
Abstract
Background:
Thyroid hormone (TH) has actions in every tissue of the body and is essential for normal development, as well as having important actions in the adult. The earliest markers of TH action that were identified and monitored clinically, even before TH could be measured in serum, included oxygen consumption, basal metabolic rate, serum cholesterol, and deep tendon reflex time. Cellular, rodent, amphibian, zebrafish, and human models have been used to study TH action.
Summary:
Early studies of the mechanism of TH action focused on saturable-specific triiodothyronine (T3) nuclear binding and direct actions of T3 that altered protein expression. Additional effects of TH were recognized on mitochondria, stimulation of ion transport, especially the sodium potassium ATPase, augmentation of adrenergic signaling, role as a neurotransmitter, and direct plasma membrane effects. The cloning of the thyroid hormone receptor (THR) genes in 1986 and report of the THR crystal structure in 1995 produced rapid progress in understanding the mechanism of TH nuclear action, as well as the development of modified THR ligands. These findings revealed nuances of TH signaling, including the role of nuclear receptor coactivators and corepressors, repression of positively stimulated genes by the unliganded receptor, THR isoform-specific actions of TRα (THRA) and TRβ (THRB), and THR binding DNA as a heterodimer with retinoid-x-receptor (RXR) for genes positively regulated by TH. The identification of genetic disorders of TH transport and signaling, especially Resistance to Thyroid Hormone (RTH) and monocarboxylate transporter 8 (Mct8) defects, has been highly informative with respect to the mechanism of TH action.
Conclusions:
The impact of THR isoform, post-translational modifications, receptor cofactors, DNA response element, and selective TH tissue uptake, on TH action, have clinical implications for diagnosing and treating thyroid disease. Additionally, these findings have led to the development of novel TH and TH analogue therapies for metabolic, neurological, and cardiovascular diseases.
Introduction
This invited review on the history of thyroid hormone (TH) action is one of a series of articles celebrating the American Thyroid Association centennial. Our understanding of TH action is the result of investigators working across multiple disciplines and experimental models, for at least 140 years. This review will focus on studies of the nuclear thyroid hormone receptor (THR), our understanding before the report of cloning of THR in 1986, and the impact of the cloning on our understanding of TH action and genetic disorders of TH action. Some of the clinical implications of insights into TH action will also be described. TH metabolism and local ligand availability, a key regulator of TH action, will be discussed in a separate review.
Early Observations of TH Action
The earliest attempts to measure TH action centered on the metabolic actions, assessing the increase or decrease in basal metabolic rate as a manifestation of TH status, body temperature, body weight, body fat composition, oxygen consumption, and the actions on circulating lipid levels, especially serum total and low-density lipoprotein cholesterol. 1 The recognition that oxygen consumption in patients with thyroid disease directly increased or decreased with the level of TH was reported as early as 1895 by Magnus-Levy, 20 years before thyroxine (T4) was isolated (reviewed in Magnus-Levy 2 ).
Additional actions of TH tracked clinically were the effects on the heart (chronotropic and inotropic), as well as deep tendon reflex relaxation time. Mitochondria were thought to be an essential site of TH action, based on TH stimulation of oxygen consumption in tissues. 3,4 In most tissues, except the brain, oxygen consumption increased or decreased with an increase or decrease in TH levels. Early studies and models recognized the importance of TH action in development, including the absolute requirement for TH to promote metamorphosis in frogs. 5 Interestingly, TH was recognized as essential for both of the key features of amphibian transition from aquatic to terrestrial life, tail resorption and limb generation.
The varied clinical manifestations of TH deficiency and TH excess, across almost every organ system in the body (Table 1), led many to consider that TH worked through a range of activating pathways. 6 –9 Although several TH targets were identified and studied, including mitochondria, ion transporters, and membrane receptors, the bulk of the action was recognized as mediated by nuclear THRs. TH is one of only two tyrosine-derived hormones, the other being catecholamines. Catecholamines circulate without being bound to proteins and act at membrane receptors, similar to the peptide hormone family. TH, which is hydrophobic, circulates bound to protein and would ultimately be shown to act through nuclear receptors, similar to steroid hormones. 10
Physiological and Biochemical Actions of Thyroid Hormone
Adapted from Tata. 13
Na+, sodium.
