stem cell proliferation and unbalanced differentiation were seen on T3 treatment in wild-type 3D intestinal organoids compared with non-treated control organoids. 45
Thus, T3 and TRα control gut development and homeostasis through the modulation of intestinal crypt cell proliferation and stem cell activity. 45 Specifically, they regulate the rate of cell renewal in normal conditions and of apoptosis and cell renewal during the process of epithelial regeneration in response to DNA damage. 26,33
The dominant role of TRα in intestinal epithelium homeostasis was also seen in the Thra1PV/+ mouse model (transgenic mice with a dominant-negative mutation in TRα that prevents binding of T3 to the TR, thereby repressing T3 positive receptor function), in which severe constipation was observed. 46 The murine intestine showed severe defects, consisting of shorter villi, increased differentiated cells in the crypts, and reduced stem-cell proliferation in the intestine compared with age-matched wild-type littermates. 46
Patients with a dominant-negative mutation in the TRα (so-called resistance to TH alpha [RTHα]) were first described a decade ago. 47 Since then, more than 40 patients with RTHα have been described with ∼25 variants in the THRA gene. 48 The observed mutations, comparable to the situation in the Thra1PV/+ mouse, result in a dominant-negative effect on the wild-type TRα, in which it fails to bind intracellular T3 and thereby actively represses T3 positive target gene transcription. 49 RTHα patients have only mildly affected circulating TH and thyrotropin (TSH) levels.
Of interest is that up to 84% of RTHα cases show symptoms of a dysfunctional gastrointestinal tract, mostly suffering from various degrees of constipation due to an increased intestinal transit time concomitant with bowel dilation. 48 Constipation is also one of the main characteristics in patients with primary hypothyroidism, defined by insufficient TH levels in the circulation. Whereas constipation usually ameliorates after initiating levothyroxine (LT4) treatment in hypothyroid patients, constipation in RTHα patients barely responds to LT4 supplementation, which might be caused by weakened smooth muscle contractility of the rectum in the latter patients. 50
To date, no data have been reported on the clinical features of human RTHα intestines. Animal studies observed abnormal morphology of the intestinal epithelium of RTHα mice characterized by shortened villi, increased differentiation in crypt cells, and decreased stem cell proliferation. 48 This is phenotypically different from RTHβ mice, in which the gastrointestinal tract is not affected. This can be explained by the fact that the TRβ1 is restricted to the differentiated epithelial cells of the villi, and no overt function for this protein has been described in the intestine. 51
In summary, intestinal epithelium homeostasis is controlled by T3. This process is regulated by the crosstalk between T3 and TRα1 and the presence of specific TH transporters and TH-activating and -inactivating DIO enzymes. However, it must be noted that these findings were primarily obtained from studies with genetically modified mice or experimental in vitro models using colorectal cell lines. It may, therefore, be difficult to translate directly to humans, although phenotypes observed in human RTHα patients do support a crucial role for TRα-mediated T3 signaling in intestinal function.
Gut Microbiome and TH Metabolism
The human gut microbiome (the collective genomic content of microorganisms) consists of 1013 to 1014 bacterial cells (microbiota) and has been identified as an important factor in various processes that impact host health and the occurrence and progression of disease. 36,52,53 The gut microbiota produce several gut-derived microbial metabolites, which act as signaling molecules allowing the gut microbes to exert their effect within the host. These microbial metabolites derive directly from bacteria or from the transformation (fermentation) of indigestible dietary components by the gut microbiota and include short-chain fatty acids (SCFAs) such as butyrate, acetate, and propionate, trimethylamine N-oxide, and branched-chained amino acids of which valine, isoleucine, and leucine are the most abundant.
These metabolites regulate immune responses 54 and maintain (intestinal) homeostasis. Changes in the abundance of these compounds are associated with several metabolic disorders, including metabolic syndrome, obesity, and type 2 diabetes. 55 –58 Other essential functions of the gut microbiota consist of the production and secretion of several vitamins (Vit K, folic acid, Vit B2, B3, B5, B6, B7, and B12) and the inducing of gut hormones secretion (leptin, ghrelin, GLP1). 59,60
It has been known for many years that the gut microbiota affect TH metabolism in the gut, which was first demonstrated by experimental rodent studies. 13,17,18,61 Several experimental studies have been performed with different administration routes of radiolabeled ([125I])-T3, -T3S, and -T3G in conventionally raised rats and rats without intestinal bacteria (using fecal suspensions from germ-free as well as from orally decontaminated rats).
