
Abstract
Members of the Order Chiroptera are the natural reservoirs of lyssaviruses and play an important role in the transmission of rabies to animals and humans. In this present study, the seroprevalence for rabies virus was determined for bats sampled from four southern provinces on the Chinese mainland. A total of 685 bats of 8 species representing 4 families were collected from 10 sites, and were tested by the indirect fluorescent antibody test using fluorescein isothiocyanate (FITC)–conjugated protein A/G mixture and viral neutralization test. Rabies antibody response was only detected from three bat species (Rousettus leschenaulti, Rhinolophus blythi, and Rhinolophus ferrumequinum). The overall rabies seroconversion rate was only 2.2% (15/685). Of the 15 positive sera, 13 (12 fruit bats and 1 insectivorous bat) were indirect fluorescent antibody test positive, and two insectivorous bats were virus neutralization positive when tested by the modified fluorescent antibody viral neutralization test, albeit extremely low. To our knowledge, this is the first published report describing rabies seroprevalences from Chinese bats. These results suggest that bats may play a role in the ecology of lyssaviruses in China, and further surveillance for the presence of lyssaviruses in bats should be undertaken throughout the country and extended to other species.
Introduction
China has 123 species of bats, belonging to 30 genera and 7 families, and are distributed nationwide (Xu et al. 2002). The first bat-associated rabies case was detected in China in 2002, and this greatly increased people's awareness of bat lyssaviruses as potential human disease threats (Tang et al. 2005). Additionally, a few bat lyssavirus reservoir species documented from other countries also exist in China. In this study, the seroprevalence of RABV was investigated in bats collected from Southern Chinese regions known for severe rabies endemics.
Materials and Methods
Reagents, positive serum, cell and virus
Dog anti-RABV hyperimmune serum with a neutralizing antibody titer of 10.26 IU/mL was prepared previously in the author's laboratory at the Institute of Veterinary Sciences (IVS), Academy of Military Medical Sciences, China, and was used as positive control serum. Negative control serum was prepared from a dog that had never been given rabies vaccine, and that was confirmed by fluorescent antibody viral neutralization (FAVN) to be rabies antibody negative. An international anti-RABV antiserum from the OIE Reference Laboratory for Rabies (Nancy, France) was used for the study. Mouse neuroblastoma cells and attenuated RABV strain SRV9 were also from IVS. RABV CVS-11 strain (ATCC: VR 959) and BHK-21 cells (ATTC: CCL-10) were provided by the Veterinary Laboratories Agency (VLA), Weybridge, UK.
FITC-conjugated protein G (FITC–protein G) and protein A were purchased, respectively, from Sigma (St. Louis, MO) and Fluka (Ronkonkoma, NJ). FITC-conjugated anti-rabies monoclonal antibody was purchased from Fujirebio (Malvern, PA). Difco minimum essential medium and fetal bovine serum were purchased from Gibco-BRL (Gaithersburg, MD).
Collection of specimens
From April 2005 through May 2006, blood samples were obtained from the wing veins of 685 apparently healthy bats collected in the following 4 Southern Chinese provinces: Guangdong, Guangxi, Yunnan, and Hainan. Bat species and sites of sample collection were randomly selected based on the proximity of the animals to human residence and the availability of their roosts. Samples were stored in an icebox until serum separation by routine microcentrifugation at 12,000 rpm for 10 min. The 685 serum samples analyzed represented 8 bat species. Of course, one species was a fruit bat, which was collected from six sites in four provinces. The other seven species were insectivorous, and collected from five sites in Guangdong Province (see Fig. 1 and Table 1). Serum samples of volumes >20 μL were tested by indirect fluorescent antibody test (iFAT), followed by confirmation with conventional FAVN (Trimarchi et al. 1996) when sufficient sample volumes allowed. Some bat sera with volumes >50 μL were assayed at VLA by previously published modified FAVN (mFAVN) method (Brookes et al. 2005).

The map of China showing eight bat collection cities in Yunnan (A), Guangxi (B), Guangdong (C), and Hainan (D) provinces. Cities are Funing (FN), Nanning (NN), Ningming (NM), Yunan (YA), Luoding (LD), Wengyuan (WY), Yangchun (YC), and Boao (BA) as acronyms represent.
MS (Mashui), KD (Kongdong), and TS (Tanshui) are three townships of Yangchun City, and other acronyms are refered to those in Figure 1.
Four species of 57 insectivorous bats captured for sampling were all from a cave in TS.
iFAT
Mouse neuroblastoma cells were grown to 3 × 105 cells/mL using Difco minimum essential medium with 2% fetal bovine serum, and 100 μL aliquots were added to each well of 96-well plates and combined with 100 μL of RABV SRV9 (300 TCID50) per well. The plates were incubated at 37°C in a humidified 5% CO2 atmosphere for 72 h, allowing the cells to reach 90% confluence. After supernatant removal, the plates were fixed with 80% cold acetone at −20°C for 30 min.