Models and Mechanisms of TH Action
The study of circulating serum proteins that bind T4 and triiodothyronine (T3) was an early focus that later informed studies of the TH nuclear receptor (reviewed in Pappa et al 11 ). It was found that there were individuals born with absent, or defective, TH binding proteins, thyroxine binding globulin, transthyretin (TTR), and albumin. Although a high fraction of circulating T4 and T3 is bound to proteins, the pituitary and hypothalamus regulate the “free fraction” of T4 and T3, such that the tissues are euthyroid despite serum total T4 and T3 concentrations outside the reference range in affected individuals. The characteristics of T3 and T4 binding revealed by TH binding protein mutations, however, were later used to guide investigation of nuclear TH binding. 11
A major milestone in studies of TH action was the demonstration by Tata in 1963 that inhibitors of RNA production and protein synthesis, actinomycin D and puromycin, blocked the biological actions of TH on growth and basal metabolic rate in a rodent model. 12 This challenged the popular view at that time of a direct action of TH on mitochondria and oxidative phosphorylation and favored an action that required protein synthesis. A series of follow-up studies by Tata, utilizing a range of animal, cellular, and amphibian models, including radioactive labeling techniques, directly demonstrated TH-induced nuclear protein synthesis, with a time course and pattern consistent with TH nuclear action (reviewed in Tata 13 ). Significant progress was made during this time by other laboratories, identifying the properties of the nuclear receptors for many steroid hormones, including estrogen, progesterone, and glucocorticoid.
These studies showed a mechanism of action with hydrophobic ligands passing into the cytoplasm, binding to receptors, the receptor–ligand complex entering the nucleus, binding specific DNA elements, and modifying gene transcription. 10 Specific nuclear T3 binding in growth hormone 1 pituitary cells was shown by Samuels in 1973. 14 A series of studies from a number of investigators in the mid-1970s, including Oppenheimer, Surks, Dillmann, Silva, Samuels, Refetoff, DeGroot, Tata, and Baxter, further characterized T3 nuclear binding in cells and tissues, linked T3 nuclear binding to gene expression of messenger RNA (mRNA) and protein, showed that T3, rather than T4, was the predominant bound ligand, and that tissues varied with respect to capacity for T3 binding, concentration highest in anterior pituitary, and then in descending order, in the liver, kidney, heart, and brain (reviewed in Sterling 3,4 and Oppenheimer 15 ).
Nuclear saturation by T3 was directly linked to the magnitude and duration of stimulation of a TH-responsive enzyme, α-glycerophosphate dehydrogenase, in the liver, which also correlated with oxygen consumption 15 (Fig. 1). Nuclear extracts were labeled with radioactive T3, and the T3 binding protein was isolated by gel electrophoresis, indicating a single species and a receptor size of about 60 kDa, similar to the size of other described steroid nuclear receptors.
