
Abstract
The Asian tiger mosquito, Aedes albopictus, has colonized nearly all the regions of Italy as well as other areas of Europe. During the summer of 2007 the tiger mosquito was responsible for an outbreak of Chikungunya in Italy, when this virus was brought in by a tourist of Indian origin returning from an endemic area. To increase the knowledge of tiger mosquito population dynamics, a survey was carried out from April to November 2008 in the municipalities of Arco and Riva del Garda (northern Italy) through a Biogents Sentinel™ (BG)-trap sampling. In particular, the aim of the study was to evaluate the influence of temperature and rainfall on the activity and dynamics of A. albopictus host-seeking females. The seasonal emergence of host-seeking females was strongly influenced by the minimum temperature, and a lower threshold of 13°C was identified. In addition, the threshold for the end of adult activity was found at a minimum temperature of 9°C. Host-seeking female abundance was positively affected by the accumulated temperatures over the period 3 and 4 weeks before the sampling week, possibly as a consequence of the positive effect of accumulated temperatures on larval density. Instead, accumulated precipitation over 1–4 weeks before sampling was negatively correlated with host-seeking female abundance. Finally, the activity of host-seeking females, estimated by the weekly increment in female abundance, was positively affected by the total abundance of females and by mean weekly temperatures. Our study provides useful information for predicting the dynamics of host-seeking Ae. albopictus females in northern Italy and for designing control strategies for preventing arbovirus outbreaks in areas colonized by Ae. albopictus.
Introduction
Ae. albopictus has considerable ecological, behavioral, physiological, and genetic adaptability (Hawley 1988). Over relatively few generations many populations of this species have evolved to resist cold winters in temperate areas; for example, diapausing eggs have developed cold-hardiness. In Italy, the tiger mosquito has adapted to living in a wide range of human containers, entering private houses, and feeding during the night (Romi et al. 2008). Since the tiger mosquito is an exophilic species that generally inhabits and rests in outdoor habitats, several studies have focused on the influence of the temperature and rainfall on Ae. albopictus population dynamics in the laboratory (Alto and Juliano 2001).
Here, we present the results of an intensive sampling of host-seeking Ae. albopictus females carried out in northern Italy during 2008 aimed at exploring the effects of temperature and rainfall on the activity and dynamics of this species. This work, therefore, provides an in-depth field investigation of the influence of temperature and rainfall on the abundance and activity of A. albopictus host-seeking females in Europe.
Materials and Methods
Study area
Mosquitoes were sampled in the neighboring municipalities of Arco and Riva del Garda (Fig. 1) in the Province of Trento, northern Italy (45°53′ N, 10°51′ E), from April to November 2008. This area, also referred to as Alto Garda, has a sub-Mediterranean climate, mitigated by Lake Garda, which is milder than that of the neighboring Alpine valleys and Padana Plain, especially during winter. The area has an average altitude of 70 m and natural vegetation dominated by Quercus ilex that is surrounded by a pre-Alpine habitat. The resident human population is 32,481 inhabitants.
Location of the study area in the Province of Trento, Italy.
Mosquito sampling
Adult mosquitoes were collected using 20 BG traps (BioGents, Regensburg, Germany), with the attractant BG lure, a synthetic lure developed to mimic human odors, consisting of lactic acid, ammonia, and caproic acid on a long-lasting lure (Krockel et al. 2006). Traps were located in 20 sampling sites, chosen within random samples of quadrats and georeferenced with a GPS device (GeoExplorer® 2005 series; Trimble, Sunnyvale, CA). Chosen sampling sites within the quadrats included private properties, school gardens, nursery schools, private houses, and garden centers. Each trap was connected to an electricity supply (12 V) during the entire sampling period, and the captures are conducted 24 h per day.
For analysis, sampling data were added and pooled by week, from the 14th week (1st week of April) to the 47th week (3rd week of November).
Sampled mosquitoes were transported live to the laboratory in an insulated thermal bag filled with ice and frozen at −20°C. The species and sex of each individual were identified in the laboratory with a stereomicroscope using the appropriate taxonomic keys (Schaffner et al. 2001). Adult males were excluded from the analysis. For host-seeking adult females, female abundance and female increment were calculated, where female abundance is defined as the total number of adult females captured per week in each trap, and female increment is calculated on the difference between the logarithm (base 10) of the number of females captured in a specific week and trap and the number of females sampled during the previous week in the same trap.
