
Abstract
Objective:
Deiodinase type I (DIO1) is crucial in maintaining thyroid hormone (TH) balance. It converts the prohormone thyroxine (T4) to the active triiodothyronine (T3) and degrades T3 to inactive 3,3′-diiodothyronine (3,3′-T2). It also acts on reverse T3 (rT3) and sulfated TH metabolites, thus contributing to TH elimination. Upregulation of DIO1 is linked to hyperthyroid conditions such as Graves’ disease and autonomous thyroid adenoma, making it a promising target for pharmacological intervention. The adverse side effects of the antithyroid drug propylthiouracil (PTU), used in clinics to treat hyperthyroidism due to its thyroid peroxidase- and DIO1-blocking action, highlight the need for novel and potent DIO1-selective inhibitors.
Methods:
Using a semiautomatic high-throughput screening (HTS) assay based on the Sandell–Kolthoff (SK) reaction in 384-well plates, we screened 69,344 low-molecular-weight compounds for DIO1-inhibitory effects. Shortlisted hits underwent detailed manual characterization, where we evaluated the potency and isoenzyme specificity by assessing their DIO-inhibitory effects on enzyme preparations from all three DIO isoenzymes, over a wide concentration range (5 nM–20 µM). To evaluate the DIO1 inhibitory effects in intact cells, we applied a novel protocol based on the SK reaction to cell culture supernatants and assessed the intracellular deiodinase activity in DIO1 overexpressing HEK293 cells.
Results:
The robust HTS assay flagged 436 (<1%) of the screened compounds as hits, also including known DIO1 inhibitors such as PTU and genistein. Based on a validation screen of 298 compounds, we prioritized 26 compounds to comprehensively characterize their DIO1-selective inhibition. We identified 15 DIO1-selective compounds (IC50 < 1 µM), more potent than the bonafide DIO1-selective inhibitor PTU. Additionally, 8 of the 13 tested compounds were found capable of inhibiting DIO1 in intact cells.
Conclusions:
With a successful SK-reaction-based HTS application, we identified novel, potent, and selective inhibitors of DIO1 with nanomolar IC50 values. Furthermore, we successfully showed that some of these compounds were also capable of inhibiting intracellular DIO1 in intact cells. These novel compounds hold immense potential in studying TH modulation, deciphering DIO1 enzyme structure, and developing structure–activity relationships. Furthermore, our novel inhibitors act as lead compounds in developing strategies to combat hyperthyroidism.
Introduction
Thyroid hormones (THs), upon secretion by the thyroid gland, regulate important physiological processes such as energy homeostasis, thermogenesis, and oxygen consumption and exert a broad range of effects on the development and growth of an organism.
1,2
The most biologically active form of TH, T3 (3,3′,5-triiodo-
DIO1 and DIO2 are generally referred to as TH activating enzymes as they convert the prohormone T4 (3,3′,5,5′-tetraiodo-
The direct regulation of DIO1 by T3 explains the often found, elevated levels of DIO1 in hyperthyroid conditions such as Graves’ disease and autonomous thyroid adenoma. 20 –22 This elevated DIO1 activity is further implicated into the increased conversion of T4 to T3, forming a vicious cycle and worsening the hyperthyroid symptoms. In clinical practice, PTU is used as an antithyroid drug in the treatment of Graves’ hyperthyroidism for over 80 years as it inhibits TH synthesis by blocking thyroid peroxidase (TPO). Additionally, the DIO1-inhibitory effect of PTU is considered advantageous in such hyperthyroid conditions. 23 However, PTU and other thioamide antithyroid drugs such as methimazole and carbimazole are associated with adverse hepatic, cutaneous, and hematological reactions. 24,25 These effects typically occur in early treatment and are reversible upon discontinuation of the drug. While serious side effects such as hepatotoxicity, agranulocytosis, and vasculitis are rare, methimazole’s effects appear to be dose-related, whereas those of PTU are less clearly linked to dose. 26 Additionally, in life-threatening situations of acute thyrotoxicosis/thyroid storm, iopanoic acid, a triple-iodized radiocontrast agent known to inhibit all DIO isoenzymes, has been successfully used in reducing the peripheral T4 to T3 conversion and normalizing the serum concentrations of T4 and T3. 15,27 –29 However, with the development of safer alternatives and improved radiocontrast agents, iopanoic acid has been discontinued. Therefore, in the treatment of hyperthyroid states such as Graves’ hyperthyroidism and thyroid storm, a potent inhibitor of DIO1 that does not affect TH synthesis via TPO inhibition but directly targets TH conversion may prove extremely valuable. Targeting selenocysteine with seleno-analogues of 2-thiouracil to increase the potency and isoenzyme specificity of DIO1 inhibitors was successful in increasing potency but failed in terms of DIO-selectivity 30 –33 and might affect also other selenocysteine-containing enzymes. Several heavy metal cations were also found to rapidly and irreversibly block deiodination by targeting selenocysteine and other SH groups found in the active sites of DIOs. However, they are also shown to cause heavy metal toxicity, especially during fetal development. 34
To examine the TH axis and factors affecting TH status, radioactive DIO assays monitoring the amount of radioactive iodide released (I-) from labeled tracer TH substrates as well as RIAs measuring DIO substrate-product ratios were developed. 35 Furthermore, recent advances in liquid chromatography–tandem mass spectrometry methods to separate and quantify THM provided a more detailed and complete picture of the DIO reaction. To overcome the complex workflows and high assay costs, a nonradioactive assay that measured the iodide released from the TH substrates using the Sandell–Kolthoff (SK) reaction was developed recently. 36,37 This nonradioactive assay readily allowed for a first screening of environmental toxicants and specific chemicals that disturb the TH system. 38 –41 However, studies encompassing a larger chemical universe with a focus on DIO1-specific inhibition besides environmental chemicals are lacking.
