
Abstract
Background:
In Graves' orbitopathy (GO), localized orbital inflammation within the fixed orbit often leads to a fibrotic phenotype resulting in restrictive myopathy or refractory proptosis. However, the molecular pathways related to the transition from inflammation to fibrosis in GO are less understood. Yes-associated protein (YAP) and its homolog, transcriptional coactivator with PDZ-binding motif (TAZ; a Hippo pathway effector), are critical mechanosensors of mechanical stimuli and activate signaling cascades for cell proliferation, differentiation, and transformation. In this study, we aimed to examine the role of YAP in both inflammatory and fibrotic GO pathogenesis.
Methods:
Based on RNA sequencing performed on freshly obtained orbital adipose tissue from patients with GO and healthy individuals, Gene Ontology analysis and gene set-enrichment analysis were performed to analyze gene-expression differences between GO and normal orbital tissues. The role of YAP in GO-related inflammation and fibrosis was studied in primary cultured orbital fibroblasts. The effects of interleukin-1β (IL-1β)-induced inflammation and transforming growth factor-beta (TGF-β)-induced fibrosis on YAP expression were evaluated using real-time polymerase chain reaction and Western blotting analyses. The effects of YAP on inflammatory and fibrotic responses were also examined by YAP silencing or treatment with pharmacological YAP inhibitors.
Results:
RNA sequencing revealed enhanced YAP expression in GO orbital tissues. Gene Ontology analysis indicated that “response to mechanical stimulus”-related genes were overexpressed in GO orbital tissues, along with those enriched for the “adipose proliferation,” “inflammatory responses,” and “hormone stimulus responses” terms. IL-1β did not enhance YAP expression, and YAP silencing decreased IL-1β-induced IL-6 expression while increasing prostaglandin-endoperoxide synthase 2 expression, leading to paradoxical pro-inflammatory effects. Conversely, TGF-β enhanced YAP expression, and YAP silencing and pharmacological YAP inhibitor (cerivastatin, verteporfin, TED-347, and CA3) treatment significantly reduced TGF-β-induced myofibroblast differentiation and collagen formation.
Conclusion:
YAP, a mechanotransducer responding to mechanical stimuli, was strongly expressed in GO orbital tissues, and YAP was induced by TGF-β in orbital fibroblasts. Our study establishes YAP as a novel mediator of GO pathobiology, potentially mediating the transition from early inflammation to chronic fibrosis in GO. The finding that YAP inhibition suppressed TGF-β-induced fibrotic response suggests YAP as a therapeutic target against the fibrotic mechanism of GO.
Introduction
Graves' orbitopathy (GO) is an orbital autoimmune disorder and the main extrathyroidal pathological manifestation of Graves' disease, the most common cause of hyperthyroidism. 1,2 The symptoms of GO include ocular discomfort, eyelid retraction, periorbital tissue edema, proptosis, diplopia, and vision loss in the most severe cases. 3,4 Although the pathogenic mechanism of GO remains to be fully elucidated, orbital fibroblasts are considered as key cells involved in the immunologic mechanism of inflammation, adipogenesis, and proliferation.
Enhanced adipogenesis, proliferation of orbital fibroblasts, and glycosaminoglycan accumulation between the muscle fibers and within the adipose connective tissues in the early active phase of GO lead to characteristic tissue remodeling, which manifests in an increased orbital soft tissue volume within the fixed orbital bony space causing eyeball proptosis and congestion. 5,6 Hypertrophy and congestion of extraocular muscles with impaired venous return possibly result in compressive optic neuropathy in very severe cases of GO.
A broad range of pro-inflammatory stimuli and chronic tissue remodeling may exacerbate the fibrotic process in the orbit, which often limits effective response to anti-inflammatory treatment and may cause refractoriness to surgical decompression. A knowledge gap exists regarding how orbital tissues in GO become fibrotic and which molecular mechanisms are involved in the switch from inflammation to fibrosis in orbital fibroblasts. 2,7
The Hippo signaling pathway is a key determinant of organ size, stem cell homeostasis, cellular proliferation, development, and wound healing. 8,9 Yes-associated protein (YAP) and its paralog, transcriptional coactivator with PDZ-binding motif (TAZ), are transcriptional coactivators that serve as Hippo pathway signal transducers, whose nuclear and cytoplasmic distributions are in turn controlled mainly by the Hippo pathway. 10 It is known that G-protein-coupled receptors activate the YAP/TAZ and Hippo pathways. 11 Recently, it has been shown that mechanical stress can trigger YAP/TAZ by detecting cytoskeletal stress, resulting in cellular mechanoresponses. 12
Recent studies have focused on the roles of YAP in inflammation and fibrosis. 13,14 YAP inactivation and consequent prostaglandin-endoperoxide synthase 2 (PTGS2) upregulation were previously linked to endoplasmic reticulum stress and persistent inflammation. 14 YAP hyperactivation also induces fibrosis in cancers and other diseases. 15 In idiopathic pulmonary fibrosis fibroblasts, small-interfering RNA (siRNA)-mediated YAP silencing reversed the pro-fibrotic phenotype by downregulating genes that mediate transforming growth factor-beta (TGF-β)-dependent cross-linking of the extracellular matrix (ECM), while boosting the expression of genes essential for matrix degradation. 16
This study was designed to investigate the role of YAP in the pathogenesis of GO. By performing RNA sequencing and Gene Ontology analysis, we demonstrated different molecular features in GO orbital tissues and healthy controls. The alteration of YAP expression under pro-inflammatory and pro-fibrotic stimuli was identified in primary cultured orbital fibroblasts. We also probed the effect of YAP silencing or pharmacological YAP inhibition on inflammatory and fibrotic pathogenesis in GO.