To permit optimal sensitivity, iFAT was conducted using FITC–protein A/G mixture as in place of a second antibody. Since bat antiserum (rabies positive and negative) is not available for use as controls, dog serum (rabies positive and negative) was used to validate the use of FITC–protein A/G. The sensitivity of iFAT using rabies antibody-positive dog serum was established by the observation that specific green fluorescence in the cytoplasm of RABV-infected cells could be seen even at 1:256 (Fig. 2A), a dilution corresponding to a neutralizing antibody titer of 0.04 IU/mL.
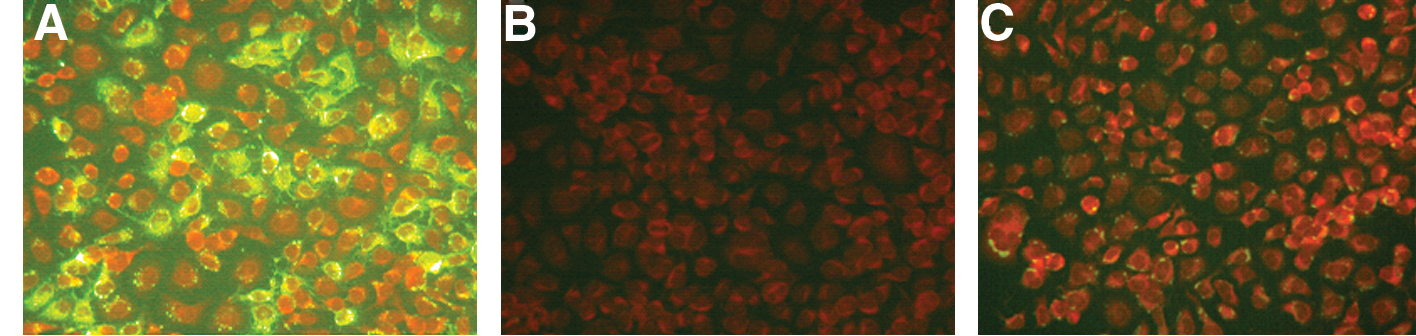
The indirect fluorescent antibody test. Dog anti-rabies virus (RABV) hyperimmune serum (1:128) was added to RABV-infected cells as a positive control (
In the test, the serum samples were complement inactivated at 56°C for 30 min and then diluted with 0.1 mol/L phosphate-buffered saline (PBS) (pH 7.2). Each bat serum was diluted 4- and 8-fold with each test using 32- and 128-fold diluted dog anti-RABV serum as a positive control. FITC–protein A and FITC–protein G were diluted separately in PBS according to the manufacturer's instructions and then mixed before use. The mixture contained 0.01% Evans Blue, and the working dilutions of FITC–protein A and FITC–protein G were 1:16 and 1:64, respectively.
The fixed plates were washed four times with PBS containing 0.05% Tween-20 (PBST), then serum was added (50 μL/well), and incubation at 37°C was continued for 90 min. Plates were washed with PBST four times, and FITC–protein A/G working solution (50 μL/well) was added, with incubation for another 90 min. After five washes, plates were observed and photographed using an Axioskop-40 fluorescence inverted microscope (Carl Zeiss, Jena, Germany).
FAVN test
If quantity permitted, iFAT-positive samples from R. leschenaulti were confirmed by conventional FAVN according to the standard World Health Organization protocol (Trimarchi et al. 1996). Since the FAVN method requires a minimum of 200 μL serum and the small body sizes of insectivorous bats limited collection volumes, an mFAVN, which reduced the volume of serum to 50 μL was used for these samples. For each mFAVN assay, serum samples were first diluted ninefold with 0.1 mol/L PBS, pH 7.2, then subjected to threefold serial dilution to 1:729. Each dilution was assayed twice, and OIE international standard serum and virus controls were included in each assay.