In this study, we evaluated a diverse set of chemicals containing multiple small molecule libraries utilizing a high-throughput screening (HTS) assay to identify potent and isoenzyme-specific inhibitors of DIO1. While previous works utilized a 96-well plate approach, we increased the throughput to a 384-well plate by optimizing and evolving DIO1-assay parameters and handling steps. We efficiently analyzed the effects of 69,344 compounds with small molecular weights on DIO1 activity. With this comprehensive screening effort, we identified several compounds that exhibited significant inhibition of DIO1 in the primary screens. We further validated these compounds in secondary screens and finally in a 96-well plate format to select the most potent and isoenzyme-specific inhibitors of DIO1. Our findings not only identify novel and potent inhibitors of DIO1 but also provide deeper insights into their usability and effects in intact cells. This article presents the results of our HTS assay, verification of the DIO1-inhibitors, analysis of their inhibitory potency and isoenzyme-selectivity, and assessment of DIO1-inhibitory potential in an intact cell system.
Materials and Methods
Selection of small molecule libraries
A total of 69,344 compounds with low-molecular-weight belonging to various compound libraries available at the Screening Unit core facility of the Leibniz-Forschungsinstitut für Molekulare Pharmakologie (FMP), Berlin, were included in the selection process. Table 1 provides the number of compounds in the compound libraries chosen for screening purposes. These libraries consist of compounds either derived or sourced from diversity sets designed on the basis of the maximum common substructure principle, 42 fragments containing carboxyl and amine groups, known ChEMBL targets, library of pharmacologically active compounds, Selleck library (comprising of FDA-approved drugs), and natural product-derived scaffolds from the AnalytiCon Discovery GmBH. These compounds were dissolved in dimethyl sulfoxide (DMSO) to prepare 10 mM stock solution and added 30 nL to each assay plate using an acoustic dispenser.
Description of Libraries of Compounds Included in the HTS
FDA, Food and Drug Administration; HTS, high-throughput screening.
Preparation of cell homogenates and assessing their DIO activities
Cell homogenates from HEK293 cells overexpressing a human recombinant DIO1, DIO2, or DIO3 were prepared as previously described. 37 Based on the DIO activity, the cell homogenates were further diluted to prepare a stock enzyme solution resulting in a consistent and stable assay readout, that is, a robust signal-to-background ratio in the SK reaction (dOD20 mins ∼ 1.2).
Nonradioactive DIO activity measurement assay based on SK reaction
Measurements of DIO activity were carried out as previously described by Renko et al. 37 In brief, cell homogenates containing DIO1, 2, or 3 were incubated along with their preferred iodothyronine substrate and cofactor for 2–4 hours. The released iodide from the reaction was eluted via DOWEX W50-X2 column centrifugation at 200 × g and quantified with SK reaction. The SK reaction is based on a colorimetric reduction of cerium IV with an arsenite solution and is catalyzed by iodide in a concentration-dependent manner. The color changes in this catalytic reaction, from yellow to colorless, are captured as the difference in absorbance at 415 nm in 20 minutes (dOD20 mins) from the start of the assay, allowing iodide quantification and measurement of specific DIO activities. Specific assay parameters for the three isoenzymes differ based on the preferred substrate, reaction pH, incubation conditions, reagent concentrations, and plate formats and are described in Table 2.
Varying Assay Parameters for DIO Activity Measurement Depending on the Plate Format and DIO Isoenzyme
The cell homogenate overexpressing the respective DIO isoenzyme is incubated with a substrate mix that is prepared in 0.1M KPi buffer adjusted to the respective assay pH. Final assay concentrations of components per well are listed.
DIO, deiodinase type; DTT, dithiothreitol; TH, thyroid hormone.
DIO1-inhibition assessment using a high-throughput screen in 384-well plate
The DIO1-inhibition assessment in a 384-well plate format was carried out by incubating DIO1 homogenate with its preferred assay substrate rT3 and cofactor dithiothreitol (DTT) in the presence or absence of test compounds (Table 2). A semiautomatic approach was implemented where a pipetting robot (Biomek Workstation; Beckman Coulter) was used to ensure a consistent addition of the cell homogenate and the substrate solution to each well of the 384-well plate. Harnessing the power of the high throughput of 384 well plates, we screened a total of 69,344 compounds at a single concentration of 10 µM. On each plate, a 100% activity control (10 µg DIO1 homogenate/well, no inhibitor; Table 2) and 0% activity control (ddH2O) for DIO1-inhibitor identification and quality control were added. The released iodide was quantified with SK reaction as described above. 36 We tested 352 compounds on every 384-well assay plate and calculated the % activity for every compound by comparing the absorbance signal after 20 minutes from the compound-treated samples to the median absorbances of the 100% DIO1 activity and 0% activity controls present on the same assay plate. To calculate the z-score, the median and median absolute deviation of the compound-treated samples on the same assay plate were used. The top hits of the primary screen were further assessed by concentration–response curves. For these concentration–response curves, a 2-fold serial dilution in DMSO was prepared, and nine concentrations in total were transferred to the assay plates.
DIO1-inhibition assessment in 96-well plate format
For the manual verification of the DIO1 hits, 20 drugs were sourced from FMP and were assessed by mass spectrometry for their stability. Pranlukast (FMP405931) and FMP407208 were sourced from Selleck Chemicals GmbH, and edaravone (FMP405651), FMP404262, and FMP407222 were sourced from Merck Chemicals GmbH. We measured the effects of these compounds on DIO1 activity with the DIO1 assay in a 96-well plate format as described previously (Table 2). 36 PTU acted as a negative control (100% inhibition control) in every experiment. We calculated % DIO1 activity by normalizing the sample dOD20 mins to the mean dOD20 mins of the 100% activity control.
Potency and DIO1-selectivity assessment of the DIO1-inhibitors
The potency of the DIO1-inhibitors was assessed in 96-well format by incubating the DIO1-containing cell homogenate with varying concentrations of the compound. For every compound, a 3-fold serial dilution, spanning final assay concentrations from 0.5 nM up to 20 µM, was prepared in 2% DMSO solution. These concentrations were incubated with the DIO1-overexpressing homogenate and the substrate mix (Table 2). The DIO1 activity in the presence of varying concentrations of a compound was determined as described above and potency was determined by calculating IC50 values for the compound using nonlinear regression. For compounds available in larger quantities, two additional concentrations of 100 µM and 200 µM were added to the potency and DIO1-isoenzyme selectivity assessment.
To determine the DIO1-selectivity of the inhibitor, the potency of the compound in inhibiting DIO2 and DIO3 was assessed in a concentration-dependent manner, as described for DIO1-potency assessment (Table 2). The experiments for DIO1-potency and isoenzyme selectivity assessment were run once with three technical replicates for each condition unless mentioned otherwise.