Materials and Methods
Reagents
The reagents used in this study are presented in the Supplementary Data.
Study approval
This study was approved by the Institutional Review Board (IRB)/Ethics Committee of Severance Hospital, Yonsei University Health System (IRB No. 4-2021-0568). The study was conducted following the Declaration of Helsinki and adhered to good clinical practice guidelines. Written informed consent was obtained from all participants.
Culturing primary human orbital fibroblasts
Human primary orbital fibroblast cultures were prepared as described previously. 17,18 Specifically, orbital explants containing adipose and connective tissues were obtained as surgical waste from 12 patients with GO (7 women and 5 men; 21–59 years of age) who underwent orbital-decompression surgery. The patients had clinical activity scores of ≤3 and were in a stable euthyroid condition at the time of the surgery.
In addition, for at least 3 months before surgery, none of the patients received radiation, any immunomodulatory therapies, or steroids. Normal control tissues were harvested from the postseptal areas of six individuals with no history of GO (four women and two men; 27–64 years of age) during upper-lid blepharoplasties or other orbital surgeries. The clinical information of the donors is presented in Supplementary Table S1.
The tissue explants were minced and plated in 10 cm culture dishes with Dulbecco's modified Eagle's medium/Ham's F12 (DMEM/F-12) containing 20% fetal bovine serum (FBS) and 1% penicillin/streptomycin. After fibroblasts had grown from the explants, they were passaged after detachment with trypsin/ethylenediaminetetraacetic acid, and the cultures were maintained in high-glucose DMEM containing 10% FBS and 1% penicillin/streptomycin using conventional cell culture techniques. Fibroblasts from at least three samples from patients with GO were used in each study.
siRNA and reagents
Reverse siRNA transfections were performed with Lipofectamine RNAiMAX (Invitrogen, Waltham, MA). AllStars Negative Control siRNA (Qiagen, Hilden, Germany) was used as a control siRNA. The following siRNAs were used in this study: YAP: 5′-GACAUCUUCUGGUCAGAGAdTdT-3′; TAZ: 5′-ACGUUGACUUAGGAACUUUdTdT-3′.
RNA sequencing
RNA was extracted from orbital explants containing adipose and connective tissues using an RNeasy Kit (Qiagen). RNA sequencing was performed by MacroGen, Inc. (Seoul, South Korea) using the Illumina Hisequation 2000 platform (San Diego, CA). A TruSeq Stranded Total RNA LT Sample Prep Kit (Gold; Illumina) was used to construct a complementary DNA (cDNA) library. FastQC (Babraham Bioinformatics, Cambridge, United Kingdom) was used to assess the quality of the raw sequence data, and Trimmomatic (version 0.36; USADELLAB, Aachen, Germany) was used to filter the data. HISAT2 was applied to match the trimmed data to the human reference genome (hg19).
After filtering, only transcripts with gene identifications, read counts, fragments per kilobase of transcript per million mapped reads, and transcripts per kilobase million remained. DEseq2 software (Illumina) was used to normalize the filtered read-count data, and variance stabilization was applied. Database for annotation, visualization, and integrated discovery (DAVID) software was used for Gene Ontology annotation analysis of upregulated and downregulated genes.
Gene Ontology is a structured bioinformatics model for studying individual gene functions according to their related biological processes, molecular functions, and cellular components, 19 which we used to identify functional annotations for differentially expressed genes between GO and control orbital tissues. The C1 hallmark and C6 oncogenic gene sets were used for gene set-enrichment analysis (GSEA; MSigDB; Broad Institute, Cambridge, MA and UC San Diego).