Results
A total of 685 serum samples were tested by iFAT and/or FAVN, collected from 8 species in 4 families, including R. leschenaulti (n = 514), Rhinolophus ferrumequinum (n = 18), Rhinolophus blythi (n = 74), Hipposideros larvatus (n = 57), Hipposideros bicolor (n = 9), Hipposideros armiger (n = 2), Myotis daubentoni (n = 1), and Scotophilus heathii (n = 10). Of the 685 samples tested, 15 were positive; that is, a 2.2% overall seropositive rate (Table 1). The seroprevalence for frugivorous bats was 2.3% (12/514) and that for insectivorous bat sera was 1.8% (3/171). Of the 15 positive sera, 13 (12 fruit bats and 1 insectivorous bat) were iFAT positive, demonstrating clear and discernible cytoplasmic fluorescence without nonspecific staining on RABV-infected cells at both fourfold and eightfold dilutions, although weaker staining than for the positive control sera (see Fig. 2C). In contrast, as expected, these 13 bat sera did not show positive staining on mock-infected cells (data not shown). Other two insectivorous bats tested by the mFAVN demonstrated a neutralizing antibody response of 0.38 IU/mL with a 1:9 starting dilution (samples were not tested by iFAT because of insufficient volumes), albeit the titers were extremely low and below the recommended sensitivity threshold used for canine serum of 0.5 IU/mL. The neutralizing antibody threshold in serum samples from chiropteran species has not yet been established using mFAVN, making the results difficult to interpret. R. leschenaulti, R. blythi, and R. ferrumequinum showed rabies antibody with 2.3% (12/514), 2.7% (2/74), and 5.6% (1/18) positive rates, respectively. With respect to geographical distribution, Yangchun City of Guangdong Province appeared to have wider distribution of infected bats than other cities, as two of its three townships, Kongdong and Tanshui (TS), had seropositive bat colonies. In Tanshui, two of four species collected from the same cave were RABV seropositive (see Table 1).
Of 636 samples tested by iFAT 18 R. leschenaulti sera (3 iFAT positive and 15 iFAT negative) were subjected to FAVN. Results showed that 3 iFAT-positive sera (two from Nanning, Guangxi Province; one from Boao, Hainan Province) had neutralization antibody titers of 0.33, 0.15, and 0.08 IU/mL, while 15 negative sera had no detectable neutralization antibody, showing 100% concordance with iFAT.
Discussion
Bats are the only true flying mammals, and possess the broadest distribution of any mammalian taxa in the world. Research undertaken during the last few decades has shown that bats represent the largest natural reservoir for more than 60 species of zoonotic viruses, with most being highly pathogenic RNA viruses (Wong et al. 2007). As a continuing source of emerging infectious diseases, bats are attracting increasing scientific concern. Clearly, more extensive surveillance needs to be undertaken to prevent spill-over of their pathogens to human and other animal populations.
The antibody neutralization test is a proven method that is commonly used for the serological measurement of lyssavirus antibodies (Arguin et al. 2002, Reynes et al. 2004, Lumlertdacha et al. 2005). This method is not ideal for large numbers of serum samples since it is time consuming and requires high biosafety procedures. Recently, peroxidase-labeled protein A/G has been used in place of a second antibody in an enzyme-linked immunosorbent assay (ELISA) to detect bat serum antibodies, since labeled anti-bat immunoglobulin antibodies are not commercially available. Reynes et al. (2004) developed an indirect ELISA to detect anti-lyssavirus antibody of bat serum using commercial horseradish peroxidase (HRP)–conjugated protein A and protein G (Pierce) as labeled secondary antibodies. Similarly, ELISA using HRP-conjugated protein A/G mixture (Sigma) in place of a second antibody was also applied to detect serum antibodies of different bats against filoviruses (Pourrut et al. 2007, Towner et al. 2007). These studies have shown that protein A/G can effectively bind to bat immunoglobulin. As further evidence, ELISA using HRP-conjugated protein A/G in the detection of Rousettus aegyptiacus serum IgG against Marburg virus provided identical results to those obtained using HRP-conjugated goat sera raised against a cocktail of IgG from several diverse bat species (Towner et al. 2007). The ELISA is a rapid and effective method for testing large numbers of samples, but it requires high-quality specific antigen for plate coating to ensure valid results.
In the present study, the iFAT was developed by modification of the regular FAT using commercial FITC-conjugated protein A/G to detect bat serum antibodies against RABV since FAT serves as the gold standard method recommended by WHO for rabies diagnosis. The validity of the iFAT in our setting was shown using appropriate controls, such as rabies-positive and -negative sera (see results). To further evaluate the broad usefulness of this method, FITC–protein A/G mixture was used to detect 33 FAVN-positive sera (5 humans, 21 dogs, and 7 mice) against rabies and 9 FAVN-negative serum samples (3 for each species). The results demonstrated 100% concordance with FAVN (data not shown). In addition, the FITC–protein A/G mixture was also used to detect mouse and pig serum antibodies raised against baculovirus-expressed Nipah virus N protein and classical swine fever virus on fixed cell monolayers infected with the corresponding virus. The result showed that specific cytoplasmic green fluorescence was clearly visible by microscopy when compared to mock-infected cell controls (images not shown). All these experiments showed the specific binding of FITC–protein A/G to immunoglobulin of human and three mammals. Our results indicate that FITC–protein A/G can be effectively used to highlight bat immunoglobulins and serological evidence of rabies exposure in bats. Neutralization antibody titers from three fruit bats and two insectivorous bats were very low (0.33, 0.15, 0.08, 0.38, and 0.38 IU/mL), but the titers corresponded with weak staining in the iFAT, further confirming the results of the latter. The iFAT requires 20 μL serum sample, a much lower volume than that required for conventional FAVN (200 μL) and mFAVN (50–100 μL), readily obtainable from small insectivorous bats.