Evaluation of DIO1-inhibitory potential of selected compounds in intact cells
To evaluate the effects of DIO1-inhibitors in intact cells, we seeded 40,000 hrDIO1 overexpressing HEK293 cells per well in a 96-well plate and treated them with 10 µM DIO1-inhibitor in Dubecco's modified Eagle medium (DMEM) media supplemented with 0.25% bovine serum albumine (BSA) and 10 µM rT3 for 24 hours. Untreated cells acted as 100% activity controls and 10 µM PTU-treated cells acted as 0% activity control. Supernatant was transferred to a DOWEX column where the released iodide was eluted by centrifugation and quantified using the SK reaction as described above. The response from compound-treated cells is normalized to the 100% and 0% activity control and represented as % DIO1 activity. We also assessed two DIO1-inhibitors for their concentration–response by treating the cells with a 2-fold serial dilution from 10 µM final assay concentration up to 0.16 µM. The experiments were run independently on three different days with three technical replicates on each plate.
Cytotoxicity assessment
Cytotoxicity was assessed for a single concentration (10 µM) of newly identified DIO1 inhibitors on hrDIO1 overexpressing HEK293 cells using an MTT assay (Merck). In brief, cells were seeded in a 96-well plate and treated in triplicates with a 10-µM concentration of DIO1 inhibitors for 24 hours. Solvent control (0.1% DMSO) and a cytotoxicity control (10% DMSO) were added to the plates. Measurement of cytotoxicity was done following the MTT assay protocol, 43 and viability in % for every compound was calculated by normalizing to the solvent control. The solvent control did not impact the cell viability and was the designated signal control representing 100% cell viability. We considered the compounds to be noncytotoxic if the sample absorbance was not beyond 2*SD (standard deviation) of the 100% viability control and mildly cytotoxic if it was 2–4 times the SD of the 100% viability control.
HTS quality parameters and statistical analysis
The “drc” package of R statistical software (version 4.4.1) was used to analyze the HTS concentration–response data. Primary screening data were analyzed using in-house software for converting the plate reader result files into a standard format and Konstanz-Information Miner “KNIME” (version 5.3.1) for hit selection and generating the screening reports. Graphpad Prism (version 9.3.1 for Windows; GraphPad Software, San Diego, CA) was used for the graphical representation of the data. In the HTS assays, the assay quality for every plate was assessed using Z’-factor as a statistical measure of assay quality. 44
To identify primary hits and false positives in the HTS, relative activity and z-scores were calculated for every compound. Eleven plates were duplicated and measured to analyze the assay repeatability with a Bland–Altmann plot. 45 DIO1-IC50 for the concentration–response curves in the HTS were performed using the four-parameter log-logistic function. For the DIO1-, DIO2-, and DIO3-potency assessment, IC50 values for inhibiting DIO isoenzymes were determined from the concentration–response curves using a nonlinear regression model available in Graphpad Prism (ver. 9.3.1). All data are represented as mean ± SD of three independent replicates unless stated otherwise. The number of plates run for each assay is described in the respective method or the results.
Results
A single-concentration HTS assay identifies 436 potential DIO1-inhibitors
Currently, PTU is the only DIO1-selective inhibitor that is available for clinical applications and in vitro discrimination between DIO isoenzyme-catalyzed reactions. Furthermore, a systemic and unbiased screening of small molecular weight compounds has not yet been conducted. Given the clinical interest in DIO1-inhibitory compounds in replacing iopanoic acid to treat acute thyrotoxicosis and their potential use of DIO1-inhibitors in treating Graves’ hyperthyroidism, we decided to screen low-molecular-weight compounds belonging to various chemical libraries for their DIO1-inhibitory potential. In total, 69,344 compounds were screened at a single concentration of 10 µM. We used a modified version of the nonradioactive DIO1 assay 36,37 evolved to 384-well plate format and tested 352 compounds on every plate.
To assess the statistical assay quality and robustness of the DIO1-HTS assay, the Z’-factor was calculated for each plate. 44,46 The Z’-factor acceptance criterion was set to >0.5 and assay plates failing this criterion were repeated or excluded from hit selection. The HTS assays resulted in a consistent and robust performance in the 384-well plate format with a median Z’-factor of 0.83. A good assay repeatability was observed, evident from the lack of systematic deviation between the replicates as seen in the Bland–Altman plot (Supplementary Fig. S1).
With the assurance of robust assay performance, we calculated the percentage of DIO1 inhibition for each compound tested in primary screening at 10 µM concentration and plotted this in descending order (Fig. 1A). By applying stringent criteria (% DIO1 inhibition >25% and z-score >4), a total of 436 compounds were identified as primary hits for DIO1 inhibition (red squares, Fig. 1A). These primary hits also include the previously identified DIO1-inhibitors such as PTU, aurothioglucose, phloretin, and genistein (Fig. 1a, black squares). In our HTS assay, several compounds were repeatedly tested as a part of multiple compound libraries and acted as interassay control. For example, genistein was added to the HTS assay plates on three separate plates, as it belonged to three different compound libraries. In each instance, Genistein got flagged as a potential hit as it resulted in >40% DIO1-inhibition. This finding suggests a robust interassay performance of the HTS assay.