Quantitative reverse transcriptase-polymerase chain reaction analysis
Total RNA from orbital adipose/connective tissue explants and primary orbital fibroblasts was extracted using an RNeasy Kit. In the presence of RNase Inhibitor, 1 g of isolated RNA was annealed with oligo dT primers (Roche Life Science, Basel, Switzerland) and reverse transcribed to cDNA using Moloney-Murine Leukemia Virus Reverse Transcriptase (RNasin Plus; Promega, Madison, WI). Messenger RNA (mRNA)-expression levels were evaluated using cDNA, customized primers, SYBR Green Supermix (Bio-Rad, Hercules, CA), and a CFX96 real-time quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) system (Bio-Rad). The sequences of the customized primers are presented in the Supplementary Data.
Western blot analysis
Cells were lysed on ice in radioimmunoprecipitation assay buffer (20 mM HEPES, 150 mM NaCl, 1% NP-40, 0.25% sodium deoxycholate, and 10% glycerol) supplemented with protease and phosphatase inhibitors (Merck Millipore, St. Louis, MO) before being subjected to Western blot analysis. The resulting cell lysates were sonicated with a Branson Digital Sonifier 250 (Branson Ultrasonics, Danbury, CT) and centrifuged (18,000 × g) for 10 minutes at 4°C. Supernatant protein concentrations were measured using a BCA Protein Assay Kit (Pierce Chemical Co., Dallas, TX). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis was performed with aliquots of each protein lysate (10–20 g).
After electrophoresis, the proteins were transferred to nitrocellulose membranes, and the membranes were incubated with blocking solution (5% skim milk and Tris-buffered saline with 0.1% Tween 20) for 45 minutes. The blocked membranes were incubated with primary antibodies overnight at 4°C and then with appropriate secondary antibodies (Abcam, Cambridge, United Kingdom) for 1 hour at 4°C. Proteins were detected using WesternBright Enhanced Chemiluminescence Western Blot Detection Solution (Advansta, Inc., San Jose, CA).
Quantification and statistical analysis
In all experiments, we analyzed cells from at least three different samples, where each sample was tested in duplicate. R software version 4.1.0 (R Foundation, Vienna, Austria) and GraphPad Prism software version 6 (GraphPad Software, San Diego, CA) were used to perform Student's t-tests or Wilcoxon's rank-sum tests to compare parameter estimates between the experimental and control groups. p-Values of <0.05 indicated statistical significance.