It was shown that conventionally raised rats administrated [125I]-T3 intravenously excreted less radioactivity with feces and urine compared with decontaminated rats (feces 15.8% vs. 25.1% and urine 17.5% vs. 23.6%), 18 suggesting decreased enterohepatic reabsorption in the latter. The form of T3 and T3 conjugates excreted in the feces also differed between the two groups. Fecal samples of conventionally raised rats contain more T3 than samples from decontaminated rats (52.5% vs. 29.6%, respectively).
Moreover, no conjugates of T3 or 3-3′-T2 were detected in feces of conventionally raised rats, whereas only a small proportion of the glucuronidated iodothyronines were hydrolyzed in fecal samples of decontaminated rats, resulting in the excretion of substantial amounts of conjugated T3 (11.5% T3G and 10.9% T3S, respectively) in the feces of decontaminated rats (Fig. 3). 18
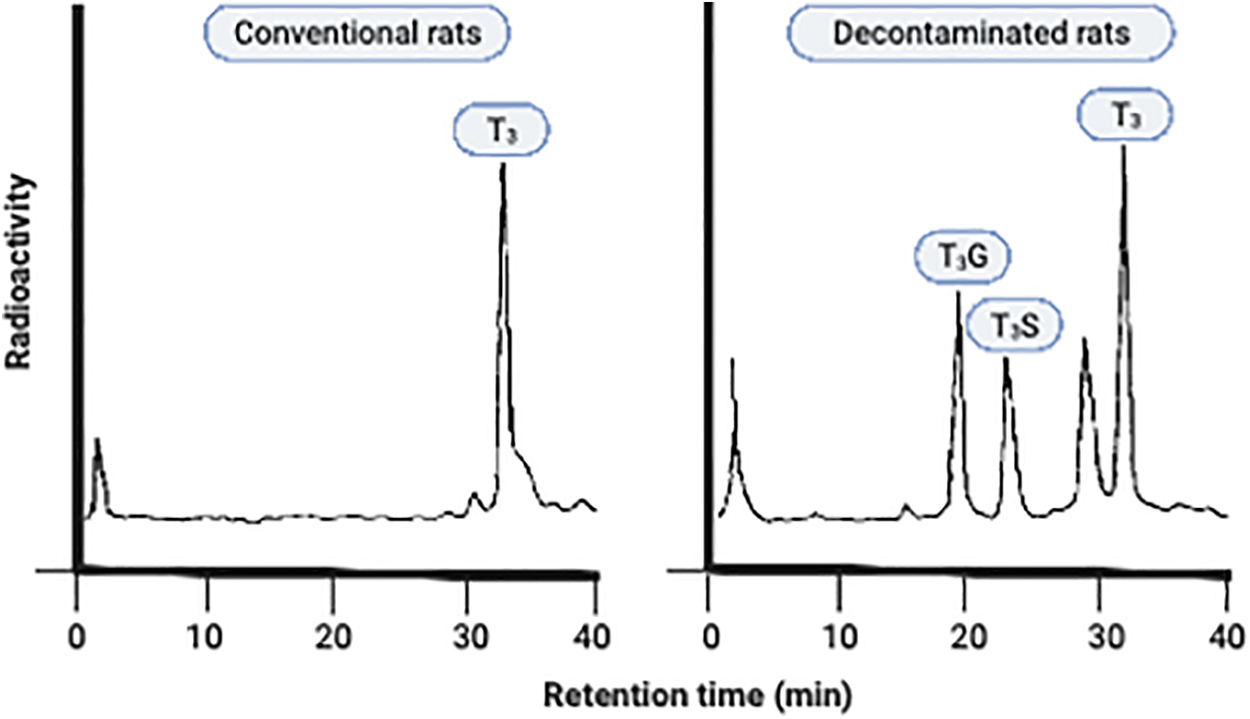
HPLC analysis of radioactivity from feces from conventional and decontaminated rats collected up to 38 hours after intravenous injection of [125I]T3. Adapted from de Herder et al. 18 Reprinted with permission. HPLC, high performance liquid chromatography.
In line with these findings, high performance liquid chromatography analysis of fecal samples of decontaminated rats receiving intragastric [125I]-T3G showed that radioactivity still existed in the form of T3G, whereas this was not the case in their conventionally raised counterparts. 18 Oral administration of [125I]-T3S or [125I]-T3G led to three to five times lower resorption of T3 in decontaminated rats compared with conventionally treated rats. Incubation of sulfated iodothyronines with feces from decontaminated rats resulted in no hydrolyzation of any of the iodothyronine sulfates. 17 In contrast, complete hydrolyzation of the various sulfate conjugates and T3G was observed when using fecal samples from conventionally raised rats, indicating that (anaerobic) gut microbiota possess sulfatase activity. 17
In conclusion, these studies demonstrate that hydrolysis of conjugated iodothyronines in the intestine is prevented by disturbing the gut microbiota composition, probably due to the lack of bacterial enzymes with deconjugation activity, β-glucuronidase and sulfatase. Of note, although β-glucuronidase-producing bacteria have been isolated from human feces, 61,62 most of these mechanistic findings are derived from animal models or in-vitro experiments and warrant validation in controlled human clinical trials. It remains unknown whether disruption of the gut microbiota composition, known as dysbiosis, affects the metabolic pathway of conjugated iodothyronines in humans.