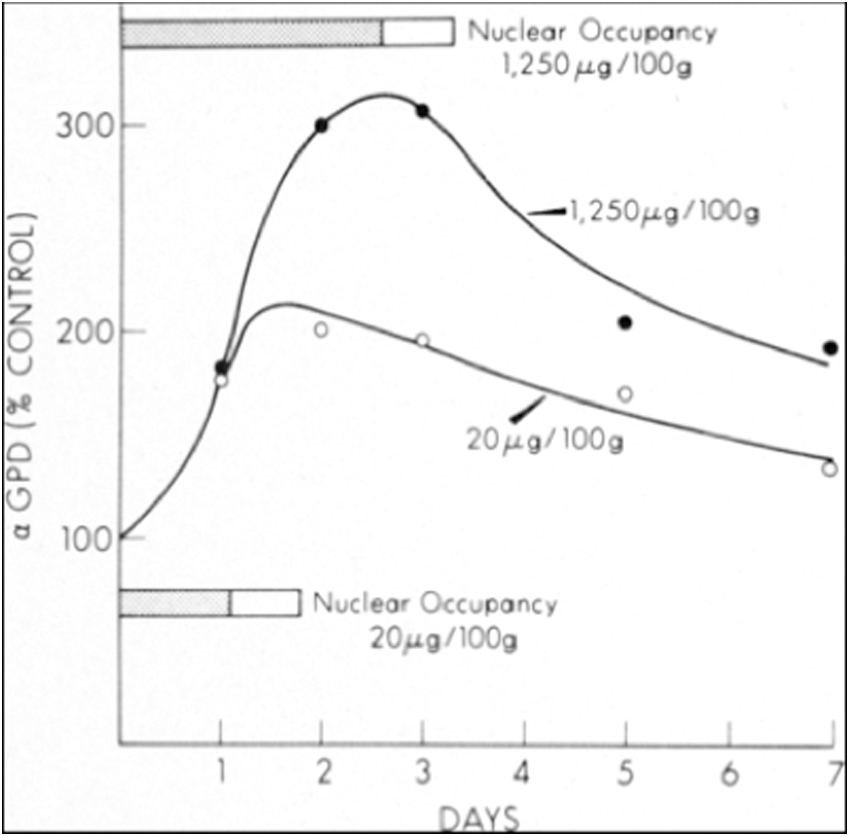
Effect of high and low doses of T3 on alpha-GPD activity and on nuclear occupancy by T3 stimulation of hepatic alpha-GPD activity in euthyroid rats given a single, low-dose or high-dose intravenous injection of T3. The peak induction of alpha-GPD was closely linked to the duration of T3 nuclear occupancy, shown in the horizontal shaded bars. Nuclear occupancy by T3 was closely linked to the magnitude and duration of enzyme stimulation. From Oppenheimer.15(p1065) Copyright © (1975) Massachusetts Medical Society. Reprinted with permission from Massachusetts Medical Society. GPD, glycerophosphate dehydrogenase; T3, triiodothyronine.
Studies of TH action, before the cloning of the nuclear THR genes, are described in reviews by Oppenheimer in 197515 and Sterling in 1979 3,4 (Table 2). The predominant mechanism of action was thought to be nuclear (Fig. 2A), but TH action was also recognized in mitochondria, adrenergic signaling, neural transmitters, and membrane actions. Some investigators focused almost exclusively on TH stimulation of thermogenesis by active sodium transport through upregulation of the sodium potassium ATPase (Na+-K+-ATPase). TH stimulation of oxygen consumption in the skeletal muscle was significantly reduced by treatment with ouabain, which inhibits the Na+-K+-ATPase. 16 Even those promoting a primary action of TH on ion transport, however, recognized that TH nuclear action on protein synthesis was the initial step, and these proteins stimulated by TH secondarily influenced ion transport.