Results
Signaling pathways upregulated in GO
We performed RNA sequencing to identify genes and signaling pathways upregulated in GO, using orbital adipose/connective tissue explants obtained from patients with GO and healthy controls. RNA sequencing revealed 34 significantly upregulated genes and 52 downregulated genes in GO (Fig. 1A and Supplementary Table S2; greater than twofold change in expression and p < 0.05). Interleukin-6 (IL-6), connective tissue growth factor (CTGF), and cysteine-rich angiogenic inducer 61 (CYR61) were identified among the upregulated genes.
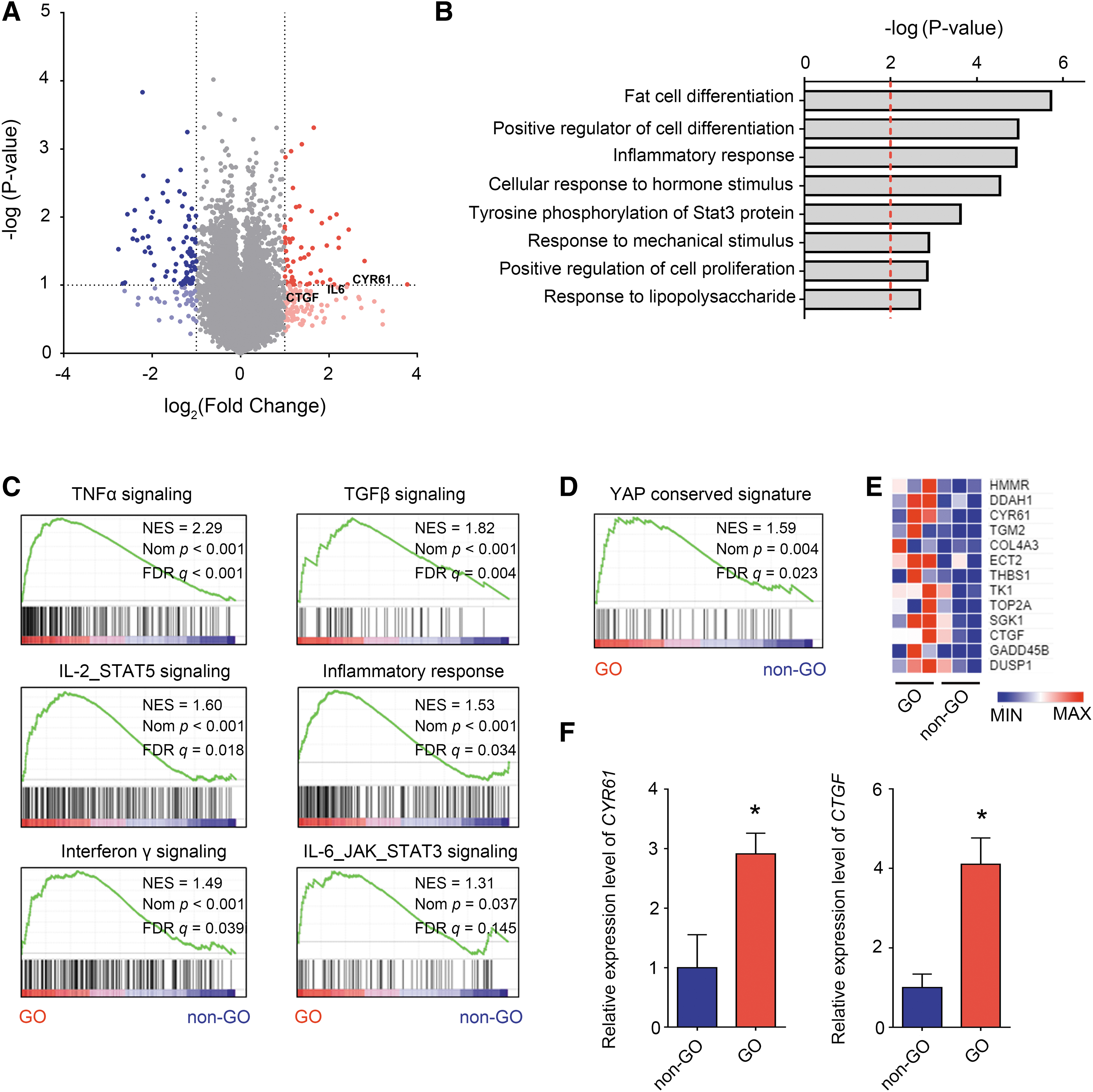
Elevated YAP activity in GO orbital tissues. Orbital tissues from individuals with GO and healthy controls (n = 3, each) were harvested, and RNA sequencing was performed to identify genes and signaling pathways upregulated in GO. (
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis showed that the terms “fat cell differentiation,” “inflammatory response,” “STAT3 signaling,” and “response to mechanical stimulus” were significantly enriched in GO (Fig. 1B). GSEA revealed increased signaling through the tumor necrosis factor-alpha, TGF-β, interleukin 2–signal transducer and activator of transcription 5 (IL-2–STAT5), inflammatory response, interferon-γ, and interleukin 6–Janus kinase–STAT3 (IL-6–JAK–STAT3) signaling pathways (Fig. 1C).
In addition to the increased responses to mechanical stimuli found by Gene Ontology analysis, an increased YAP signature was identified by GSEA (Fig. 1D, E). Representative YAP target genes, CYR61 and CTGF, were upregulated in GO orbital tissues (Fig. 1F).
Paradoxical effect of YAP suppression during IL-1β-mediated inflammation
IL-1β is the major mediator of the inflammation phase in GO. Administering IL-1β to orbital primary fibroblasts increased the expression of IL-6, IL-8, and PTGS2, which are essential for inflammatory responses in GO (Fig. 2A). In contrast, IL-1β did not influence the expression of YAP and TAZ, as indicated by the observation that the IL-1β-induced changes in YAP's target genes were less pronounced; a relatively small increase in CYR61 was noted, and CTGF expression was slightly (but not significantly) lower (Fig. 2B).
