
Abstract
Zoonotic cutaneous leishmaniasis is an important health problem in Iran and a great economic burden on the health resources. In southern Iran (Fars Province), Meriones libycus was reported as the main reservoir of zoonotic cutaneous leishmaniasis in Arsanjan and Marvdasht cities, and Tatera indica and Gerbillus spp. were the reservoirs reported in Larestan and Kharameh districts. Because of an increase in human cutaneous leishmaniasis in Fars Province, this study was performed to identify the rodent hosts in this region. From April 2004 to April 2006, live traps were used to catch rodents in different parts of Fars Province. Fifty-seven Rattus norvegicus were caught and checked for Leishmania infection using a combination of microscopy, culture, nested polymerase chain reaction (PCR), and enzyme electrophoresis. One female R. norvegicus was found to be smear positive for amastigotes in Giemsa-stained skin sample of sole, and it was also culture positive for Leishmania. Results of PCR and isoenzyme electrophoresis indicated that this infected rodent was harboring Leishmania major. PCR was also positive for L. major in biopsy of soles, ear, liver, and spleen of 29 other R. norvegicus hosts that were negative in smears and cultures. There were no lesions seen in any parts of infected rodents' bodies. As L. major has not been previously reported in R. norvegicus in Iran or elsewhere, the rodent can be considered as a possible reservoir in transmission of the disease in Fars Province, and it should be brought into consideration when planning for preventive measures.
Introduction
Materials and Methods
Study area
Fars Province is one of the main and environmentally fragile areas in southern Iran, with an occupied area of 122,780 km and a population of 323,626. Shiraz is the major city of the province, with an altitude of 1500 m above sea level. It has cold, rainy winter and moderate to hot dry summer. The average temperature of Shiraz is 16.8°C, ranging between 4.7°C and 29.2°C. The geographical and climatic variation of the province causes variations in plants, and consequently, variation of the wildlife. Other cities included in this study are located around Shiraz, with moderate differences in geographical characteristics (Fars Budget and Planning Organization 2000).
Trapping
Rodents were caught alive in 35 × 12 × 12-cm wire traps baited with bread and roasted peanuts (Lacher et al. 1989). Between April 2004 and April 2005, 100 traps were set inside and outside houses and in agricultural plantations close to them, in both rural and urban settings, in or close to following cities: Shiraz, Kazerun, Kavar, Arsanjan, Marvdasht, Zarghan, and Estahban. At each collection site, traps were left for 96 consecutive nights, being checked and rebaited early each morning. Trapped animals were identified by the staff of Shiraz University, Department of Biology, using the relevant taxonomic criteria: forms of teeth, comb of tooth, shape of cranlum, size, and prominence of food pad (Eisenberg and Redford 1999). Fars Environmental Protection Organization gave the permission for trapping, and all the rodents caught were handled and killed according to methods approved by the Iran Veterinary Organization.
Smears and cultures
Each animal was killed by chloroform and carefully evaluated for any skin lesions. Impression smears of tissue samples taken from ears, tail, soles, and patent lesion (Edrissian et al. 1982) were stained with Giemsa and examined microscopically for presence of amastigotes. Tissue samples collected aseptically from spleen, liver, and skin were cultured at 25°C in rabbit blood agar (Evans 1989). Cultures were checked regularly for parasites and only considered negative if no promastigotes were seen within 2 months. Parasites from any positive culture were coded and cryopreserved in liquid nitrogen, pending their identification in a PCR-based assay (see below).
DNA extraction
DNA from cultured promastigotes was extracted as describes by Motazedian et al. (2002). And DNA from soles, ear, liver, and spleen was isolated as follows: small cuts of tissue were divided into very smaller pieces using lancet and then squashed by a slender rod in an appendorf tube.
One hundred milliliters of distilled water was added and tubes were frozen and thawed in liquid nitrogen three times. Tubes were then boiled for 10 min and centrifuged at 1000 rpm for 2 min. After that, the standard method of phenol chloroform was used for DNA extraction. Briefly, a heat-sealed Pasteur pipette was used to homogenize the samples with 200 μL lysis buffer (50 mM Tris-HCl [pH 7.6]; 1 mM EDTA, and 1% Tween 20) and 12 μL of a proteinase K solution (Cinna Gen) (containing 19 μg of the enzyme/mL) in a 1.5-mL microcentrifuge tube. The homogenate was then incubated at 37°C overnight before 300 μL of a phenol:chloroform:isoamyl alcohol mixture (25:24:1, by vol) was added. After being shaken vigorously, the tube holding the mix was centrifuged (10,000 g for 10 min) and then the DNA in the supernatant solution was precipitated with 400 μL of cold, pure ethanol, resuspended in 50 μL double-distilled water, and stored at −20°C until it could be tested for Leishmania kDNA. All the procedures were done in a sterile way.