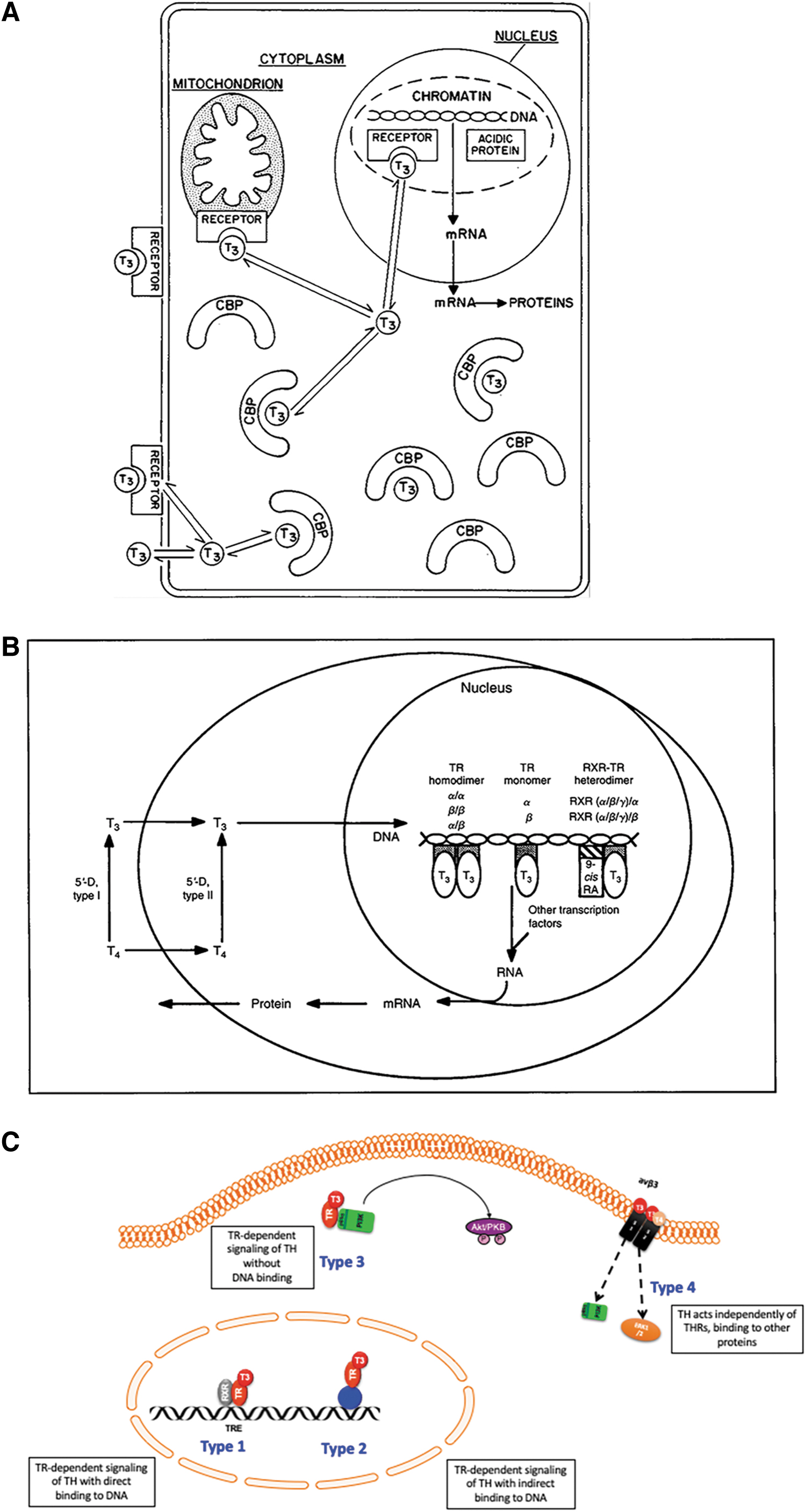
Models for TH action on the target cell 1979, 1994, and 2021.
Pathways of Thyroid Hormone Signaling from the 1970s
GPD, glycerophosphate dehydrogenase; Na+-K+-ATPase, sodium potassium ATPase; TH, thyroid hormone.
Cloning of the THR Genes
A major turning point in understanding TH action occurred in 1986, with publication of the cloning of the THR genes. 17,18 The work leading to this point represented the contribution of many laboratories and investigators using a wide range of models and tools to characterize T3 nuclear binding. Many of the key steps were the result of findings from studies of other steroid hormone signaling pathways. 10,19 Ultimately, however, TH had many unique features and actions that informed subsequent studies, especially in terms of DNA binding characteristics, nuclear receptor cofactors, and with respect to nutrient signaling and cross talk with other nuclear receptors regulating fat and carbohydrate metabolism. 1,6,7,9
The rapid progress in the field was also shaped by clinical/translational investigation, identifying individuals with defects in the thyroid signaling pathway. This was most notable for studies of Resistance to Thyroid Hormone (RTH), but also patients with defects of TH transport. 20,21 Molecular biology tools became available to clone genes of interest by a range of techniques, and multiple members of the steroid receptor family were cloned. Two groups, the laboratories of Evans and Vennström, working independently, published the cloning of the THR in the same issue of Nature. 17,18
The THR had similarities to an erythroblastic leukemia virus, v-erb-A, and so, the initial clone was referred to as cellular erb-A (c-erb-A). The structure of the THR was similar to those of other steroid nuclear receptors already cloned and included an N-terminal domain, DNA binding domain, and a carboxy-terminal ligand binding domain. Studies showed that these domains were “cassettes” that could be interchanged among the nuclear receptors and maintain their specificity of the nuclear receptor of origin. 19 A progesterone receptor DNA binding domain, for example, could be substituted for the estrogen receptor DNA binding domain, and the chimeric receptor would bind estrogen but recognize the progesterone DNA response element. 22 TR was later shown to have these same interchangeable components.