Gut Microbiome and AITD
A potential effect of the gut microbiota composition on the thyroid gland was first hypothesized in the 1970s and was investigated using experimental rodent studies. The thyroid glands of rats exposed to antibiotics showed a significantly decreased uptake of radioactive iodine, indicating reduced thyroid functioning. 63 Studies with germ-free mice (lacking gut microbiota) revealed an increased TSH secretion of 25% compared with their conventionally raised counterparts. 64 Interestingly, specific pathogen-free rats were less susceptible to autoimmune thyroiditis than conventionally raised rats, suggesting that some gut microbiota might constitute a protective effect on the thyroid gland. 65
Further evidence to support this metabolic symbiosis between host and gut microbiota has only been demonstrated over the past decade. As most gut microbiota taxa are mainly obligate or facultative anaerobes, traditional culture techniques could not identify specific microbial species. 66 With the development of recent novel culture techniques combined with microbial gene sequencing (16s rRNA gene analysis, whole-genome shotgun sequencing, and metagenomics), a renewed interest has emerged in the human-microbiota interaction in health and disease. 36,67
As such, various studies have recently been published on the association between gut microbiota composition and AITD, such as Graves's disease (GD), Graves' orbitopathy (GO), and Hashimoto's thyroiditis (HT) (Tables 1 and 2) and have been recently reviewed by Virili et al. 68 By comparing AITD patients' fecal samples with healthy controls, these studies have shown significant differences in the diversity and composition of microbial strains between these two groups.
Characteristics of Studies that Investigated the Association Between Gut Microbiota and Hashimoto's Thyroiditis Disease
↑, increased; ↓, decreased; AITD, autoimmune thyroid disease; AUC, area under the curve; F/B ratio, Firmicutes/Bacteroidetes ratio; HT, Hashimoto's thyroiditis; LT4, levothyroxine; SMD, standardized mean differences; TPOAb, anti-thyroid peroxidase autoantibody.
Characteristics of Studies that Investigated the Association Between Gut Microbiota and Graves' Disease and Graves' Orbitopathy
GD, Graves' disease; GO, Graves' orbitopathy; LPS, lipopolysaccharide; SPF, specific pathogen-free; TRAb, thyrotropic receptor autoantibodies.
Several of these studies 69 –72 reported significant correlations between the relative abundance of the gut microbiota and diagnostic parameters of thyroid status (serum levels of TSH and free T4) and thyroid antibodies (TPOAb, TgAb, and TRAb), indicating the potential clinical significance of dysbiosis in these patients: The genera Bacteroides, Dorea, Faecalibacterium, and Coprococcus showed a significant inverse association with TPOAb or TRAb, whereas Blautia, Lactobacillus, Alistipes, Ruminococcaceae, and Enterobacteriaceae were positively correlated with the presence of TPOAb, which may be due to molecular mimicry from autoepitopes from these bacterial strains driving a T and B cell-mediated autoimmune reaction against thyroid gland tissue. 73
The productive function of the intestinal microbiota in the development of thyroid disease has also been studied 70,74 –77 using a random forest analysis to find discriminative microorganisms that could distinguish between patients and the control group. As such, hyperthyroid patients (area under the curve [AUC]-values are ranging between 0.76 and 0.98; Table 2) as well as hypothyroid patients (AUC-value of 0.92; Table 1), 74 could be identified with high accuracy based on their microbial composition. The identified microbiota (Tables 1 and 2) are of particular interest, as these might be useful for finding new biomarkers.
The earlier mentioned studies suggest the presence of a so-called gut-thyroid axis, representing a bidirectional signaling axis that regulates thyroid homeostasis. 78,79 However, the cross-talk pathways remain to be unraveled as these studies have yielded inconsistent and conflicting results in terms of diversity and specific microbiota identified. For example, one study 80 found a significantly higher diversity in GD patients, whereas others 81 –83 showed a reduced diversity among these patients. Similar conflicting results have also been found in studies with hypothyroid patients. 71,74,84 –87 This might be due to the fact that the studies have been conducted with relatively small heterogeneous patient groups (treated vs. untreated thyroid patients; Tables 1 and 2) with the inclusion of anti-TPO-negative patients, 74,86 different ethnicities 88 and using different techniques for microbial assessment (reverse transcription polymerase chain reaction, high-throughput sequencing of 16s rRNA and/or polymerase chain reaction and denaturing gradient gel electrophoresis).