Climatic data
Daily mean, minimum, and maximum air temperatures and daily total precipitation for the study period were obtained from the Arco meteorological station (
Statistical analysis
To quantify the threshold temperatures for the emergence and the end of activity of host-seeking females, the intercept of linear regressions between the weekly female abundance (from weeks 14 to 26 for emergence and 38 to 47 for the end of activity), and average minimum temperatures of the week before sampling was computed. Generalized Linear Mixed Model (GLMM-IRREML) with negative binomial distribution and logarithmic link was used to investigate whether Ae. albopictus adult female abundance was influenced by temperature and rainfall. The response variable was the total weekly Ae. albopictus adult female abundance, and the selected explanatory variables were the average weekly temperatures (minimum, mean, and maximum), the total weekly precipitation, and all the accumulated temperature and rainfall variables (see Climatic data section for details). A linear mixed model (REML) was used to assess the relationship between the weekly increment in Ae. albopictus adult females and the temperature and rainfall variables. The weekly increment was the response variable, whereas Ae. albopictus adult female abundance and both weekly and accumulated temperature and rainfall variables were included as independent factors. To account for spatial and temporal variation among different traps, the trap identification code and the trapping week were entered as random factors. All statistical analyses were performed using Genstat Eighth Edition (Lawes Agricultural Trust 2005), and results were considered significant if p < 0.05. Starting with the full model, including all explanatory variables, a minimal adequate model was selected using stepwise backward deletion (Crawley 2002); several models were tested, but only the best models are presented and discussed.
Results
A total of 5140 female Ae. albopictus mosquitoes were sampled from April to November 2008. No Ae. albopictus adults were collected during the first 6 weeks of sampling.
Adult female abundance increased from the 20th to the 37th week, with a peak at week 31 and a second higher peak at week 37 (Fig. 4). Female abundance dropped sharply from week 38 and continued to decrease until the final sample had been collected (week 46).