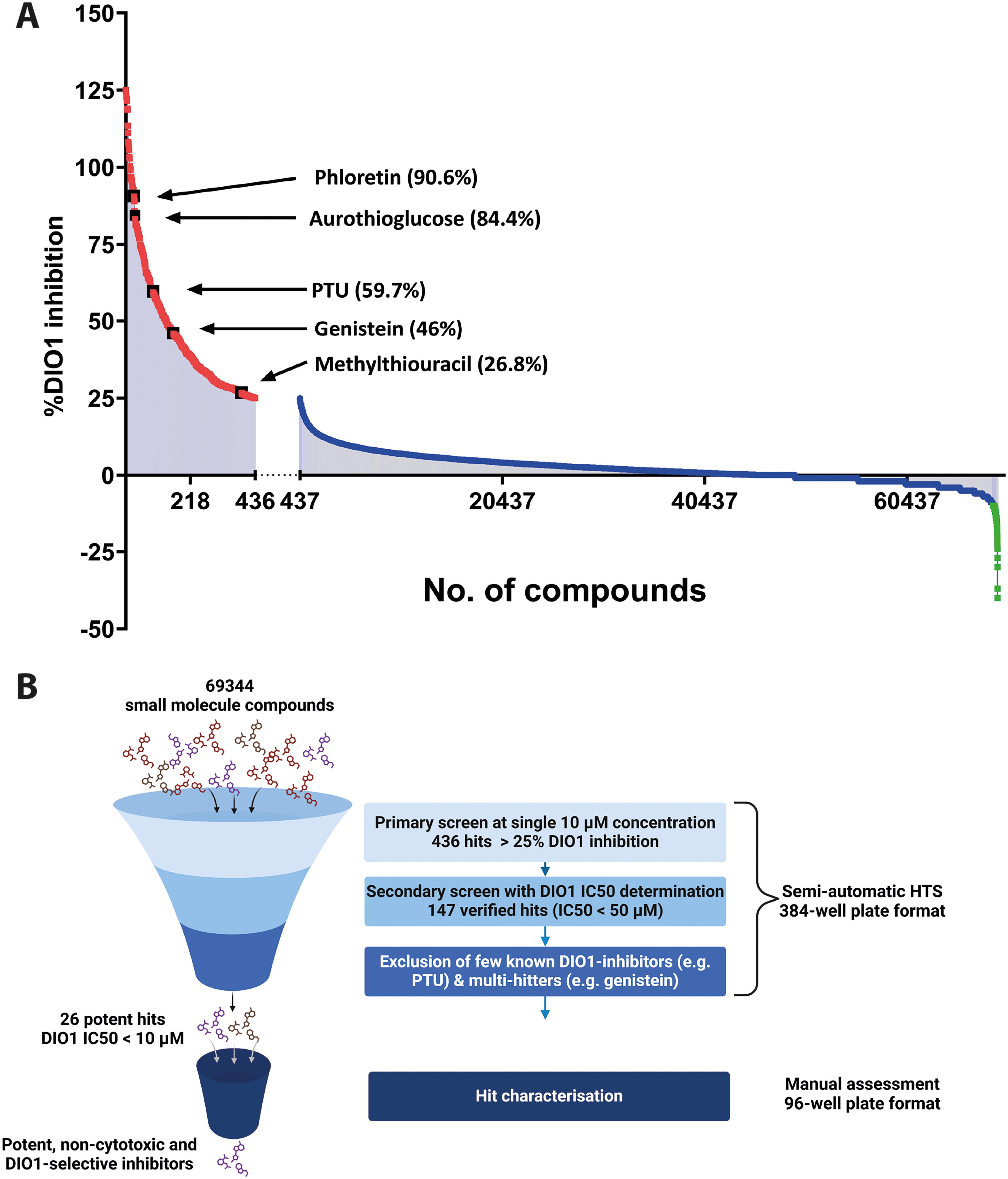
While we did not observe a significant impact of 68,468 compounds on DIO1 activity (Fig. 1a, blue squares), the remaining 440 compounds resulted in a relative DIO1 activity higher than the 100% activity control samples (Fig. 1A, green squares), indicating either assay interference in the SK reaction (e.g., by containing iodide as counter-ion in some preparations or covalently bound) or increase in DIO1 activity.
26 hits shortlisted for manual characterization based on a thorough selection criterion
Following the primary HTS screen that flagged 436 (<1%) compounds as potential DIO1-inhibitors, a secondary screen was employed to exclude false positives, to verify the true-positive hits, and to characterize them regarding potency. In this secondary screen, which included top 298 hits of the primary screen, the respective compounds were tested by preparing a 2-fold dilution series covering a concentration range from 0.1 to 50 µM with technical duplicates for each condition. Concentration–response curves for each compound were generated along with the determination of IC50 and Hill-coefficient values (Supplementary Table S1). To verify a hit as a true DIO1 inhibitor, the overall curve fit and a minimum response difference of 25% between the highest (50 µM) and the lowest (0.1 µM) concentrations were considered.
Out of the 298 chosen hits, 151 compounds were excluded as false positives, and 147 compounds exhibited a robust sigmoidal concentration–response curve with IC50 values <50 µM. From the remaining 147 compounds, known DIO1-inhibitors, and compounds such as genistein, which are known to target multiple cellular processes in a biological system, 47 were excluded from further consideration. Finally, 19 compounds were selected for their high potency (DIO1-IC50 < 1 µM), along with seven additional compounds with their DIO1-IC50 between 1 and 10 µM. These 26 shortlisted hits (DIO1-IC50 < 10 µM) included preclinical and clinical drug candidates, as well as other small molecules with more complex structures. They underwent a thorough characterization process to assess their potency, selectivity for DIO isoenzymes, and effects in intact cells. Figure 1B depicts a schematic of our selection process and criteria detailing our efforts in identifying potent, DIO1-selective inhibitors.
Potency and isoenzyme selectivity assessment revealed 15 compounds to be highly potent and DIO1-selective inhibitors
Shortlisted inhibitors were experimentally validated for isoenzyme-selectivity and potency confirmation for DIO1, by applying the DIO assays for all three isoenzymes (Table 2) at varying inhibitor concentrations (0.5 nM–20 µM) in 96-well assay format.
For every compound, we plotted a concentration–response curve for all DIO isoenzymes and determined its DIO-IC50 value to assess its potency. DIO-IC50 values for each isoenzyme were calculated with high accuracy covering a concentration range between 0.5 nM and 20 µM. For six compounds that were available in larger quantities, we tested two additional concentrations (100 and 200 µM) to obtain data comparable to the DIO1 screening data generated by the EPA ToxCast project, USA. 41
Figure 2 depicts the results of the potency and isoenzyme selectivity assessment for four compounds out of the 26 shortlisted hits. Compound FMP405302 or phloretin (Fig. 2A), a known DIO1-inhibitor, showed potent and DIO1-selective inhibition, with an IC50 value of 0.7 µM, without inhibiting DIO2 and DIO3 at the highest tested concentrations. On the contrary, compound FMP407222 (Fig. 2B) completely inhibited all 3 DIO-isoenzymes at the very high concentration of 200 µM, but the potency of inhibition was several fold higher for DIO1 than for DIO2 and DIO3. We determined the IC50 of FMP407222 for DIO1 to be 9 nM, demonstrating significantly greater potency compared with its IC50 for DIO3 (∼3.7 µM, over 400-fold higher) and DIO2 (∼20.3 µM, over 2,000-fold higher). Compounds FMP203345 and FMP205373 tended toward a nonselective DIO inhibition at concentrations higher than 10 µM (Fig. 2C, D) and therefore were excluded from further testing.