The first cloned steroid hormone receptors, estrogen, progesterone, androgen, and glucocorticoid, were predominantly in the cytoplasm bound to heat shock protein, which was displaced by ligand, entered the nuclear, and bound as homodimers to palindromic DNA response elements. 19,22 In contrast, THR and other “type 2” steroid and steroid-like receptors reside predominantly in the nucleus bound to DNA, form heterodimers with retinoid-x-receptor (RXR) that bind to direct repeat elements, with various spacing that gives specificity, bind to corepressor complexes that repress gene expression, displaced by ligand, coactivator complexes are recruited, and gene activation occurs. 23,24
Before full identification of the THR protein or cloning of the gene, it was possible to prepare nuclear protein extract and characterize DNA binding. Genes known to be directly stimulated by TH were limited, but it was known that growth hormone levels were very low in hypothyroid rodents. The rat growth hormone gene was shown to be stimulated by TH and glucocorticoids. 3,4 The earliest natural TH response element (TRE) was characterized using these methods and reported a sequence characterized as a “direct repeat” of 6 base pairs, with the sequence AGGTCA, and a 4 base pair “gap” 25,26 (Fig. 2B). Other TH-responsive genes were subsequently shown to have some variations in half site arrangement and sequence, but the direct repeat with two half sites separated by a 4 base pair gap, AGGTCANNNNAGGTCA, was considered the consensus site. 27
A similar DNA response element was identified that mediated the response to retinoic acid and TH. 28 This finding was an important divergence from the DNA response element that mediated binding of the other steroid receptors studied at that time, estrogen, progesterone, glucocorticoid, and mineralocorticoid. These receptors also bound to 6 base pair “half site” sequences, but these “half sites” were arranged as palindromes thought to facilitate homodimer binding, placing the common interface for covalent binding in direct proximity. A direct repeat DNA response element would bind a homodimers oriented in tandem with “opposite” receptor domains in close proximity, not something that had been previously reported for steroid receptors.
The resolution of this puzzle led to identification of a second class of nuclear hormone receptors, RXR, and expanded the mechanisms to modulate various TH signals. Early studies showed that the addition of nuclear extract enhanced THR binding and that these “heterodimers” were resistant to disruption by the addition of T3. 29 The identity of the heterodimer was later shown to be RXR, a member of the retinoid receptor family, which heterodimerized with THR and enhanced binding and transcription 30 (Fig. 2B), as well as heterodimerizing with retinoic acid receptor, vitamin D receptor, and nutrient receptors, such as peroxisome proliferator-activated receptors alpha and gamma. 31
THR Isoforms and THR Gene Products
It was recognized, as with some other steroid and steroid-like receptors, that there were two different THR isoforms, alpha (TRα) and beta (TRβ), with different expression developmentally, and in adult tissues, TRβ predominant in the pituitary and liver, TRα in the brain and skeletal muscle, and both TRα and TRβ in the heart. There were also several splice product variants of unknown significance. Much of the original descriptions examined mRNA expression in various tissues. The limited availability of high-affinity antibodies that reliably distinguished the THR isoforms limited research in this area.