A positive linear relationship (F 1, 140 = 23.51; p < 0.001) between weekly female abundance from week 14 to 26 (when abundance was increasing) and the average minimum temperature recorded in the previous week of sampling was found (Fig. 2). The intercept of the regression gives an estimate of the lower temperature threshold for seasonal emergence of adult females, and a value of 13.07°C with a 95% confidence interval ranging from 12.6°C to 13.5°C was found. From the 38th to the 47th week (when abundance was decreasing) a negative linear relationship (F 1, 151 = 22.27; p < 0.001) between adult female abundance and the average minimum temperature of the previous week of sampling was found. In this case, we found a value of the intercept that indicates an estimated threshold for the end of adult female activity of 9.01°C with a 95% confidence interval ranging from 8.52°C to 9.5°C (Fig. 3).
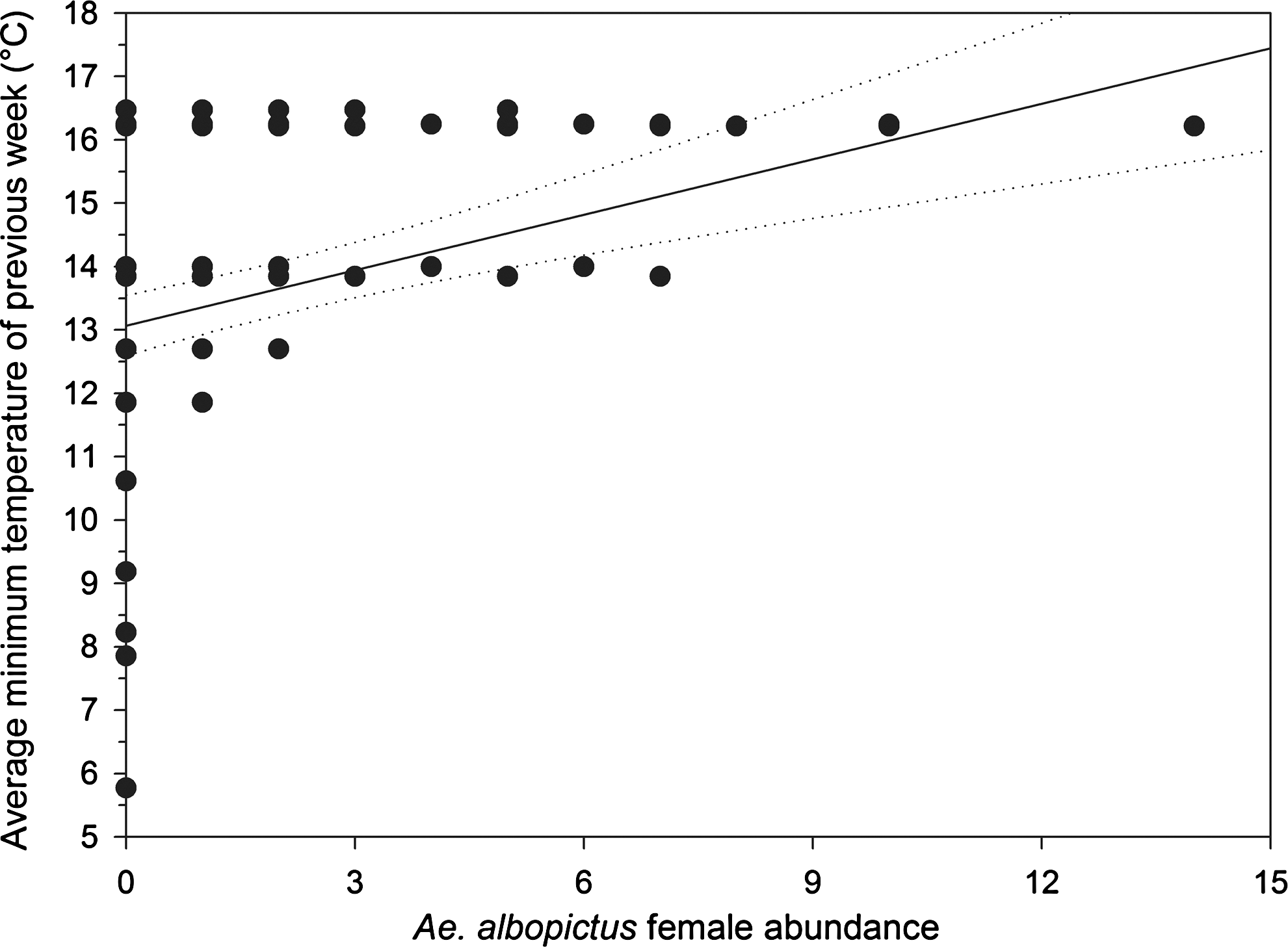
Relationship between the average minimum temperatures of the week before sampling and the abundance of Aedes albopictus females (see text for details). The threshold (intercept) for seasonal emergence of Ae. albopictus adult females is 13°C.

Relationship between the average minimum temperatures of the week before sampling and the abundance of A. albopictus females. The threshold (intercept) for the end of activity of Ae. albopictus adult females is 9°C.
The minimal adequate model for explaining the variation in female abundance of Ae. albopictus included the accumulated temperatures of 3–4 weeks prior (i.e., the period covering the third and fourth weeks before the sampling week), the accumulated rainfall of 1–4 weeks prior, the accumulated temperatures of 1–4 weeks prior, the average weekly maximum temperature, and the total weekly precipitation (Table 1). Specifically, the accumulated temperatures of 3–4 weeks prior had a positive effect on female abundance (Wald = 71.45; p < 0.001), and the accumulated rainfall of 1–4 weeks prior had a negative effect (Wald = 4.51; p = 0.034).
Figure 4 shows the temporal pattern of Ae. albopictus female abundance and the two significant explanatory variables: the accumulated temperatures of 3–4 weeks prior and the accumulated rainfall of 1–4 weeks prior.