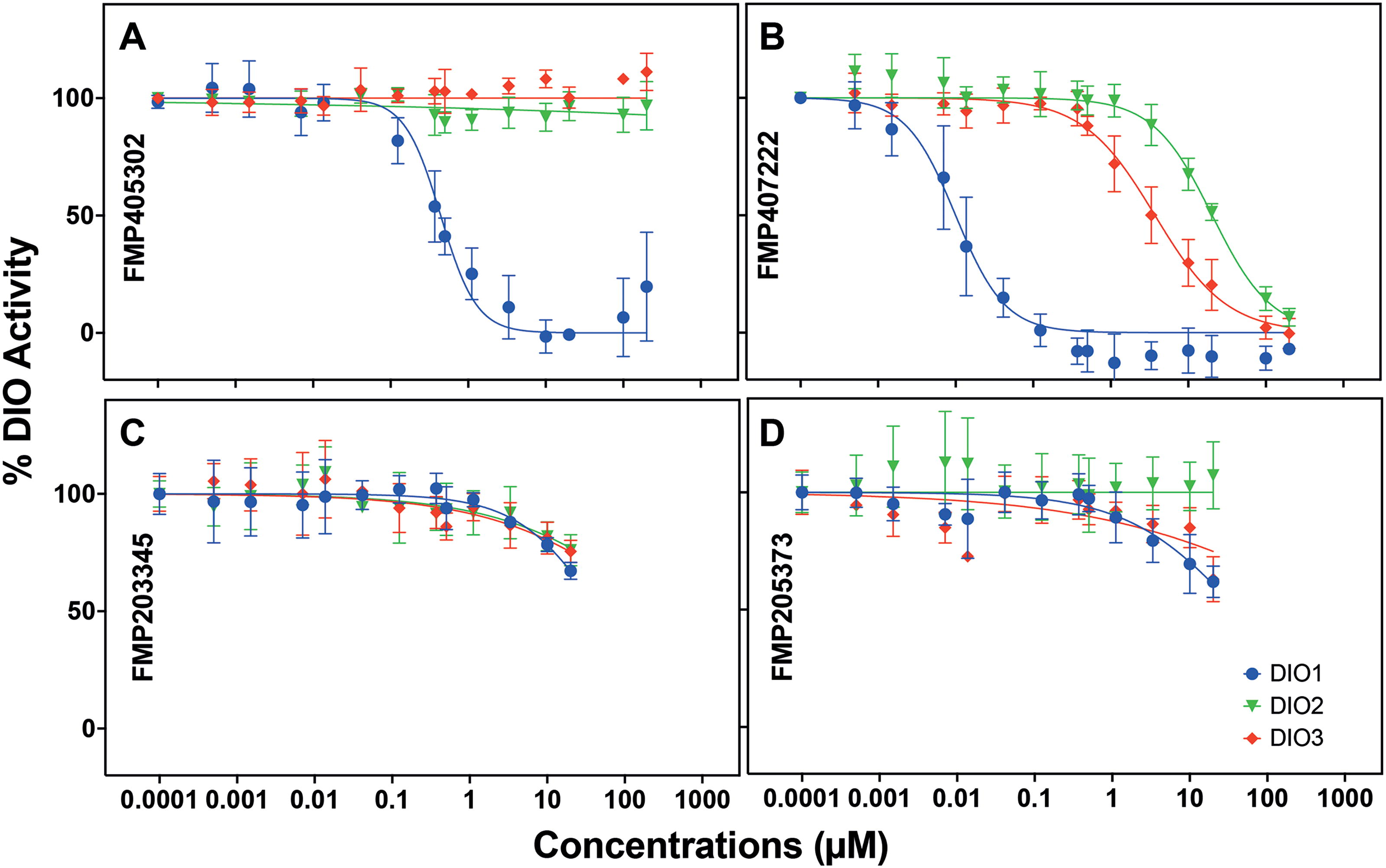
Concentration–response curves for DIO activity inhibition by four compounds, tested at concentrations ranging from 0.5 nM up to 200 µM for all three DIO isoenzymes. % DIO activity is calculated by normalizing the response to the response of vehicle control-treated samples and plotted against respective concentrations. The IC50s for each compound were calculated by curve-fitting using a nonlinear regression model. Compound
The other shortlisted compounds also showed a DIO1-selective inhibition without inhibiting DIO2 or DIO3. Of the 26 compounds, three compounds including FMP203345 and FMP205373 have a DIO1 IC50 value >10 µM (Fig. 2C, D), eight compounds show a DIO1-IC50 value between 1 and 10 µM and 15 compounds, including phloretin and FMP407222 (Fig. 2A, B), were highly potent and DIO1 selective with their IC50 values in the nanomolar range (DIO1 IC50 < 1 µM). For all 26 compounds, the IC50 values for inhibiting DIO1, DIO2, and DIO3 are summarized in Table 3, and the concentration response curves for the remaining 22 compounds can be found in Supplementary Figure S2.
Characterization Summary for the 26 Shortlisted Compounds
Potency and DIO1-selectivity of the compounds is assessed by comparing the IC50 values for DIO1, DIO2, and DIO3 inhibition. PTU, a DIO1-selective inhibitor, resulting in a DIO1-IC50 value of ∼1.5 µM, and Xanthohumol, a pan-DIO inhibitor, results in IC50 value ∼ 1.5–3 µM for all three DIOs.
PTU, propylthiouracil.
To consider these shortlisted compounds for their potential use as pharmaceutical agents, we measured and compared the cell viability upon the compound treatment with an MTT assay. The % viability for the 26 shortlisted compounds is summarized in Table 3. With 24 hours of compound treatment at 10 µM, we observed that 12 compounds did not result in any significant cytotoxic effects in hrDIO1 overexpressing HEK293 cells, and the remaining 14 compounds exhibited only mild cytotoxicity. Figure 3 shows the % viability of seven compounds compared to the solvent control and the cytotoxicity assessment for the remaining compounds is summarized in Supplementary Figure S3. Based on these results, we did not exclude any compound from further consideration.