Classical RABV is the principal agent responsible for human rabies worldwide, and canids are the most important terrestrial reservoir for rabies transmission. Mainland China has ranked second worldwide in human rabies death each year (Tang et al. 2005). In 2008, 2378 people in China died of rabies (The Ministry of Health of China 2008), and dogs play a pivotal role in its transmission (Tang et al. 2005, Zhang et al. 2006). Outbreaks of human rabies in China are more severe in the south than in the north (Tang et al. 2005). Investigations on the prevalence of RABV infection not only in dogs but also in wild canids, and especially bats roosting in these provinces, will be essential for elucidating the transmission dynamics and epidemiological characteristics of rabies in Southern China. Therefore, four utmost southern provinces, which have abundant bat resources (Xu et al. 2002), were selected for this study, with Guangxi and Guangdong having highest rabies prevalence rate (respectively, 372 and 321 reported human deaths in 2008), and Yunan and Hainan having rapid rabies increase (respectively, 109 and 106 reported human deaths in 2008) (The Ministry of Health of China 2008). Although three species of bats were identified as seropositive, frugivorous R. leschenaulti appeared most important because seropositive individuals of this species were identified from four sites in three provinces. It should be noted that the assay of 57 serum samples, comprising 4 insectivorous species collected from a cave in township Tanshui of Yangchun City of Guangdong Province, produced 3 seropositives (2 from R. blythi and 1 from R. ferrumequinum), indicating that these two species from this hotspot should be further studied. Because of small numbers involved and the potential for nonrepresentative sampling, it is difficult to draw a firm conclusion. Moreover, positive antibodies do not exclude antigenic cross reactivity by other extant lyssaviruses (Reynes et al. 2004, Hanlon et al. 2005). Therefore, a larger sample size from wider areas as well as cross-neutralization tests against other lyssaviruses will help better define the seroprevalence of lyssaviruses in bats in China.
In North America, some species of insectivorous bats appear to be natural RABV hosts (Davis et al. 2006), and epidemiological studies have shown that most human rabies cases reported in the United States in the last 40 years were caused by RABV variants from insectivorous bats (Messenger et al. 2002). An investigation of lyssavirus prevalence in bats from Cambodia showed that the overall seropositive rate of neutralization antibody against four members (RABV, European bat lyssavirus type-1, ABLV, and Lagos bat virus) was 16% with a greater prevalence in frugivorous bats. Cynopterus sphinx and Pteropus lylei were the two main species in Cambodia with the highest rates of lyssavirus seroprevalence (Reynes et al. 2004). Similar studies undertaken in Thailand revealed 4% lyssavirus seroconversion in bats when tested using a modified rapid rabies fluorescence inhibition test against ABLV, and Pteropus lylei was the most seroprevalent species, although neutralizing antibodies against RABV were not detected in any species (Lumlertdacha et al. 2005). In the Philippines, the neutralization antibody–positive rate of bats was 9.5% when tested against ABLV, mainly in Miniopterus schreibersi (Arguin et al. 2002).
Although China is the largest country in Southeast Asia, lyssavirus infection in bats has not been investigated there until now. This study has provided the first preliminary data addressing the prevalence of this disease. Further investigation is needed to determine which lyssaviruses are circulating, and potential threat posed to public health. In Southern China, R. leschenaulti is a very common bat species and its fruit-eating trait brings this species into proximity with human communities. Additionally, in some regions of Southern China people have a dietary predilection for the meat of wild animals, including fruit bats. Therefore, bat viruses, including lyssaviruses, in China might pose a considerable threat to human health. In this regard, surveillance on these animals would likely play an important role in understanding the natural ecosystem and epidemiological patterns of bat viruses, and may contribute to the formulation of rational strategies for the prevention of emergence of bat-associated viral diseases.
Footnotes
Acknowledgment
This work was funded by National Natural Science Foundation of China (Project No. 30600445), National 863 Programme (2006AA02Z456), China–UK Science Networks (No. 0531105), and the Department of Science and Technology of Guangdong Province (Key Project No. 2004A20403001). The authors specially thank Dr. Libiao Zhang at the Institute of Zoology, Chinese Academy of Sciences, Beijing, and Dr. Keping Sun at Northeast Normal University, Changchun, whose strong background in bat taxonomy provided us critical assistance in identifying bat species.
Disclosure Statement
The authors have declared that no competing financial interests exist.