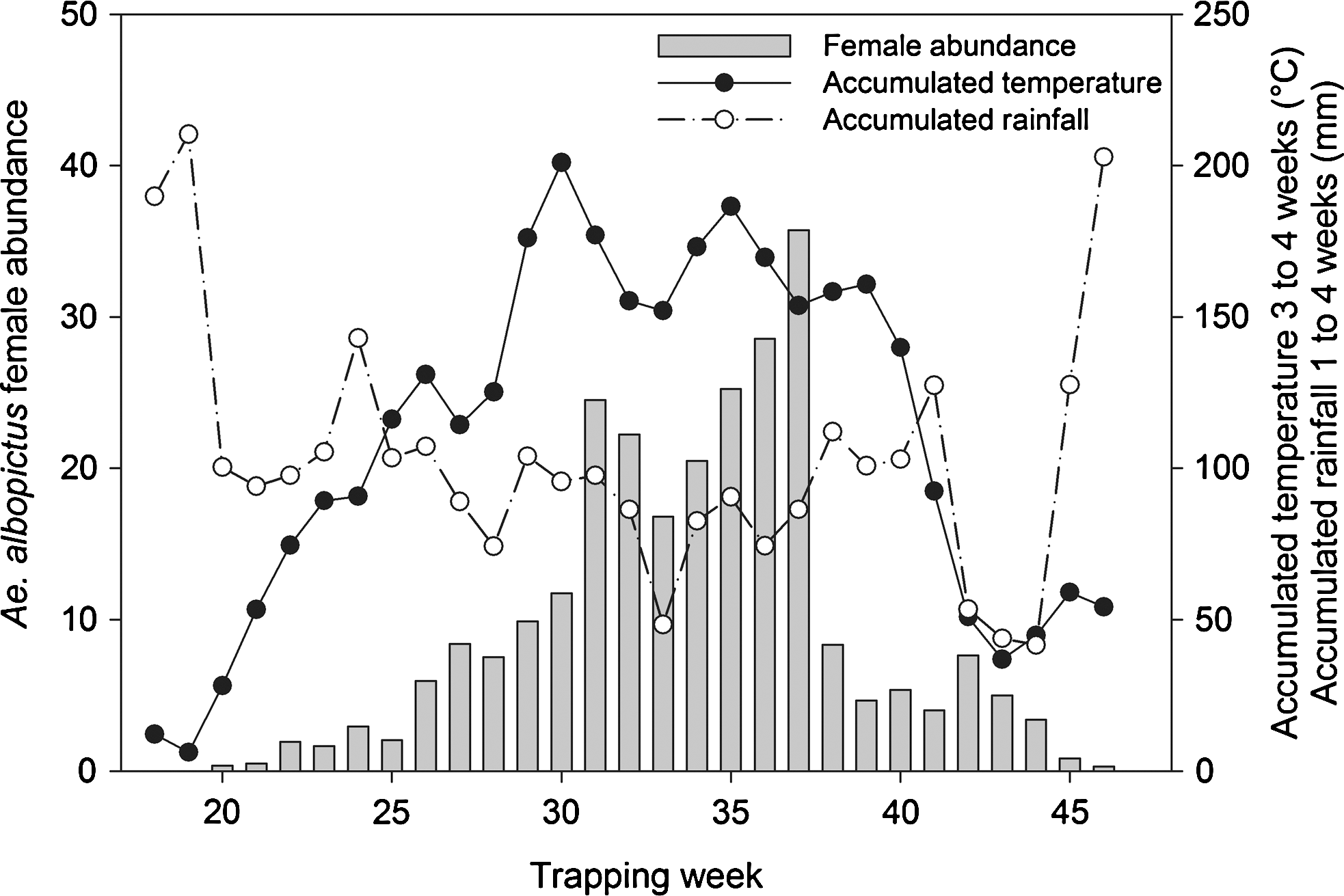
Seasonal pattern of the weekly female Ae. albopictus abundance, averaged over all trapping sites, compared to the accumulated temperatures 3–4 weeks before the sampling week and the accumulated rainfall of 1–4 weeks before the sampling week.
The minimal adequate model for explaining the increment of Ae. albopictus included female abundance, the average weekly mean and maximum temperatures, the accumulated temperatures of 3–4 weeks prior, and the accumulated rainfall of 1–4 weeks prior. Only female abundance and the mean weekly temperature were significant and had a positive effect on the weekly increment of Ae. albopictus host-seeking adult females (Table 1). Figure 5 shows the temporal pattern of Ae. albopictus female increment and the two significant explanatory variables: female abundance and mean weekly temperature.