Representative example highlighting the effects of 24-hour treatment of 7 DIO1-inhibitors on the viability of hrDIO1 overexpressing HEK293 cells at a 10 µM concentration as measured by MTT assay. Data are expressed as mean % viability ± SD (n = 1). DIO1, deiodinase type I.
Summarizing the results of our HTS efforts, we unbiasedly screened 69,344 compounds for their DIO1 inhibitory potential, and we found 15 novel compounds highly potent and selective for DIO1 inhibition.
Eight compounds out of 13 are capable of inhibiting DIO1 in intact cells
Our assessment of DIO1-inhibition using homogenates from DIO1 overexpressing cells can only generate data on a biochemical level, omitting fundamental cellular functions. Evaluation of potential cellular impact of DIO1 inhibition is crucial for interpretation of local cellular TH availability and any resulting downstream physiological processes. In physiologically intact cells, compound efficacy in affecting a biological process may vary due to cellular uptake, stoichiometric requirements, and compound metabolism. To answer whether a compound can enter a cell and interfere with the deiodination reaction, we modified our DIO1 assay to evaluate DIO1 inhibition in intact cells. Taking advantage of the fact that the cells expel the iodide that is released in the deiodination process, we measured the released iodide in the cell-culture supernatant with the SK reaction. The quantity of iodide measured in the supernatant allowed us to determine how functional the DIO1 enzyme in an intact cell system is, and whether the inhibitor treatment has any effect on the deiodination process. The assay schematic is represented in Figure 4A.

A 24-hour treatment of cells with 13 shortlisted DIO1-inhibitors demonstrated that not all compounds could inhibit DIO1 in a physiologically intact cell system using 10 µM rT3 as substrate. With eight out of the 13 tested compounds, the iodide found in the cell-culture supernatants was quite low and comparable with the PTU-treated cells. These results indicated that eight compounds were able to enter the cells and inhibit the DIO1 blocking the iodide release into the supernatant. The remaining compounds were not able to inhibit the DIO1 activity (Fig. 4B) under the given conditions.
To delineate whether the DIO1 inhibitors also caused a differential inhibition of DIO1 in an intact cell system, we incubated the cells in a 96-well plate with varying concentrations of phloretin and FMP407222. After 24 hours, we measured the released iodide from the cell-culture supernatants. Both phloretin and FMP407222 resulted in a concentration-dependent decline in the % DIO activity, as shown in Figure 4C. For phloretin, the DIO1-IC50 determined in intact cells and cell homogenates is ∼0.4 and 0.7 µM. For FMP407222, the difference in the DIO1-IC50s determined in intact cells and cell homogenates is quite stark, with DIO1-IC50 in intact cells being ∼0.4 µM and DIO1-IC50 in cell homogenates being 0.009 µM. To summarize, we have demonstrated the DIO1-inhibition in intact cells by eight newly identified and highly potent DIO1-selective inhibitors, thereby providing the first indication of their potential use as drugs or reference compounds for DIO1-selective inhibition.
Discussion
We present a successful and comprehensive screening of 69,344 compounds with a high-throughput technology that led us to identify 15 novel, potent, and selective inhibitors of human DIO1. We have also demonstrated that 8 out of these 15 inhibitors successfully inhibit DIO1 in cells.
Adapting the nonradioactive SK-based DIO1 assay 36 to a 96-well plate format, various research efforts were targeted toward identifying (environmental) toxicants disrupting the TH system by inhibiting DIO1. 39 –41 To address the gap of an unbiased, comprehensive screening effort to identify DIO1-selective inhibitors, we opted to screen a large and diverse set of low-molecular-weight compound (<1000 Da) libraries. These low-molecular-weight compounds mimic the size of THs and comprise of small molecule low molecular weight (<1000 daltons) organic compounds (SMOLs), compounds used as tools or probes, drugs approved for medical use, pharmacologically active compounds, etc. Screening for DIO1 inhibitory effects at a 10 µM concentration enabled us to compete with the 10 µM rT3 substrate concentration, and to identify compounds as potent as, or more potent than, the widely used DIO1 inhibitor PTU (IC50 ∼ 1.5 µM) without generating potential interference with cell membrane integrity and cell viability in cellular applications. Our success in optimizing the DIO1 assay to run in a 384-well plate format allowed us to screen a total of 69,344 compounds and supported our choice of an unbiased screening approach.
The semiautomated screening assays in the 384-well plate format performed in a robust and reproducible manner. This is evident considering the Z’-factor >0.5 of the assay plates, low variability in signal controls and similar signals for the replicated chemicals, for example, genistein. The successful flagging of previously identified DIO1-inhibitors such as PTU and other thiouracils, aurothioglucose, and phloretin, a well-known potent plant-derived DIO1 inhibitor 48 further highlights the robustness of the HTS assay. After the identification of hits through a primary screen at a singular 10 µM concentration, we verified a total of 147 hits through a verification screen and excluded several false-positive compounds, which did not exhibit DIO1 inhibition up to concentrations of 50 µM. When comparing our DIO1-HTS results with previous publications that aimed to test ToxCast phase 1, phase 2 and e1K libraries for DIO-inhibition, 40,41 we found 83 compounds in our screening compound libraries that were also present in the ToxCast phase 1, phase 2, and e1K libraries, and these 83 compounds included compounds such as sulfasalazine, PTU, and methylthiouracil. These compounds were flagged as DIO1-inhibitors in both studies, including ours. However, the different concentration ranges employed for potency assessment, different sources of enzymatic activity and the increased assay throughput (96 vs. 384-well plates), combined with the specific focus of our inhibitor testing, makes a direct comparison difficult. Two compounds, temephos, and 2-chloro-N-phenylacetamide, that exhibited a DIO1-selective inhibition with IC50 values below 20 µM, 41 were not identified as potential DIO1-hits in our initial screens and hence were not pursued.