The most readily available tool was to examine mRNA expression, but there was recognition of the potential discordance between mRNA and protein. The availability of specific antibodies was especially a challenge to distinguish the various isoforms. Perhaps, the most puzzling is TRα2, a TRα isoform with a truncated ligand domain resulting in loss of T3 binding and shown functionally to antagonize T3 action. 32 Despite high levels of TRα2 mRNA expressed, especially in the brain, the expression of the TRα2 protein has been difficult to demonstrate.
A wide range of genetic rodent models with THRB and THRA gene inactivation and mutations showed consistent phenotypes that generally matched the distribution of THR isoforms. 6,7 The initial knockout mice for THRB had a phenotype similar to that seen in humans with RTHβ, goiter, elevated serum T4, and nonsuppressed thyrotropin (TSH), 33 subsequent evaluation revealed a significant hearing defect. 34 A wide range of mice with dominant negative mutations of the TR genes that reduced ligand affinity and disrupted signaling by the wild-type TR, generally had a more robust phenotype. 35
There were also distinct phenotypes with mutations of THRA, although the metabolic phenotype varied depending on the THRA mutation that was being modeled. 1 The ultimate evidence of THR isoforms having distinct physiological function was the clinical report of individuals with mutations of the THRA gene. 36,37 Since TRα is not expressed significantly in the pituitary or hypothalamus, there was no impairment of T3 feedback, so serum TH levels remained largely normal, but there was evidence of reduced TH action with reduced growth, hypometabolism, bony defects, and constipation. Mutations of the THRA gene have been associated with skin tags and melanocytic nevi. 38
Early characterization of the THRA gene and associated transcripts identified a related protein that did not bind T3 or was involved in TH signaling. ReverbA is transcribed from the opposite strand of the THRA gene, did not bind T3, and the function was unknown. 39 This THR-related gene was subsequently shown to be a key link in the regulation of the circadian clock, serving as a heme sensor that is a negative regulator of the circadian clock and suppresses hepatic gluconeogenic gene expression and glucose output. 40
Basal Gene Repression by Unliganded THR
Despite some early skepticism about the validity of a direct repeat TRE in the rat growth hormone gene, others identified genes regulated by TH with a similar TRE configuration. 7 Although in vitro synthesized THR at high concentration could be shown to bind a direct repeat in a gel retardation assay, a major breakthrough was the recognition that the addition of nuclear extract dramatically increased binding. 29 The predominant protein in nuclear extract that was ultimately identified to bind THR was another nuclear receptor, RXR. 30 Another feature of this receptor family, first identified with THR, was that unliganded receptor bound DNA and repressed gene expression of positively regulated genes.
This was first shown with transient transfection assays and could be demonstrated as a receptor “dose” effect. 41 There was an underlying concern, however, that this repressive effect was an artifact of the transfection assay and that the high expression of heterologous promoters used in transient transfection could be consuming transcription factors that were artifactually lowering gene expression, a phenomenon referred to as “squelching.”
A wide range of subsequent studies confirmed that this repression was mediated by unliganded receptor, but more importantly, an underlying mechanism was identified, which dramatically expanded the ways in which TH signaling could be modulated. Corepressor binds to unliganded DNA-bound THR and represses gene expression. Addition of ligand disrupts corepressor binding and promotes binding of coactivator to THR, stimulating gene expression. The other receptors that form heterodimers with RXR have similar properties. Ultimately, a family of corepressors, the most prominent that binds THR, nuclear receptor corepressor (NCoR), and small mediator for retinoid/thyroid hormone, were identified and shown to be very significant in TH signaling (reviewed in Astapova and Hollenberg 42 ).