Nested PCR
This PCR method was used to reduce the risk of contamination in products. Nested PCR involves two sets of primers used in two successive runs of PCR, the second set intended to amplify a secondary target within the first-run product. First-round PCR mixture contained 0.4 U Taq DNA polymerase (Sina Gene), 2.5 mM deoxynucleoside triphosphate (Sina Gene), 2.0 mM MgCl2, 2.5 μL PCR buffer (Sina Gene), 1 μL DNA, and 40 ng of each of outer pair of primers CSB2XF (C/GA/GTA/GCAGAAAC/TCCCGTTCA) and CSB1XR (ATTTTTCG/CGA/TTTT/CGCAGAACG). Cycling conditions were as follows (Corbette): 30 cycles of 94°C for 30 s, 55°C for 1 min, 72°C for 90 s. One microliter of a 1:10 dilution in water of the first-round product was used as template for the second round under the same conditions as those for the first round, except with nested primers (“internal” to the first primer) LiR (TCGCAGAACGCCCCT) and 13Z (ACTGGGGGTTGGTGTAAAATAG). Second-round PCR product was loaded onto 1.5% agarose gel (Roche). Amplification products were separated by electrophoresis and visualized by ultraviolet illumination following ethidium bromide (Roche) staining. Parasite species were identified according to the molecular weight of DNA bands (L. tropica 750 bp, Leishmania infantum 680 bp, L. major 560 bp) (Noyes et al. 1998).
Agarose gel electrophoresis
Samples of 10 μL of the PCR products mixed with 5 μL of loading buffer were run in 1.5% agarose gel. Runs were stained with ethidium bromide and visualized under ultraviolet light. The World Health Organization reference strains L. tropica (MHOM/AZ/1974/SAF-K27), L. major (MHOM/TM/1973/5ASKH), and L. infantum (MHOM/TN/1980/IPT1) were obtained from Pasteur Institute.
Enzyme extraction and electrophoresis
Enzyme extraction from the peletted organism was done by the method described by Hatam et al. (1999).
Analysis was done using five enzymatic systems (namely malate dehydrogenase E.C.1.1.1.37, phosphoglucomutase E.2.7.51, glucose-phosphate isomerase E.C.5.3.1.9, nucleoside hydrolase I E.C.3.2.2.1, and nucleoside hydrolase II E.C.3.2.2.1) in discontinuous polyacrylamide gel electrophoresis. Electrophoresis was performed as described earlier (Evans 1989).
Sequencing
Twenty of 29 samples from different area, which showed a high-quality yield in PCR product, and according to distance from Shiraz, the capital city of Fars Province, were chosen and sequenced (Faza Biotech Iran and Macro Gene companies South Korea). Results of sequences were compared using multiple sequence alignment by CLUSTALW from
Results
Of 57 Rattus norvegicus caught in agricultural plantations close to houses (Table 1), 29 were positive by PCR. One female R. norvegicus was found to be smear positive for amastigotes in Giemsa-stained skin sample of soles, and it was also culture positive for leishmania (Table 1). Results of the PCR and isoenzyme electrophoresis indicated that this infected rodent was harboring L. major (Figs. 3 and 4). PCR was also positive for L. major in soles (23), ears (18), liver (10), and spleen (5) of R. norvegicus hosts, which were negative in smears and cultures (Fig. 4). The infected rodents were caught from Shiraz, Kazerun, Kavar, Arsanjan, Marvdasht, Zarghan, and Estahban cities, Fars Province. There were no lesions on any parts of the bodies of R. norvegicus. Amastigotes were observed in smears of sole skin.

Rattus norvegicus captured in Marvdasht town, Fars Province, southern Iran.

The denture of Rattus norvegicus captured in Marvdasht town, Fars Province.