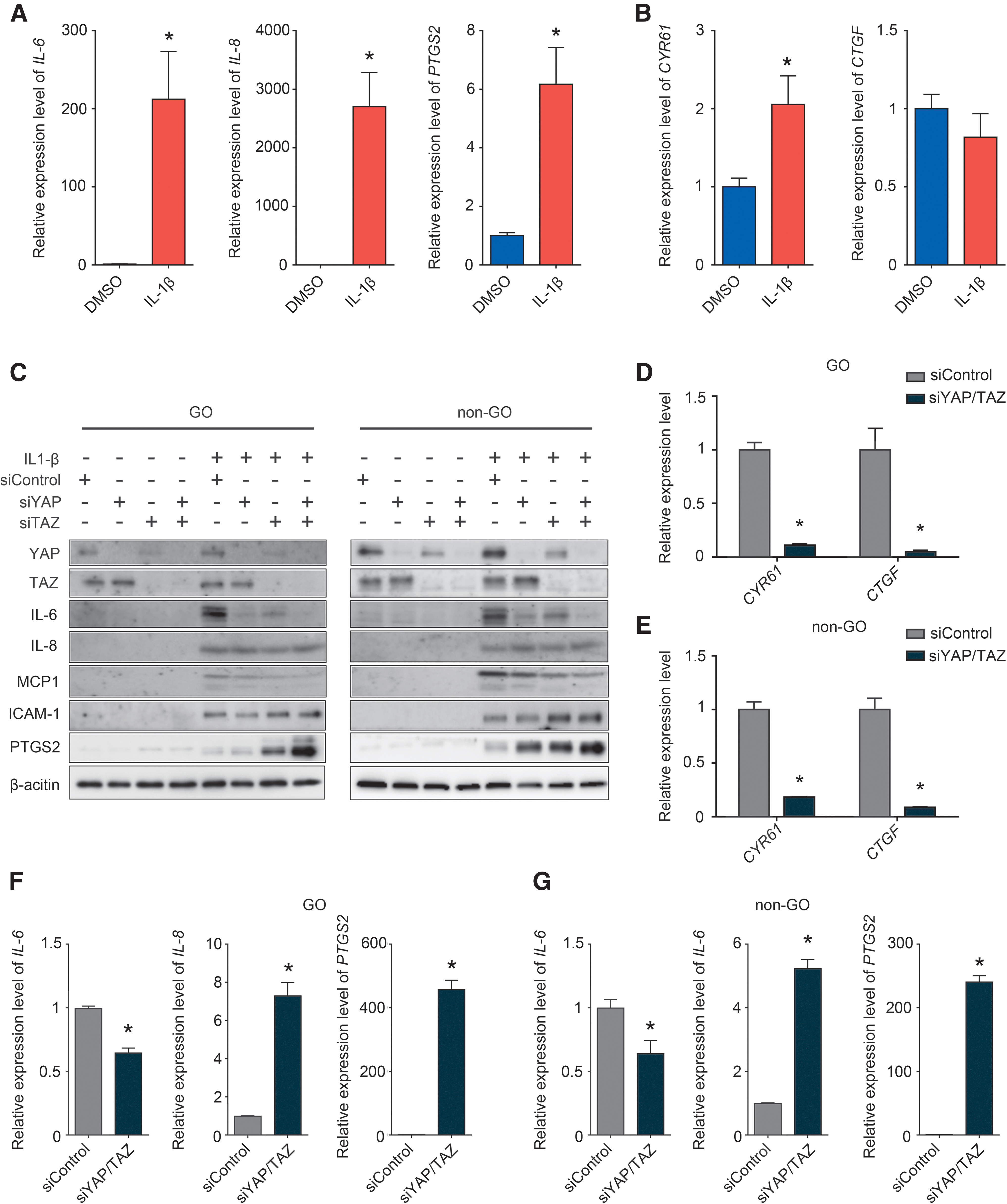
Paradoxical effect of YAP depletion during IL-1β-mediated inflammation. (
In GO and non-GO orbital primary fibroblasts, siRNA-mediated YAP/TAZ inhibition reduced IL-6 and monocyte chemoattractant protein expression and increased PTGS2 and intercellular adhesion molecule 1 expression (Fig. 2C). qRT-PCR analysis showed that silencing YAP/TAZ with siRNAs in GO and non-GO primary orbital fibroblasts significantly downregulated YAP target genes (CYR61 and CTGF) (Fig. 2D, E). Similar to our immunoblotting results, qRT-PCR revealed that siRNA-mediated YAP suppression reduced IL-6 expression while increasing IL-8 and PTGS2 expression (Fig. 2F).
YAP mediated TGF-β-induced fibrosis
TGF-β is the key mediator of the fibrosis phase of GO. Administering TGF-β to primary GO orbital fibroblasts significantly increased the expression levels of alpha smooth muscle actin (αSMA), fibronectin 1 (FN1), and collagen type I alpha 1 (COL1A1) (Fig. 3A). Unlike IL-1β treatment, when TGF-β was applied to induce fibrosis, the CYR61 and CTGF expression levels increased significantly, representing TGF-β-induced YAP activation (Fig. 3B).

YAP may mediate TGF-β-induced fibrosis in primary cultured orbital fibrosis. (
Western blotting showed that TGF-β increased YAP and TAZ expression. Furthermore, siRNA-mediated silencing of YAP, and TAZ decreased TGF-β-induced COL1α and αSMA expression, and the reduction was more profound in both YAP siRNA (siYAP) and TAZ siRNA (siTAZ) co-transfected cells (Fig. 3C). qRT-PCR analysis showed that siYAP/TAZ treatment caused downregulation of CTGF (a YAP target gene) and αSMA (Fig. 3D, E).
Pharmacological inhibition of YAP relieved TGF-β-induced fibrosis
We investigated whether pharmacological inhibition of YAP activity might reduce TGF-β-induced fibrosis. Cerivastatin and verteporfin are drugs that are well known to reduce YAP activity. Cerivastatin causes phosphorylation of Ser127 in YAP, resulting in its cytoplasmic sequestration. Verteporfin inhibits interactions between YAP and TEA domain family members, resulting in lower transcriptional activity. CTGF, CYR61, αSMA, and COL1A1 expression decreased in GO and non-GO fibroblasts in the presence of each of these distinct YAP inhibitors (Fig. 4A–D).