This was perhaps the most surprising property of TH that the clinical manifestations of hypothyroidism are not only the absence of ligand and reduced thyroid induced gene expression but also basal repression of genes based on occupancy of the response element by unliganded THR, which is bound to corepressor. The prediction from this finding is that the physiological consequences of reduced ligand in the presence of receptor would be partially restored toward normal in the absence of receptor. This was supported by a report of a mouse model with deletion of both THRA and THRB, which had a unique phenotype of retarded growth and bone maturation, reduced female fertility, and a hyperactive pituitary–thyroid axis. 43 Further evidence of the repressive role of TRα was a model with knockout of THRA, which makes up 80% of THR in the cerebellum that reversed the defects of hypothyroidism. 44
The role of NCoR was shown by an in vitro model expressing a mutant NCoR that does not bind to THR, showed that TH-dependent gene expression in the liver was upregulated when the mice were euthyroid, and expressed in the normal range when hypothyroid, demonstrating that NCoR physiologically regulated THR-dependent genes. 45 The repressive action of unliganded THR on positively regulated genes also led to an important connection of THR and related factors, to the effects of THR to mediate differentiation in development and then when those differentiation actions were disrupted, to promote oncogenesis.
Human Disorders of Genetic Defects of TH Signaling and Transport
A range of genetic syndromes of reduced TH action have provided important insights into mechanisms. 20,46 The syndrome, RTHβ, with clinical manifestations of goiter, elevated serum T4 concentration, and a nonsuppressed TSH, with variable bone and metabolic manifestations was initially described by Refetoff et al in 1967, 47 has provided key information on the physiological role of THR. Once the THR genes were cloned, the THRB gene was linked to RTH. 48 The initial family reported was ultimately shown to be homozygous for deletion of the THRB gene, but the majority of reported patients are heterozygous for a “dominant negative” mutation, most in the ligand binding domain, which results in interaction with corepressor, which is not relieved by the usual levels of ligand. 49,50 Animal models that selectively mutate the primary THR corepressor, NCoR, were shown to partially normalize thyroid signaling in models of RTH with a dominant negative THRB mutation. 51
A number of investigators identified membrane transporters for TH. 21 The specific transport of TH with high specificity was shown for the monocarboxylate transporter 8 (Mct8). 52 The linkage of Mct8 gene mutation to a previously described X-linked disorder, with intellectual deficit, hypotonia, spastic paraplegia, and hypermetabolism, demonstrated that the Mct8 transporter was essential for normal T3 action on brain development in humans. 53,54 The importance of the OATP1C1 transporter was demonstrated by the report of an adolescent girl with an OATP1C1 gene mutation and evidence for dementia and neurodegeneration, with some response to treatment with the TH analog Triac, which does not require a transporter to enter neurons. 55
Developmental Actions of TH
Sensory development, the inner ear and retina, have requirements for TH to progress normally (reviewed in Ng et al 56 ). This developmental process revealed a key principle of TH action in development, apparent from the earliest amphibian studies of TH in frog metamorphosis, where premature exposure to TH induced early metamorphosis (reviewed in Tata 5 ). TH availability in development is regulated by the deiodinase enzymes. Expression of the inactivating enzyme, 5-deiodinase type 3, reduces T3 levels and expression of the activating enzyme, 5′-deiodinase type 2, increases T3 levels. Premature or delayed exposure to T3 can result in developmental defects. This seems to be important for TH action in sensory development and the brain.
Although many studies of TH action have used cellular, rodent, and human models, key findings have come from amphibian models as well as more recently zebra fish (reviewed Tata 5 ). Xenopus has THRA and THRB, but there are gene duplications of both. Early models that tracked TH action showed distinct requirements for both key features of metamorphosis, tail resorption and limb development. Zebra fish also contain THRA and THRB and have been very useful developmental models.
Ligands and Analogues
The structural characterization of the ligand binding domain crystal structure in 1995 57 led to important insights. Distinct from T3 binding to the serum protein, TTR, which did not produce any conformational change in the protein, the prediction from the crystal structure of the ligand bound TRα was that the central hydrophobic core would be present in the absence of ligand, and the addition of T3 would induce a conformational change. Key residues in the ligand binding pocket also matched mutations associated with RTH, which included mutations of residues with direct contact with ligand as well those that disrupt stabilization of the bound confirmation.