The SK-based HTS assay offers several advantages in terms of reproducibility and robustness over a defined concentration range for every tested compound. This is especially important when minimizing any false-negative hits, but it is quite likely to have missed few inhibitory compounds on account of low compound efficacy and inaccuracies of compound addition to the 384-well plates. Another significant advantage offered by the SK-based HTS assays is the versatility, where the assay can be modified and applied to assess the functioning of other critical targets of the TH-axis such as iodotyrosine dehalogenase 1 (DEHAL1), 49,50 transmembrane transporters, 51 –54 and potentially also TH distribution proteins that handle iodinated compounds.
The thorough characterization of 26 compounds for their potency and DIO1-selectivity revealed that 23 compounds were found to be DIO1-selective. Out of the three compounds that show a nonselective inhibition, FMP407222 is one of the most potent inhibitors of DIO1 found in our study and exhibits a huge difference in potency for the three isoenzymes (Fig. 2B). These compounds, with an exception of FMP407222, may represent a class of pan-DIO inhibitors, such as xanthohumol (a known pan-DIO inhibitor with IC50 ∼ 2 µM for all DIOs). 37 These pan-DIO inhibitors have the potential to significantly alter TH-status, either in specific tissues or throughout the entire body. Thus, careful monitoring of such inhibitors is required to limit their exposure to humans and/or other organisms. The DIO1-selectivity observed for the 23 remaining hits may be explained by the two-substrate ping-pong mechanism of DIO1 when compared with the sequential deiodination by DIO2 and DIO3. However, three compounds with DIO1-selective inhibition (Table 3), namely phloretin, edaravone, and pranlukast, depicted no structural similarities or other motifs common with the widely used DIO1-inhibitor PTU and iopanoic acid, a competitive DIO1-substrate, 36 that may confer and explain this observed isoenzyme-selective inhibition (Fig. 5). Nevertheless, we identified 15 compounds that were highly potent with their DIO1-IC50s in nanomolar range, when compared with the DIO1-IC50 of PTU (∼4 µM). With such potent and selective inhibition of DIO1, we also observed that these shortlisted compounds did not cause any major cytotoxic effects in HEK293 cells with DIO1-overexpression (Fig. 3 and Supplementary Fig. S3). These findings ensure that these inhibitors, with further characterization and development, hold a high potential for their use as lead structures for the development of pharmaceutical agents in modulating local TH availability, particularly when targeted to tissues with high DIO1 activity such as the liver, kidneys, and thyroid gland.
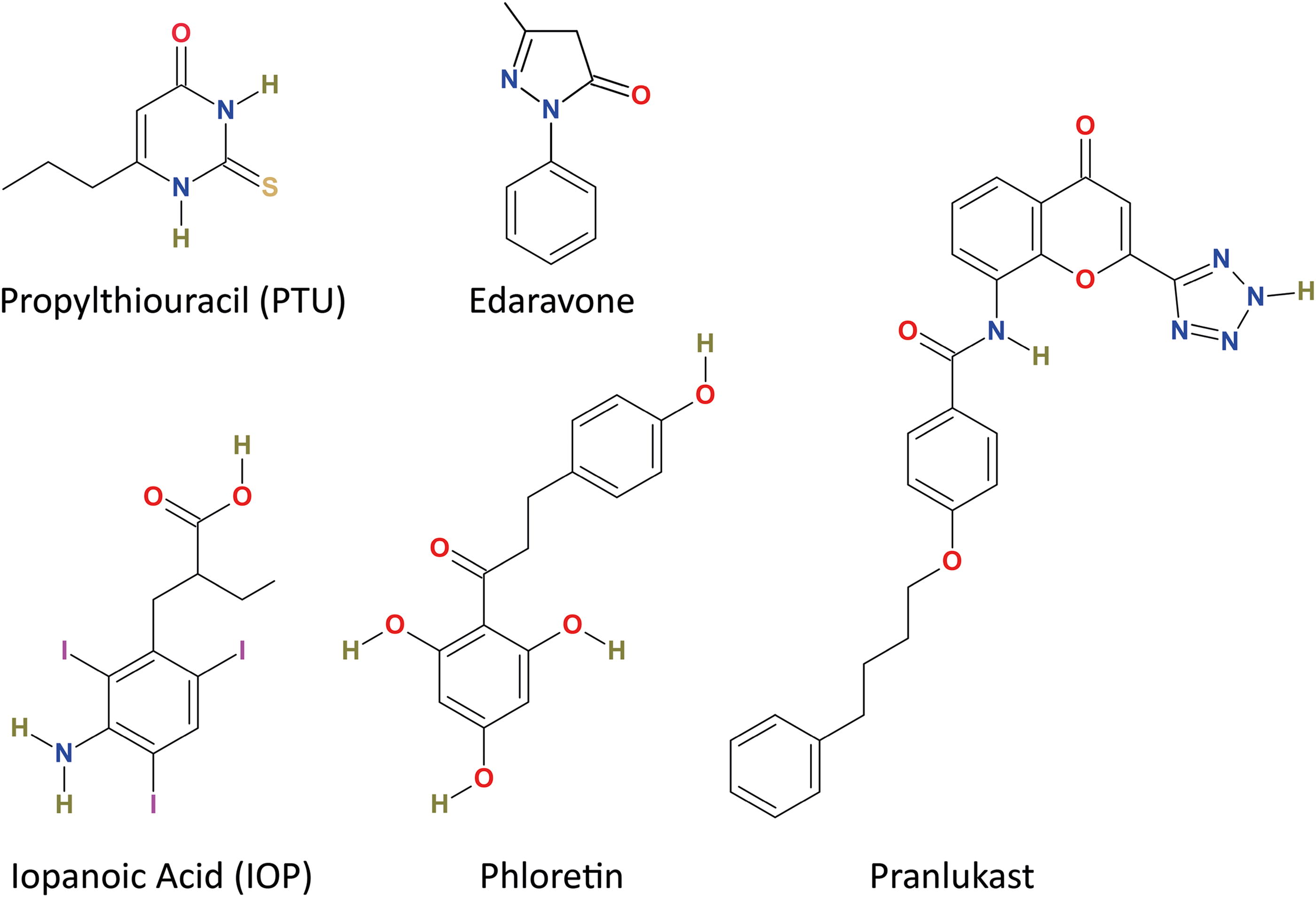
Chemical structures of widely used DIO1-inhibitors, propylthiouracil and iopanoic acid, along with a previously identified DIO1-selective inhibitor, phloretin and two drugs, edaravone and pranlukast, identified in this study for their DIO1-selective inhibition.