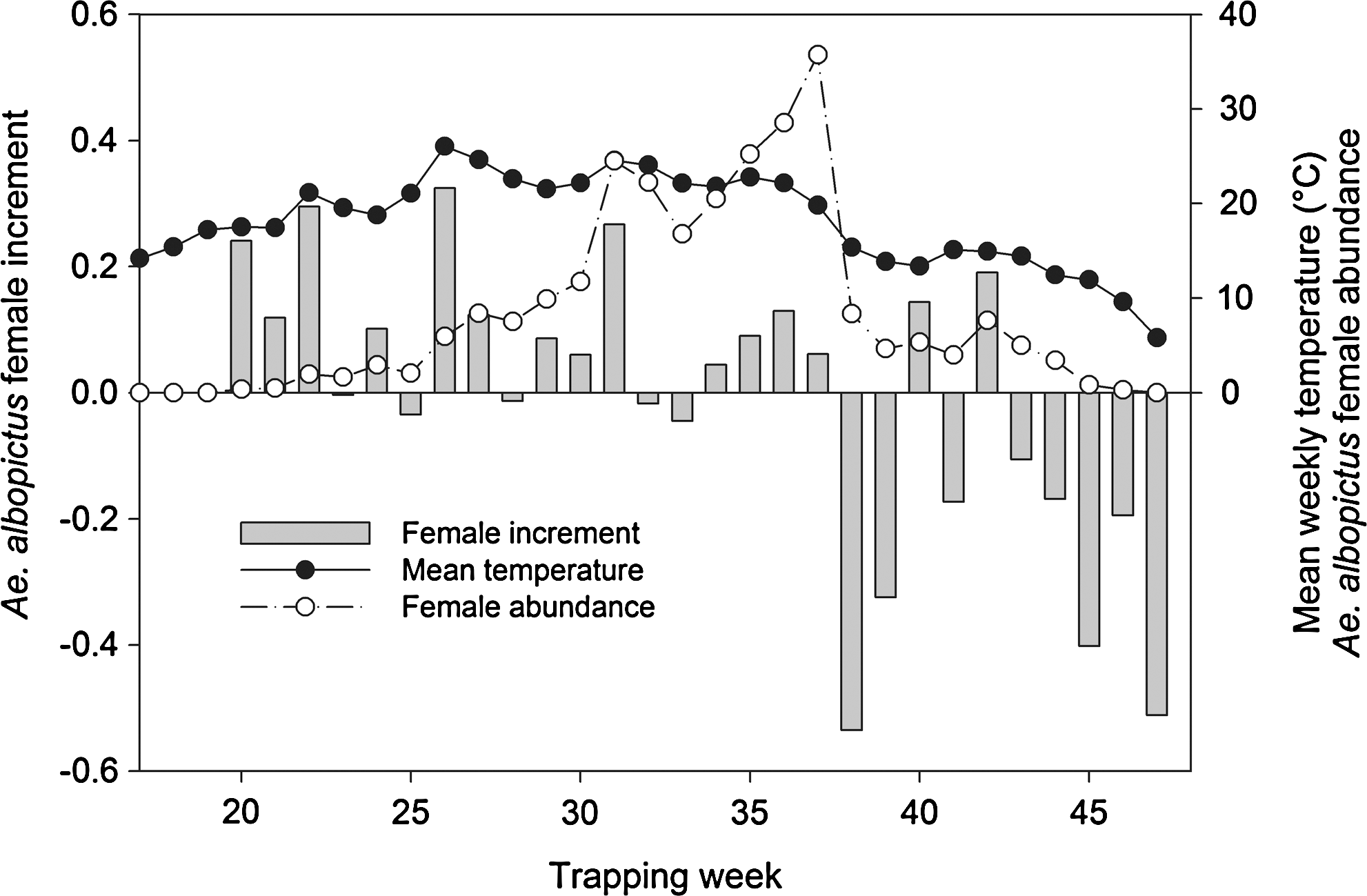
Seasonal pattern of the weekly female Ae. albopictus increment, averaged over all trapping sites, compared to the mean weekly temperature of the same sampling week and female Ae. albopictus abundance recorded in the same sampling week. Female increment is calculated on the difference between the logarithm (base 10) of the number of females captured in a specific week and trap and the number of females sampled during the previous week in the same trap.
Discussion
This work describes a detailed field investigation aimed at exploring the influence of temperature and rainfall on the abundance and activity of Ae. albopictus host-seeking females. This species is an effective vector of several arboviruses in different areas of the world, and the present study increases current knowledge of the ecology of Ae. albopictus European populations and the potential spread of mosquito-borne diseases.
The dynamics of Ae. albopictus were similar to those noted for other regions of Europe (Romi et al. 2008), but the length of the reproductive season was considerably shorter than of more southerly latitudes, such as Rome, presumably due to differences in temperature and photoperiod (Di Luca et al. 2001).