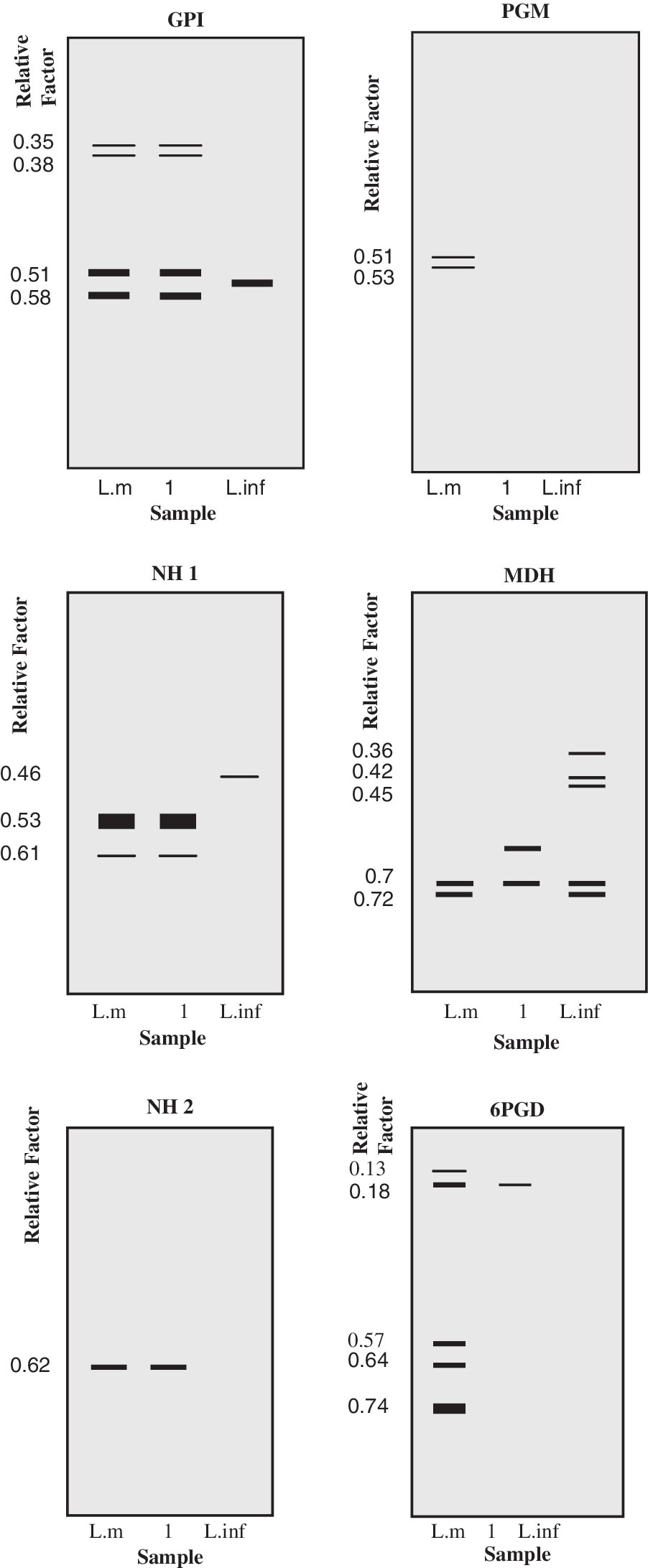
Electrophoretic profiles obtained with soluble extracts of Leishmania promastigotes, using five enzymatic systems in Rattus norvegicus in Marvdasht. L.inf, Leishmania infantum, L.m, Leishmania major.
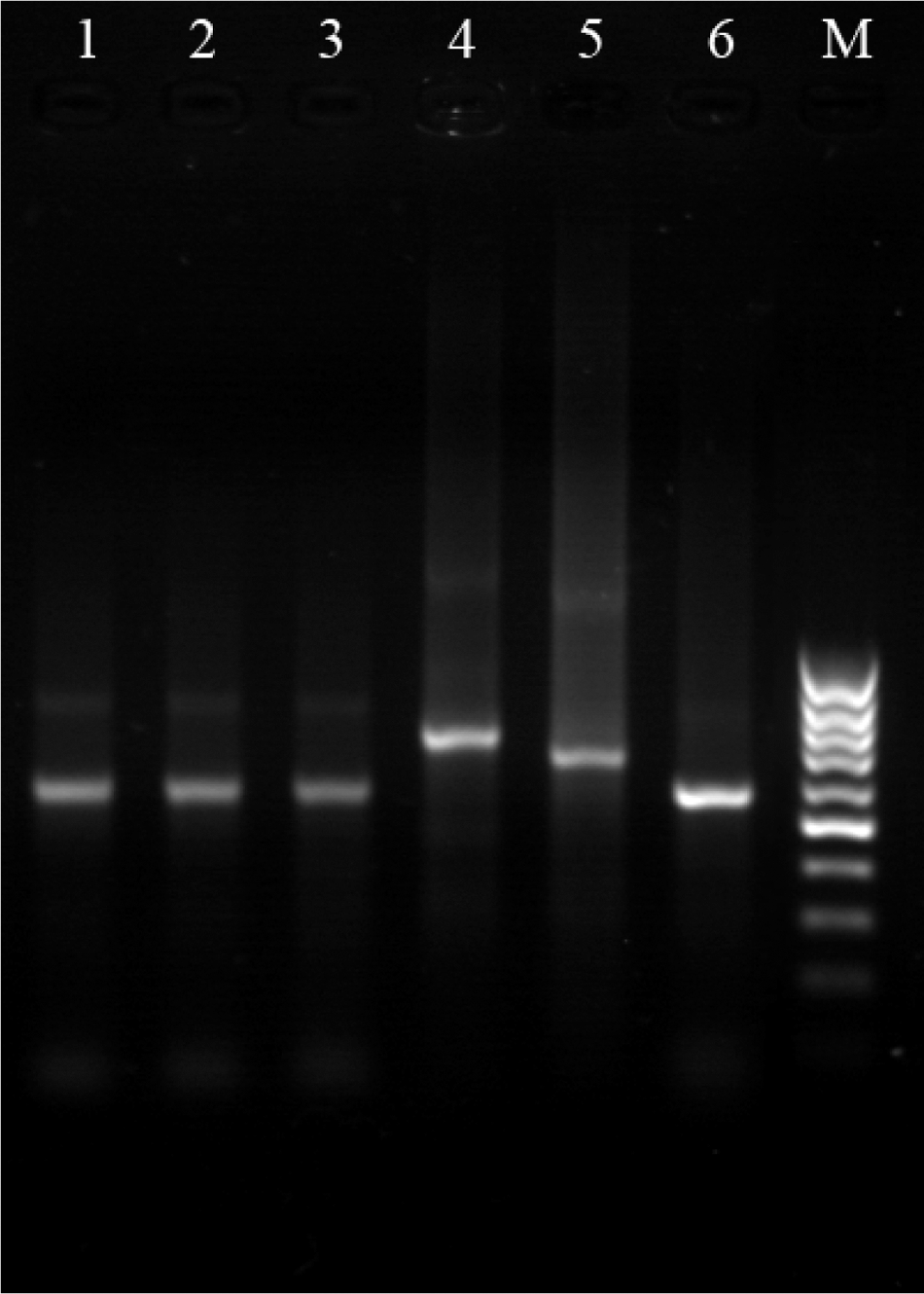
Agarose (1.5%) gel electrophoresis of polymerase chain reaction products: M: marker; 1: isolated parasite in culture from R. norvegicus; 2: isolated parasite in liver from R. norvegicus; 3: isolated parasite in spleen from R. norvegicus; 4: Leishmania tropica (MHOM/IR/89ARD2) reference strain; 5: L. infantum (MHOM/FR/59/LEM 188) reference strain; 6: L. major (MHOM/IR/54/LV39) reference strain.
PCR, polymerase chain reaction.
Sequencing result
According to dendrogram obtained by this method, infected mice were classified into two main groups. One group contains infected mice from different areas far from each other. Mice number 5 (5m) is from Kazeroun 100 km far away from number 15 (15m) Marvdasht, indicating genetic similarity in different area. High similarity was observed between mice from one place and different places, for example, number 11m from Shiraz suburb, 9m also from shiraz suburb, 13m from Shiraz suburb, and 15m from Marvdasht.
The other group contains three main branches with 14 infected mice. One sequence of L. major (21 L. major) was obtained from GenBank (Smith et al. 1989) and compared with these samples. It showed similarity with 2m and 18m from Shiraz. High similarity and genomic distance were also observed between mice from one place and different places (Fig. 5).

Twenty polymerase chain reaction products of Leishmania major minicircles isolated from infected Rattus norvegicus. Number 1m to 20m from sh: shiraz; sh su: shiraz suburb; ka: kazerun; ma: marvdasht. Number 21: L. major minicircle from gene bank.