The application of these novel and highly potent DIO1-inhibitors in mechanistic studies with enzyme test procedures better suited for this purpose could elucidate the differences in the reaction mechanism of DIO1 in comparison with DIO2 and DIO3. 55,56 The DIO1-inhibitors would allow us to develop structure–activity relationships (SAR) for DIO1, complementing the SAR analysis using DIO1 substrates. 57 While there are some initial insights into the active site and mechanisms of DIO1, the enzyme structure remains unknown, and studying the DIO1 inhibition via different types of inhibitors could provide further information on the active site structure and essential amino acid residues possibly involved in the catalytic mechanism of DIO1. 58
We have, for the first time, described a successful application of SK reaction to measure the iodide content of the cell-culture medium after intracellular deiodination. This quantification of iodide expelled by the cells into the cell-culture medium allowed us to assess the effects of DIO1-inhibitors on the process of intracellular deiodination. While eight compounds out of the 13 tested were able to enter the DIO1-overexpressing HEK293 cell and inhibit DIO1, we found five compounds that were unable to do so. Interestingly, we observed that the compounds either had a strong inhibitory effect (>90% inhibition) on the deiodination process or had no effect under the chosen assay conditions. Furthermore, concentration-dependent DIO1-inhibition was observed for both phloretin and FMP407222, when tested at varying concentrations in intact cells. The difference in IC50 values observed when cell homogenate is treated to when intact cells are treated with a DIO1-inhibitor could be attributed to barrier to cell entry, higher affinity for other cellular processes, or a quick cellular metabolism of the drug. While we opted for HEK293 cells that overexpressed DIO1 to study the DIO1-inhibitory effects of the compounds, detailed experiments will be required to elucidate the effects of these compounds in complex cellular or organoid models with physiologically relevant levels of endogenous DIO1 activity. These experiments will help distinguish the inhibitor’s impact on cytotoxicity and other cellular processes from details of the inhibitory reaction mechanism of the long-lived DIO1 (t1/2 > 12 hours) enzyme.
In our screening efforts, we found two drugs, edaravone (FMP405651) and pranlukast (FMP405931), which are approved for medical use (Fig. 5). Edaravone is a free-radical scavenger used in the treatment of amyotrophic lateral sclerosis, and pranlukast is a leukotriene receptor-1 antagonist used for alleviating asthmatic symptoms. Recently, edaravone has been shown to ameliorate the adverse effects of hepatic lipid accumulation and western diet by alleviating the symptoms of metabolic dysfunction-associated steatohepatitis in mice via the activation of AMPK. 59 Pranlukast exerts varying effects on cytochrome P450. On one hand, pranlukast is metabolized by Cyp3A4, and on the other hand, it has also been reported to reversibly inhibit other enzymes of the Cytochrome P450 family (CYP2C8 and CYP2C9). 60 While we observed a DIO1-inhibition comparable to the potency of PTU on testing with DIO1-containing cell homogenates, we found that edaravone, but not pranlukast, was able to inhibit the DIO1 in our experiments with the used intact cell systems (Fig. 4B), highlighting a differential intracellular response. These drugs are approved for their medical use, but there are no available reports on either of the drugs characterizing their effects on TH system in patients.
A target-specific inhibition of DIO1 would allow us to locally modulate TH concentrations in a reversible and concentration-dependent manner, in contrast to the genetic knock-down or knockout approach. This local modulation holds high potential in deciphering the role of a particular tissue in TH economy. Furthermore, novel strategies can be developed to combat various disorders that are either directly TH-related or present through local alterations of TH status. In hyperthyroid states, such as Graves’ disease, or autonomous thyroid adenoma, antithyroid drugs such as PTU are used to inhibit TH synthesis by blocking TPO. While PTU also blocks DIO1, a more potent DIO1-inhibitor would be extremely beneficial in reducing high T3 levels and alleviating the effects of hyperthyroidism. Furthermore, DIO1-selective inhibitors could be critical in treating the life-threatening thyroid storm, where iopanoic acid has been successfully used. Administration of iopanoic acid, a known DIO1-substrate, 36 also results in a long-term persistence of the iodinated agent in vivo releasing iodide, 61 which renders it unattractive for use in the treatment of thyroid storm.
To conclude, our unbiased HTS approach successfully identified 15 novel, potent, and selective inhibitors of DIO1 from an extensive library of 69,344 compounds. The in-depth characterization of the inhibitors validated their potency, selectivity, and minimal cytotoxicity in intact cells. While not all compounds were able to intracellularly inhibit DIO1 overexpressed in HEK293 cells, their effects in other cell lines may vary. The possibility of varying organ-specific response only highlights the complexity in translating in vitro findings to functional biological systems. However, we have laid a groundwork for studying the DIO1 structure, SAR, and provided novel tools to study and potentially modulate local TH availability and action by DIO1 inhibition.
Footnotes
Acknowledgments
The authors thank Anja Fischbach, Gabriele Boehm, and Niklas Wiese and the rest of the team at the Institut für Experimentelle Endokrinologie at Charité-Universitätsmedizin Berlin for their support. The authors would also like to acknowledge the role of BioRender in generating figures.
Authors’ Contributions
R.S.: Methodology, investigation, validation, formal analysis, data curation, visualization, and writing—original draft. C.S.: Investigation and data curation. S.K.: Investigation and data curation. M.N.: Data curation, formal analysis, visualization, project planning, and writing—review and editing. J.P.v.K.: Writing—review and editing, conceptualization, and project planning and co-ordination. C.F.: Methodology and data analysis and review. K.R.: Methodology and writing—review and editing. E.K.W.: Methodology, data curation, writing—review and editing, visualization, project administration, supervision, and funding acquisition. J.K.: Data analysis, article revision, project administration, supervision, funding acquisition, conceptualization, and project planning and coordination.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This project receives funding by the Deutsche Forschungsgemeinschaft within the framework of CRC/TR 296 Locotact P16 to J.K. and E.K.W. and by the EU Horizon 2020 program for the ATHENA (Assays for the identification of Thyroid Hormone axis-disrupting chemicals), grant number 825161, to J.K. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the article, or in the decision to publish the results.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1