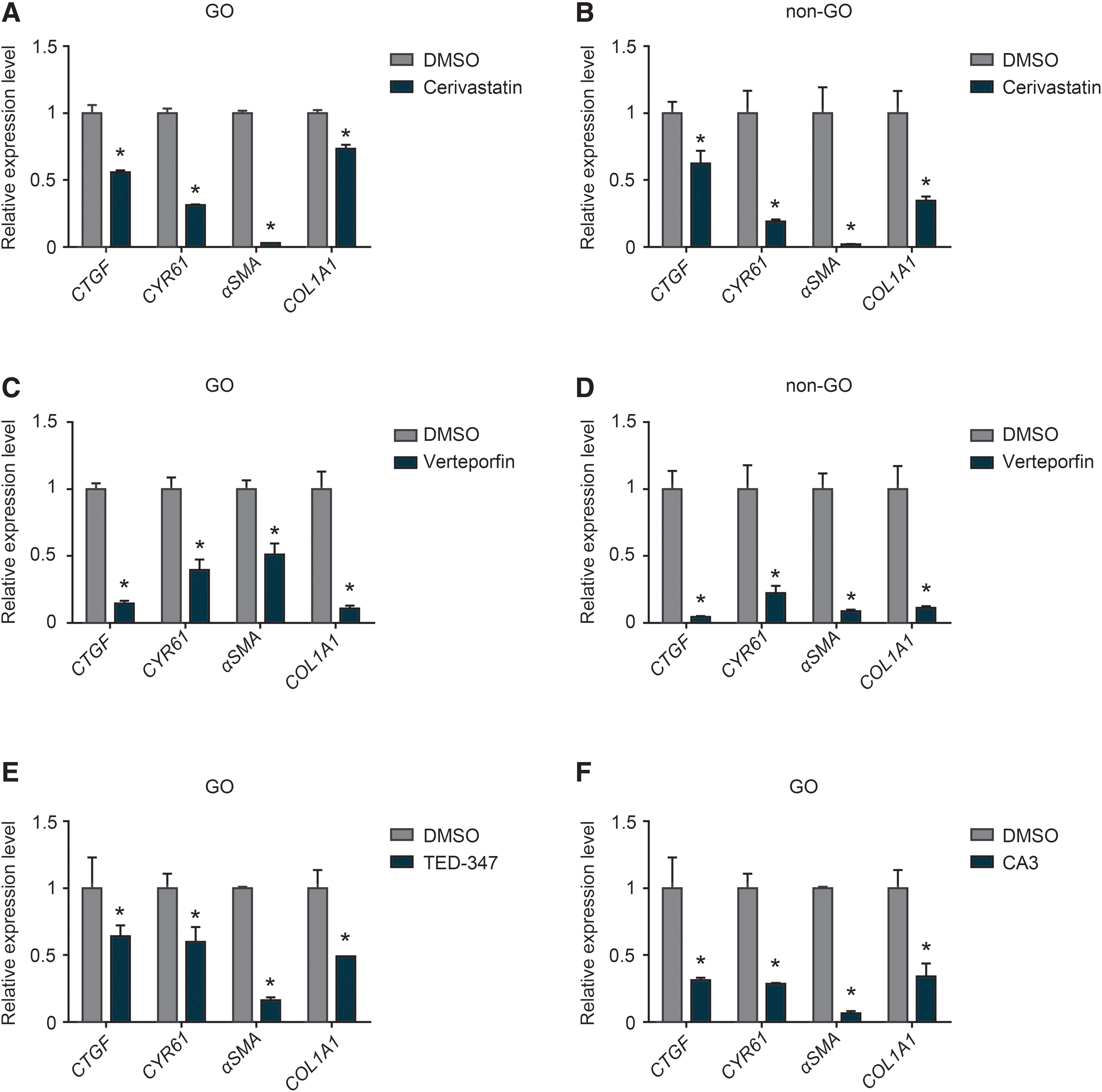
Pharmacological inhibition of YAP/TAZ decreased the expression of genes related to TGF-β-induced fibrosis. Confluent orbital fibroblasts derived from individuals with or without GO were either not pretreated or were pretreated with a pharmacological YAP/TAZ inhibitor (
The reduction in αSMA expression was often more substantial, suggesting that YAP inhibition interferes with myofibroblast differentiation of orbital fibroblast under pro-fibrotic stimulation. Similar findings were found in GO fibroblasts treated with TED-347 and CA3 (Fig. 4E, F), which have recently been shown to inhibit YAP activity. 20
Discussion
In this study, we found that GO orbital tissues had higher expression levels of the YAP-associated genes and its downstream target molecules than healthy normal orbital tissues. TGF-β potently increased YAP expression in primary cultured orbital fibroblasts, whereas IL-1β did not. YAP showed a paradoxical role in the pro-inflammatory pathogenesis in GO; YAP silencing suppressed IL-1β-induced IL-6 expression while inducing the expression of IL-8 and PTGS2.