Discussion
Human ZCL caused by L. major is a widely distributed zoonotic disease in several countries. In Sudan, the reservoir host for ZCL has been the Nile rat (Arvicanthis niloticus) (el-Hassan and Zijlstra 2001). Meriones sacramenti has been reported from North Sinai, Egypt, as a reservoir host (Morsy et al. 1993), and Psammomus obesus has been reported as the reservoir host of ZCL in Jordan, Algeria, and Egypt (Saliba et al. 1994, Harrat et al. 1996, Morsy et al. 1996). Tatera robusta is the main reservoir host of ZCL in Baringo district of Kenya (Githure et al. 1996). In Israel, Meriones crassus and P. obesus were the reservoir hosts (Schlein et al. 1984), and also Gerbillus dasyurus and P. obesus were positive for Leishmania in Nizzana, Israel (Waaserburg et al. 2002). Rioux et al. (1986) isolated L. major from the rodent Meriones shawi shawi in central Tunisia. A misleading trypanosomatid parasite (Leishmania amastigote) has been isolated from Gerbillus pyramidum and Gerbillus andersoni in an endemic area of L. major in North Sinai, but the parasites failed to be identified as a species (Mikhail et al. 1996). L. major infection was reported in P. obesus, M. crassus, and M. libycus in Saudi Arabia and in P. obesus in Syria (Rioux et al. 1992, el Sibae et al. 1993). Parasitic infection in R. norvegicus was reported first by Morsy et al. (1988) from Alexandria, Egypt, but despite isolation and growth of parasite on blood agar, they failed to determine and confirm the species of the parasite by isoenzyme technique. Giannini (1985) showed that experimental infection in Sprague–Dawley rats, which were injected intradermally with L. major promastigotes, indicates possible involvement of R. norvegicus in the transmission cycle of CL. Petrovic et al. (1975) reported the presence of Leishmania donovani infantum infections in Rattus rattus and R. norvegicus in some areas of former Yugoslavia.
Rhombomys opimus, M. libycus erythrousus, and Meriones persicus were shown as the rodent hosts of the disease in central Iran (Nadim and Faghih, 1968, Nadim et al. 1968, Javadian 1988, Doroudgar et al. 1995, Yaghoobi-Ershadi et al. 1996, 2001, Javadian et al. 1998, Rassi et al. 2001, Mohebali et al. 2004, Parvizi et al. 2008), and T. indica and N. indica in western Iran, Khorasan Province, and Tehran (Seyedi-Rashti and Nadim 1967, Javadian 1988, Javadian et al. 1998, Seyedi-Rashti and Salahzadeh 1990). T. indica and Meriones hurrianae were reported as natural reservoir hosts in southeast Iran (Seyedi-Rashti and Nadim, 1984, Yaghoobi-Ershadi et al. 1996). In southern Iran, Fars Province, L. major infection was reported in M. libycus in Arsanjan and Marvdasht (Rassi et al. 2001, Moemenbellah-Fard et al. 2003) and in T. indica and Gerbillus spp. in Larestan and Kharameh (Asgari et al. 2007, Mehrabani et al. 2007).
In this study, the isolated L. major from R. norvegicus was characterized by both nested PCR and isoenzyme techniques. Soles, ear, liver, and spleen of 29 R. norvegicus were also positive by PCR, indicating the possibility of this rodent Bering as a main reservoir host for L. major both around and in cities of this province. Sequences analysis of minicircles showed similarity and divergence of the parasites in one area and different areas far from each other. This indicates the genomic variation in L. major in this province. R. norvegicus is one of the rodents in the area, which lives in agricultural regions near Shiraz, the capital city, and other big cities of Fars Province. These regions are suitable for agriculture and have enough water supplies providing suitable locations for reproduction of the rodents and Phelebotomus sand flies. In our study, the reservoir host of L. major was different from those reported in other parts of Iran, which may be due to difference in plantation, weather, sunshine duration, humidity, and type of soil in the area. Our results vary with studies of Rassi et al. (2001) and Moemenbellah-Fard et al. (2003) in Arsanjan and Marvdasht, who reported L. major infection in M. libycus, which could be due to difference in the place of trapping. In our study, traps were set inside and outside homes and in agricultural plantations close to houses, whereas in the other two studies, they caught the rodents far from the houses. It seems that rodent control is a challenge to the health authorities involved in the control of ZCL in southern parts of Iran. Rapid urbanization, construction of buildings in farms near the colonies of rodents, storage of waste materials around the town, which are suitable for building nests by rodents, new agricultural projects in this area, existence of some animal shelters among old mud houses, and socioeconomic changes in the area may help increase the number of wild rodents and sand flies to provide a very efficient cycle for the transmission of the disease. Migration of refugees from Afghanistan has also provided suitable conditions for further spread of the disease (Ardehali 1996). As L. major has not been previously reported in R. norvegicus elsewhere and in Iran, the rodent can be considered as a possible reservoir in transmission of the disease in Fars Province and should be brought into consideration when planning preventive measures. Further studies are needed to shed light on the disease cycle in the area to demonstrate the role of this rodent as a definite reservoir for CL.
Footnotes
Acknowledgments
The authors thank the Office of the Vice-Chancellors for Research of Shiraz University of Medical Sciences, Shiraz University, for financial support. The authors also thank Dr. Motahareh Motazedian for help with improving the English language in the manuscript.
Disclosure Statement
No competing financial interests exist.