Moreover, most of these studies lacked a functional assessment of the gut microbiota and did not report on the participants' dietary intake. Lastly, these studies were mainly conducted in China (Tables 1 and 2), which might impact the generalizability of the results to other regions of the world, as ethnicity and geography are significant factors in the variation of the gut microbiota composition. 89,90
Altogether, these data show an association between gut microbiota composition and (autoimmune) thyroid diseases but do not imply causality. Proving a causal relationship between dysbiosis and disease onset remains challenging, as the composition of the gut microbiota is influenced by many factors, including host delivery mode (vaginal delivery vs. C-section), use of medication (especially antibiotics), lifestyle and behavioral characteristics, genetics, and nutrition. 67
Fecal Microbiota Transplants in AITD
Humanized gnotobiotic mouse models are needed to unravel the underlying molecular mechanism and to better dissect the impact of healthy or dysbiotic microbiota on the progression or onset of AITD. Recent studies have investigated the effect of mice treated with fecal microbiota transplants (FMTs) with feces from healthy human individuals compared with FMTs from AITD patients (HT, 74 GD, 75 and GO, 91 respectively).
The FMTs from AITD patients led to an increase in disease incidence and severity. 74,75,91 concomitant with an increase in serum LPS level 74,75 and intestinal permeability 74 and a decrease in fecal SCFA concentrations. 74,75 Interestingly, mice treated with vancomycin showed a reduction in GO and GD incidence and severity accompanied by a lower microbiota diversity, whereas mice treated with FMT from GO patients inherited their GO donor's microbiota, leading to an increase in GO incidence. 91 This demonstrates a significant variation in gut microbiota composition in these murine models, correlating with GD heterogeneity. 91
However, various concerns should be addressed when translating gut microbiota research results from experimental murine models into humans, as there are notable differences in anatomy, genetics, and physiology. 90,92 Therefore, future research should include prospective studies assessing gut microbiota composition and functionality together with thyroid function, as well as randomized clinical trials that determine the effects of altering the gut microbiota composition on disease progression in AITD patients.
Conclusion
This review provides a comprehensive insight into the interplay between TH metabolism and gut homeostasis, the so-called gut-TH axis. The gut microbiota has been identified as an essential factor in health and disease, depending on its compositional and functional profile. It produces several gut-derived microbial metabolites, which act as signaling molecules, allowing the gut microbes to exert their effect within the host.
The TH is effectively metabolized via sulfation and glucuronidation (conjugation) of T3 and T4. Glucuronidated iodothyronines (T3G and T4G) are rapidly eliminated via biliary excretion into the intestine. Once excreted in the intestine, T3G and T4G can be hydrolyzed back to T3 and T4 by the gut microbiota, which can be reabsorbed into the enterohepatic circulation. This suggests that T3G and T4G serve as an intestinal TH reservoir.
The TH is involved in several processes in the gut. The homeostasis of the intestinal epithelium is controlled by T3 through its interactions with TRα1, the dominant TR isoform in the intestine. This homeostasis depends on tight regulation of local T3 concentrations, regulated by specific TH transporters and deiodination enzymes in the intestine. Patients and experimental murine models with a dominant-negative mutation in the TRα exhibit gross abnormalities in the morphology of the intestinal epithelium and suffer from severe symptoms of a dysfunctional gastrointestinal tract supporting the crucial role of TRα-mediated T3 signaling in intestinal function.
Disruption of gut microbial homeostasis (dysbiosis) is associated with AITD. However, a causal role of dysbiosis in AITD is yet to be established. Recent studies using experimental murine models suggest that FMTs can be a promising tool to treat AITD patients in the future. However, more research is needed to better understand the effects of altered gut microbiota composition on disease progression in AITD patients.
Footnotes
Authors' Contributions
A.C.F. and A.B. researched data for the article, made substantial contributions to the discussion of the content, wrote the article, and reviewed/edited the manuscript before submission. A.H.S. made substantial contributions to the content discussion and edited the manuscript before submission. M.N. and E.B. reviewed the manuscript before submission.
Author Disclosure Statement
M.N. is on the Scientific Advisory Board of Caelus Pharmaceuticals, the Netherlands. None of these are directly relevant to the current paper. There are no patents, products in development, or marketed products to declare. The other authors declare no competing financial interests.
Funding Information
A.F. is appointed on a LeDucq consortium grant 2017 17CVD01 (to M.N.). M.N. is funded by a personal ZONMW VICI grant 2020 [09150182010020].