In contrast, YAP played a consistent role in the pro-fibrotic pathogenesis in GO; TGF-β-induced myofibroblast differentiation and collagen synthesis were suppressed by YAP knockdown or pharmacological YAP-inhibitor treatment. Based on our results, the YAP/TAZ signaling pathway, identified as a key mechanotransducer, may play a significant role in the GO pathogenic mechanism, and its modification can potentially serve as a therapeutic target.
Transcriptome analysis of GO orbital tissues using RNA-sequencing technology has shown that genes responsible for mediating inflammation, cytokine signaling, and adipogenesis are upregulated in GO orbital tissues. 21 –23 Our KEGG and GSEA analyses in this study confirmed that adipocyte differentiation, inflammatory responses, and the STAT3 signaling pathway were augmented. Our transcriptome-analysis findings are consistent with previous results describing the pathophysiology of GO. 2,21,22,24,25
YAP and its homolog TAZ (a Hippo pathway effector) have been identified as key mechanotransducers that sense mechanical stimuli and relay signals that control transcriptional programs for cell proliferation, differentiation, and transformation. Additional findings showed that cellular mechanoresponses are mediated by YAP/TAZ, which sense cytoskeletal tension. 12 To transform mechanical stimuli into cell-specific transcriptional programs, YAP and TAZ read a wide range of mechanical cues, including shear stress, cell shape, and ECM rigidity. 12
YAP and TAZ mechanotransduction is critical for modulating stem cell behavior and regeneration, and is responsible for the aberrant cell mechanics that are instrumental for the onset of multiple diseases. 26 Activation of the YAP pathway by weight-bearing activity has recently been linked to the development of plantar melanoma, demonstrating that increased pressure can activate YAP in vivo. 27
Our RNA-sequencing data showed that GO orbital tissues expressed enhanced levels of genes related to responses to mechanical stimuli. Through RNA-sequencing and Gene Ontology analysis, we demonstrated that GO orbital tissues expressed higher levels of genes associated with reactions to mechanical stimuli than healthy orbital tissues. Specifically, the expression levels of YAP and YAP-target genes were elevated in GO tissues. It is thought that in GO, orbital adipocyte proliferation and inflammation lead to orbital tissue expansion and that the accompanying elevation in orbital pressure stimulates YAP/TAZ signaling to sense mechanical stimuli and convey the signal to control transcriptional programs for homeostasis.
The role of signal integration between the TGF-β and YAP/TAZ pathways in the initiation of organ fibrosis has received much attention recently. 10,28 Previous data showed that YAP and TAZ might trigger the TGF-β/SMAD3 signaling pathway (a critical signaling pathway in ECM homeostasis) during human skin fibroblast development and that the YAP/TAZ and TGF-β signaling systems were connected. 28,29 In this study, we found that TGF-β stimulation elevated the expression of YAP and TAZ, implicating YAP's role in the pathogenic process of GO fibrosis.
This association is supported by the fact that CTGF, a representative target molecule of YAP, is widely documented to contribute to myofibroblast transdifferentiation in GO orbital fibroblasts. 30 In addition, fibrosis markers (αSMA and COL1A1) were consistently suppressed by YAP knockdown or pharmacological YAP suppression, suggesting that YAP is a potential therapeutic target for blocking fibrosis in GO.
GO pathogenesis is thought to involve secondary adipogenesis and fibrosis caused by primary orbital autoimmune related inflammation. Thus, most known GO-related etiologies have similar impacts on adipogenesis, fibrosis, and inflammation, 7,17,24,31 indicating that most of the pro-inflammatory response pathways in GO lead to increased adipogenesis or fibrosis. However, in this study, we found paradoxical roles for YAP in inflammation and fibrosis.
Pro-inflammatory stimuli did not trigger the expression of YAP/TAZ, and its target molecules, CYR61 and CTGF, did not exhibit any substantial alterations under pro-inflammatory stimuli (Fig. 2B, C), in contrast to the TGF-β-driven increase in the expression of YAP and YAP target molecules (Fig. 3C). In addition, YAP showed a paradoxical effect on pro-inflammatory signaling in GO orbital fibroblasts; silencing YAP suppressed IL-1β-induced IL-6 expression while boosting IL-8 and PTGS2 expression (Fig. 2F).