Characterization of the THR ligand binding pocket also led to the design of compounds that were more selective for activation of TRβ or TRα. Most of the initial focus was on compounds that favored TRβ binding, with the therapeutic intent of targeting the beneficial metabolic actions of TH, weight loss, and lowering of cholesterol, without the adverse actions on the bone, heart, and skeletal muscle. GC1, or sobetirome, was among the earliest with a favorable profile of metabolic actions. 58 A broad range of compounds were identified with significant success, especially cholesterol lowering in statin-refractory patients; although the effects of these compounds on the bone and cartilage in animal toxicity testing have limited widespread use, there are a range of clinical applications being investigated. 59
Newer Findings in TH Action
The thyroid nuclear receptor is the fundamental pathway of TH action, but the impact of DNA response element sequence and location, coactivator and corepressor interactions, relative THR isoform expression, TH transport, and local ligand availability have all led to a recognition of factors that modulate TH signaling, especially at the local tissue level. An expanded description of modes of TH action 60 (Table 3; Fig. 2C) is more sophisticated mechanistically but recognizes a very similar range of actions as described in TH action studies from the 1970s (Table 2; Fig. 2A). Recognition of the wide range of TH action is especially relevant in consideration of thyroid replacement therapies. 61
Updated Nomenclature for Modes of Triiodothyronine Action
Adapted from Flamant et al. 60
RAR, retinoic acid receptor; T3, triiodothyronine; T4, thyroxine; THR, thyroid hormone receptor.
Noncanonical TH signaling pathways include TRα and TRβ interfacing in the cytoplasm with signaling pathways, such as PI3 kinase, and these actions are blocked by THR mutations in regions outside the ligand binding domain. 62 Direct tags of THR genes in rodent models showed that, in comparison to the original mRNA studies, TRα is the predominant isoform in most tissues, except the liver and pituitary, even in those tissues where THRA and THRB mRNA levels were similar. 63 Whole genome approaches have confirmed the core DNA response direct repeat consensus element sequence for the majority of TH signaling but also indicates TH signaling, which does not require direct DNA binding, especially for TH-mediated negative regulation. 64
The dogma of a direct ligand-induced switch from corepressor to coactivator is challenged by a study in the liver, showing that there is a ligand-induced “shift,” a predominance of coactivator or corepressor, rather than a complete “switch.” 64 THR resides predominantly in the nucleus, but cytoplasmic to nuclear TH transport may regulate some TH signaling. 65 THR undergoes post-translational modification by phosphorylation 66 and sumoylation, 67 which also impacts signaling and may respond to nutrient signals and signal the fed or fasted state. Nongenomic actions of TH, including direct histone modifications as well as membrane, mitochondria, and other targets, are being increasingly described and documented. 6
Clinical Applications of TH Action, Present and Future
TH has significant actions on stem cell differentiation and has been shown to stimulate stem cells in white and brown adipose tissue, pancreatic islet cells, skeletal muscle satellite cells, hepatocytes, and intestinal epithelial cells. 68 TRα has a specific role in skeletal muscle stem cells (satellite cells) essential for skeletal muscle development and regeneration after injury. 69 TH promotes recovery after brain injury as well as use in a range of neurological diseases. 70 TH is essential for bone development, and differential expression of THR isoforms underlies the clinical manifestations of reduced and excess TH. 71
TH action on the heart is central to the clinical manifestations of thyroid excess and deficiency and has also been a site with demonstration of the full range of genomic and nongenomic actions of TH. 72 Few agents are available to treat nonalcoholic fatty liver disease, but several clinical trials of T3, T4, as well as TH analogues, such as resmetirom, specifically acting or activated in the liver, show significant promise for treatment. 73 The liver-selective actions of resmetriom, a TRβ-selective analogue, are accomplished both by cell-specific transport and by TR isoform preference. 74
Author's Contributions
G.A.B. prepared this invited review and is solely responsible for all writing and editing and approval of the submitted version.
Footnotes
Author Disclosure Statement
G.A.B. confirms that there is no conflict of interest in relation to this work.
Funding Information
This work was partially supported by the United States Veteran Administration Merit Review 01BX001966 to G.A.B.