The temperature threshold necessary to initiate activity in adult female mosquitoes was found to be 13°C. This is the first time this threshold has been identified for European female Ae. albopictus populations in the field, and it is consistent with the threshold values of 11°C found for the hatching of eggs of this species in Asia, North America, and Rome (Hawley 1988, Kobayashi et al. 2002, Toma et al. 2003) and with recent studies that have modeled the distribution of Ae. albopictus in Europe and have reported that 81% of the municipalities where this vector was found have a minimal annual mean temperature above 11°C (Schaffner 2009). The temperature threshold for the end of adult activity is a mean minimum temperature of 9°C. However, mosquitoes can extend their reproductive season when the outdoor temperature decreases by seeking out a favorable microclimate that allows the development of larvae; for example, in urban environments adult mosquitoes find resting and breeding locations inside or near human habitations (Toma et al. 2003, Romi et al. 2006).
Our study clearly shows that the abundance of A. albopictus host-seeking females in northern Italy is positively influenced by the accumulated temperatures of the third and fourth weeks prior to sampling and negatively influenced by the accumulated rainfall of the first to the fourth weeks prior, the effect of temperature being stronger. These results corroborate previously published laboratory experiments, and temperature has also been found to have a stronger influence on Ae. albopictus abundance than precipitation (Alto and Juliano 2001). Several field studies in Japan, the United States, and Italy have shown a positive correlation between temperature and seasonal pattern (Toma and Miyagi 1990, Toma et al. 2003, Richards et al. 2006, Higa et al. 2007), but none of these previous studies have noted any relationship between accumulated temperatures and the abundance of host-seeking females or other stages of Ae. albopictus. As temperature strongly affects the development of larval mosquitoes, the effect of accumulated temperatures on the abundance of host-seeking females probably reflect a time delay related to the reproductive biological life cycle of Ae. albopictus in our study area.
On the other hand, a correlation between Ae. albopictus abundance and rainfall has not always been found (Hawley 1988). In fact, while a positive effect of precipitation on Ae. albopictus dynamics has been documented in several field studies (Ho et al. 1971, Toma and Miyagi 1990, Estrada-Franco and Craig 1995, Lourenço-de-Oliveira et al. 2004, Richards et al. 2006), in other cases no relationship has been found (Sulaiman and Jeffery 1986, Toma et al. 2003). The lack of a clear relationship between precipitation and Ae. albopictus abundance could be explained by the ecology of this species. This mosquito often breeds in small artificial containers located around human dwellings, so that its seasonal pattern might be more influenced by variations in human water supply than changes in precipitation. Nonetheless, heavy rains could have a negative effect on larval abundance since intense rainfall may flush the larval habitat, thereby affecting the adult mosquito population (Hassan et al. 1996, Higa et al. 2007, Koenraadt and Harrington 2008). The negative effect of rainfall should be limited to specific climatic zones with frequent periods of high precipitation, whereas it should not be found in areas of lower or more constant, lighter precipitation.
This study also indicates that Ae. albopictus host-seeker activity, measured by the weekly increase in adult females, is related to total female abundance and to mean weekly temperatures. The positive effect of temperature on female activity confirmed the results obtained in previous studies (Estrada-Franco and Craig 1995, Alto and Juliano 2001). On the other hand, large female populations facilitate the activity of host-seeking females, as measured by their weekly increase. To summarize, this study has shown that the dynamics of Ae. albopictus is strongly influenced by temperature, rainfall, and female abundance. With respect to our study area, we propose that to predict Ae. albopictus host-seeking female abundance and dynamics, the accumulated temperatures of the third and fourth weeks prior to sampling and the accumulated rainfall of the first to fourth weeks prior together with the mean weekly temperature should be monitored. These data should be useful for preventing arbovirus outbreaks in northern Italy, and for designing more effective control campaigns against this invasive mosquito, one of the worst pest vector species of this century.
Further experimental and modeling work is now needed to predict how this species will change its spatial distribution and seasonal activity in relation to changes in climate and land use in Europe where this alien species could exert a significant impact on human health.
Footnotes
Acknowledgments
The authors wish to thank Heidi Hauffe for a critical reading of the article. This work was supported by The Autonomous Province of Trento, postdoctoral project Risktiger: Risk assessment of new arbovirus diseases transmitted by Aedes albopictus (Diptera: Culicidae) in the Autonomous Province of Trento.
Disclosure Statement
No competing financial interests exist.