YAP serves a complex role in inflammation by exerting both pro-inflammatory and anti-inflammatory effects under different conditions. 32 Studying crosstalk between the Hippo-YAP pathway and nuclear factor kappa B (NF-κB) signaling (which is critical in inflammatory and immune responses) revealed that YAP blocked NF-κB activation and that the YAP/TAZ-TEAD complex inhibited the transcriptional activation of NF-κB-targeted genes. 33 These findings were also demonstrated in a disease model, confirming that YAP activation directly attenuated NF-κB signaling resulting in an anti-inflammatory effect that blocked disease progression. 34,35
Considering that IL-8 and PTGS2 are effectors of the NF-κB pathway, their overexpression during YAP/TAZ silencing (Fig. 2F) may reflect the anti-inflammatory role of YAP in GO via crosstalk with NF-κB. However, several pro-inflammatory cytokines (such as monocyte chemoattractant protein 1, tumor necrosis factor-α, and IL-6) are stimulated by YAP after associating with the TEAD-binding motif in the promoter regions of genes encoding inflammatory cytokines, explaining our results showing that silencing YAP suppressed IL-1β-induced IL-6 expression. 32
Although it is difficult to ascertain the role of YAP in the inflammatory phase due to such conflicting inflammatory responses to YAP, the facts that pro-inflammatory stimuli do not increase YAP expression and activation suggest that low YAP activity is maintained in the early inflammatory phase of GO. After the onset of the late quiescent phase, YAP expression increases, presumably because of elevated orbital pressure and stimulation due to TGF-β release from adjacent lymphocytes. 36 This enhanced YAP activity is thought to enhance the fibrotic response at the later quiescent phase.
The absence of evidence that increasing mechanical pressure promotes YAP activity in GO orbital tissues constitutes a significant limitation of this study. Increased mechanical stress and YAP activity have been extensively documented in other tissues and linked to the onset of diseases. 12,26,27 However, we were unable to confirm this connection in GO orbital tissues owing to the lack of a tissue culture model of mechanical stress and an established animal model of GO. 25 If low YAP activity in early inflammatory-phase GO orbital tissues could be evaluated, then such findings would be compared with the substantially enhanced YAP activity in the late inactive-phase tissues shown in this study.
However, it is practically difficult to obtain inflamed orbital tissue during the initial active stage of GO because surgical decompression is generally not required during this phase. 3,37 Although orbital decompression is carried out in patients with dysthyroid optic neuropathy during the active GO stage, surgery is usually considered during or after intravenous high-dose steroid treatment. 38 Because corticosteroid treatments are likely to alter the molecular phenotype of orbital fibroblasts, we examined tissues from patients who had not been treated with steroids for at least 3 months.
This study focuses on the little-understood GO pathophysiology at the point when the inflammatory phase changes into the fibrotic phase. This study revealed for the first time, to our knowledge, that increased orbital pressure can contribute to GO pathogenesis, and the YAP/TAZ system appears to contribute to that process. A limitation of this study is that it did not include data on GO tissue in the acute inflammatory phase before the transition from inflammation to fibrosis. Future analysis of GO orbital tissue at the hyperacute stage before orbital pressure increases or research using a reliable animal model will allow for further clarification of this.
In summary, we demonstrated that YAP and other genes that respond to mechanical stimuli were overexpressed in orbital adipose tissues in patients with chronic inactive GO. YAP (induced by TGF-β and potentially by elevated orbital pressure during disease progression) might mediate TGF-β-induced orbital fibrosis. The possibility exists that YAP facilitates the transition from the early inflammatory phase of GO to the later fibrotic phase.
Furthermore, because YAP is targeted by several statins, the current findings suggest further explanations for the favorable clinical effects of statins reported previously. 39 Blocking YAP by a pharmacological agent such as the FDA-approved compound, vertoporfin, or statins might have an additive therapeutic effect on GO fibrotic mechanism, as blocking YAP downregulated IGF-1R signaling-related protein. 40 Further investigations are required to elucidate the relationship of YAP/TAZ signaling pathway with the inflammation/fibrosis mechanism of GO in vivo.
Footnotes
Authors' Contributions
Validation, resources, data curation, writing—original draft, writing—review and editing, and project administration by J.K. Conceptualization, methodology, validation, formal analysis, investigation, data curation, writing—original draft, writing—review and editing, and visualization by Y.J.K. Formal analysis, investigation, and visualization by S.H.C. Validation, formal analysis, writing—review and editing, and supervision by C.S.L. Conceptualization, methodology, validation, resources, writing—review and editing, supervision, project administration, and funding acquisition by J.S.Y.
Author Disclosure Statement
The authors declare that no competing financial interests exist.
Funding Information
J.K., Y.J.K., S.H.C., and C.S.L. declare no funding information. This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Science and ICT (grant No. NRF-2021R1F1A1046652 to J.S.Y.).
Supplementary Material
Supplementary Data
Supplementary Table S1
Supplementary Table S2